
Abstract
Regulatory T cells (Tregs) play an indispensable role in autoimmune disorders. FOXP3 controls the development and function of CD4+CD25+ Tregs. Common variants in the FOXP3 gene are believed to alter messenger RNA (mRNA) expression. The aim of the present investigation was to determine whether FOXP3 polymorphisms predispose an individual to the development of Graves’ disease (GD) in a Chinese population. A total of 534 GD patients who reported to and/or were admitted to the Department of Endocrinology, People’s Hospital of Zhejiang Province, Yuyao, and 630 healthy controls from similar geographical areas were enrolled in this study. FOXP3 polymorphisms (C-2383T, C-3279A, A-3499G, and T+459C) were genotyped by polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP). The relative expression of FOXP3 was assessed by reverse transcription polymerase chain reaction (RT-PCR) in 110 patients and 110 controls. Statistical analysis was performed using GraphPad Prism software. Heterozygote and minor allele (C-2383T, C-3279A, and T+459C) frequencies were significantly higher in GD patients relative to healthy controls. The relative expression of FOXP3 was significantly lower in GD patients than in healthy controls (P < 0.0001). Furthermore, FOXP3 expression was significantly associated with FOXP3 polymorphisms (C-2383T, C-3279A, and T+459C): the wild-type allele was associated with higher FOXP3 expression and mutant alleles were associated with lower FOXP3 expression. In conclusion, this study revealed that FOXP3 polymorphisms are correlated with FOXP3 expression and are significantly associated with susceptibility to GD in a Chinese cohort.
Introduction
Graves’ disease (GD) is an organ-specific autoimmune disorder characterized by the production of autoantibodies against the thyroid gland, hyperthyroidism, diffuse goiter, and abnormal thyroid hormone production. GD affects approximately 5% of the general population and is more common in females. The exact etiology of GD is unknown but is believed to be multifactorial (including both genetic and environmental factors). Various epidemiological studies have demonstrated the roles of susceptible genes in the development of GD. Possible associations between GD and environmental factors, such as smoking, stress, use of certain drugs, infectious agents, and iodine intake, are also well characterized. 1
Regulatory T cells (Tregs) play an important role in the pathogenesis of various autoimmune diseases, such as systemic lupus erythematosus, rheumatoid arthritis, multiple sclerosis, and type 1 diabetes. 2 Additionally, several reports in the literature have demonstrated an association between Tregs and GD. For example, the depletion of Tregs in an animal model increased susceptibility to GD. 3 In addition, GD patients display diminished levels of Tregs compared with healthy subjects, 4 suggesting an important role for Tregs in the pathogenesis of GD.
Tregs express forkhead box P3 (FOXP3), which is crucial for the maintenance of self-tolerance. The FOXP3 gene is located on the X chromosome and controls both the development and functions of Tregs. 5 Functional mutations in the FOXP3 gene have been associated with susceptibility to various autoimmune diseases; variants in the promoter region (C-2383T, C-3279A, and A-3499G) and intron 9 (IVS9 +459) have been associated with altered expression of FOXP3. Investigations on the roles of common FOXP3 polymorphisms with susceptibility/resistance to GD development are limited. Independent studies in Chinese, 6 Polish, 7 and Japanese 8 populations have demonstrated an association between FOXP3 mutations and susceptibility to GD. However, another study in the United Kingdom failed to demonstrate such an association. 9
To identify an association between FOXP3 variants and susceptibility to GD, as well as a relationship between altered FOXP3 expression and various genotypes, we conducted a hospital-based case–control study in a Chinese population. In this study, four common FOXP3 gene polymorphisms (promoter region: C-2383T, C-3279A, and A-3499G; and intron region: T+459C) were genotyped in GD patients and controls. The expression of FOXP3 was also quantified and correlated with the respective genotype.
Materials and methods
Patients and controls
In this case–control study, 534 GD patients who reported to the Department of Endocrinology, People’s Hospital of Zhejiang province, Yuyao, and fulfilled American Association of Clinical Endocrinologists (AACE) guidelines were enrolled. Patients with a history of any other autoimmune disorder were excluded. Serum parameters, including free T3, free T4, thyroid-stimulating hormone (TSH), and thyroid autoantibodies (TRAb, TGAb, and TPOAb) were quantified using standard procedures on samples collected before treatment. In total, 630 healthy controls from similar geographical areas without any history of autoimmune disorders were included. Intravenous blood samples (~5 mL) were collected from each participant. This study was approved by the Institutional Ethical Committee of People’s Hospital of Zhejiang Province, Yuyao, and written consent was obtained from each participant.
Genotyping of FOXP3 polymorphisms
Genomic DNA was isolated from whole blood using a DNA isolation kit (SIGMA). Genotyping of the FOXP3 polymorphisms (C-2383T, C-3279A, A-3499G, and IVS9 +459) was performed by polymerase chain reaction (PCR) followed by restriction fragment length polymorphism (RFLP) as previously described. 6
RNA extraction and reverse transcription
RNA was extracted from 300 µL of blood using TRIzol LS reagent (Invitrogen, Carlsbad, CA, USA). The concentration of isolated RNA was measured on a NanoPhotometer. To exclude possible contamination of genomic DNA, approximately 1 µg of total RNA was treated with 2U DNase (Sigma-Aldrich, St. Louis, MO, USA) for 30 min at 37°C. Next, RNA was reverse-transcribed using a First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. Complementary DNA (cDNA) was stored at −70°C until further use.
Reverse transcription PCR
FOXP3 messenger RNA (mRNA) expression was quantified in healthy controls and subjects with GD by reverse transcription PCR. In brief, each 20 µL PCR contained 2 µL of cDNA, 10 µL of 2× MESA GREEN qPCR MasterMix Plus (Eurogentec), and 15 pmol of FOXP3 primers (FOXP3-F: 5′-TCA TCC GCT GGG CCA TCC TG-3′ and FOXP3-R: 5′-GTG GAA ACCTCA CTT GGT C-3′). Glyceraldehyde-3 phosphate dehydrogenase (GAPDH) served as a control (GAPDH-F: 5′-GGT ATC GTG GAA GGA CTC ATG AC-3′ and GAPDH-R: 5′-ATG CCA GTG AGC TTC CCG TTC AGC-3′). PCR was carried out in a real-time thermal cycler under the following conditions: initial denaturation at 95°C for 4 min; 35 cycles at 95°C for 30 s, 58°C for 30 s, and 72°C for 1 min (with a single fluorescence measurement), followed by a final elongation step at 72°C for 10 min. PCR product specificity was determined by dissociation curve analysis. The melting curve program was fixed between 55°C and 95°C with a heating rate of 0.1°C/s with continuous fluorescence measurement. RT-PCR amplicons were further subjected to agarose gel electrophoresis and visualized under ultraviolet light for confirmation (expected product size of FOXP3: 487 bp, GAPDH: 187 bp). The expression of FOXP3 in each sample was calculated by the 2−ΔCt method (ΔCt = Ct of FOXP3 − Ct of GAPDH).
Results
Baseline characteristics of patients and controls
In total, 534 GD patients and 630 healthy controls were enrolled in this study. Most GD patients were female (83.5%). Therefore, to maintain uniformity, 83% of the healthy control population was female. As shown in Table 1, the levels of free T4, free T3, TSH, TR antibodies, anti-TG antibodies, and anti-TPO antibodies were significantly higher in GD patients relative to controls. All patients were treated with methimazole or propylthiouracil according to standard protocols.
Baseline characteristics of Graves’ disease patients and healthy controls.
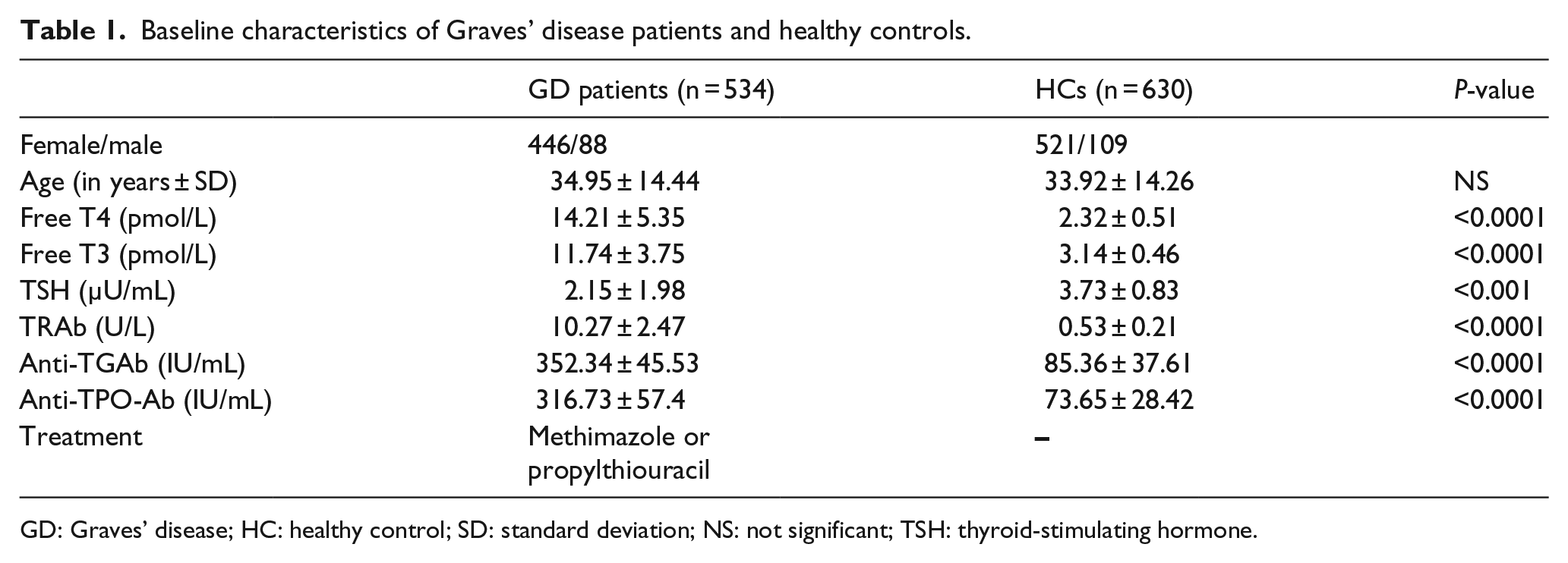
GD: Graves’ disease; HC: healthy control; SD: standard deviation; NS: not significant; TSH: thyroid-stimulating hormone.
Prevalence of FOXP3 polymorphisms in healthy Chinese population
All healthy controls were successfully genotyped for C-2383T, C-3279A, A-3499G, and IVS9 +459 by PCR-RFLP, as described in section “Materials and methods.” The frequencies of various genotypes are shown in Table 2. For the C-3279A polymorphism, CC and CA genotypes were more frequent than the homozygous mutant (CC, 74%; CA, 21%; AA, 5%). The distribution of IVS9 +459 followed a similar pattern (TT, 75%; TC, 22%; CC, 3%). However, in the other two FOXP3 polymorphisms (C-2383T and A-3499G), the heterozygous genotype frequency was higher than those of C-3279A and IVS9 +459 (C-2383T: CC, 62%; CT, 32%; TT, 6%; and A-3499G: AA, 62%; AG, 30%; GG, 8%). The genotype distributions were not in Hardy–Weinberg equilibrium for C-3279A and A-3499G (C-2383T: χ2 = 2.48, P = 0.11; C-3279A: χ2 = 25.82, P < 0.0001; A-3499G: χ2 = 14.43, P = 0.0001; and IVS9 +459: χ2 = 3.79, P = 0.05).
Genotype and allele distributions of FOXP3 polymorphisms in Graves’ disease patients and healthy controls.

HC: healthy control; GD: Graves’ disease; OR: odds ratio; CI: confidence interval.
Figures in bold are statistically significant.
Associations between FOXP3 polymorphisms and GD
To reveal a potential association between FOXP3 polymorphisms and GD, the distribution of alleles and genotype frequencies were compared between GD patients and healthy controls. As shown in Table 2, the homozygous mutant (TT) and minor allele (T) frequencies of the C-2383 polymorphism were significantly higher in GD patients than in healthy controls (TT: P = 0.001, odds ratio (OR) = 2.05, 95% confidence interval (CI) = 1.32–3.17; T: P = 0.003, χ2 = 8.35). For the other two single-nucleotide polymorphisms (SNPs) (-3279 and IVS9 +459), the prevalence of heterozygous mutants and variant alleles was significantly higher in GD patients than in controls (-3279 (CA: P < 0.0001, OR = 1.77, 95% CI = 1.35–2.31; A: P < 0.0001, χ2 = 18.17); IVS9 +459 (TC: P < 0.0001, OR = 1.70, 95% CI = 1.30–2.21; C: P < 0.0001, χ2 = 15.26)), indicating an association between these mutations and susceptibility to GD. However, the distribution of the A-3499G polymorphism was comparable among patients and controls (Table 2). A post hoc power analysis revealed that this study had 99% power to detect a possible association between FOXP3 polymorphisms and susceptibility/resistance to GD.
GD patients display decreased FOXP3 expression relative to controls
To investigate a possible role for FOXP3 in GD, we quantified the expression of the FOXP3 gene in 110 GD patients and 110 healthy controls by RT-PCR. Relative expression of the FOXP3 gene was compared in two clinical categories using Student’s t-test. Patients with GD displayed significantly lower levels of FOXP3 compared with healthy controls (P < 0.0001), as shown in Figure 1.
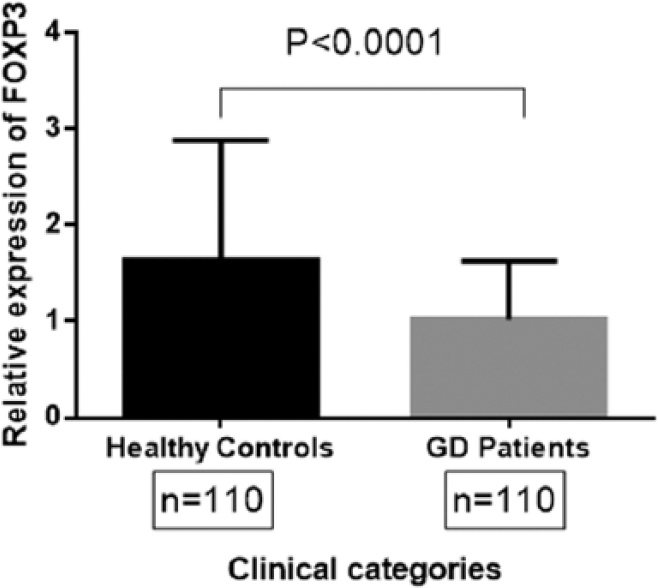
Relative FOXP3 expression in healthy controls and Graves’ disease (GD) patients. FOXP3 gene expression in healthy controls (n = 110) and GD patients (n = 110) was quantified by RT-PCR. Mean relative expression of FOXP3 in healthy controls and GD patients was compared by Student’s t-test. A P-value less than 0.05 was considered significant.
Associations between FOXP3 polymorphisms and mRNA expression
Of 1164 subjects, mRNA was isolated from 220 individuals (110 patients and controls each) to reveal potential associations between FOXP3 polymorphisms (C-2383T, C-3279A, A-3499G, and IVS9 +459T>C) and relative FOXP3 expression (Figure 2). A significant association between FOXP3 polymorphisms (C-2383T, C-3279A, and IVS9 +459T>C) and mRNA expression was observed: individuals with the wild-type allele (C-2383T: CC; C-3279A: CC; IVS9 +459T>C: TT) displayed a significantly higher FOXP3 mRNA level relative to those with the heterozygous (C-2383T: CT; C-3279A: CA; IVS9 +459T>C: TC) and homozygous (C-2383T: TT; C-3279A: AA; IVS9 +459T>C: CC) mutant allele. However, the difference between heterozygous (C-2383T: CT; C-3279A: CA; IVS9 +459T>C: TC) and homozygous (C-2383T: TT; C-3279A: AA; IVS9 +459T>C: CC) mutants did not reach significance. No significant association between the other FOXP3 polymorphism (A-3499G) and mRNA expression was observed.

Association between FOXP3 polymorphisms (C-2383T, C-3279A, A-3499G, and IVS9 +459T>C) and FOXP3 expression. FOXP3 gene expression in healthy controls and GD patients was quantified by RT-PCR and compared among the different FOXP3 polymorphisms (a: C-2383T; b: C-3279A; c: A-3499G; and d: IVS9 +459T>C). A P-value less than 0.05 was considered significant.
We also analyzed an association between FOXP3 polymorphisms and its expression in GD patients and controls individually. Interestingly, the association between FOXP3 genotype and expression remained both in GD patients and healthy controls (data not shown). These observations suggest important roles for FOXP3 variants in the determination of FOXP3 levels in subjects, irrespective of clinical phenotype.
Discussion
The role of Tregs in GD pathogenesis has been well characterized. Various studies have demonstrated lower levels of CD4+CD25+ Tregs in GD patients. In addition, a mouse model of Graves’ hyperthyroidism showed diminished levels of CD4+CD25+ Tregs, suggesting the importance of Tregs in GD. FOXP3 plays an indispensable role in the development and functions of Tregs. The suppressive function of Tregs against the development of autoimmune diseases is hampered by decreased levels of or a deficiency in FOXP3. 10 In this study, we observed decreased expression of the FOXP3 gene in GD patients relative to healthy controls, which corroborates an earlier report by Wang et al. 11 Similarly, a mouse model of GD hyperthyroidism also demonstrated diminished FOXP3 gene expression, indicating a vital role for FOXP3 in the pathogenesis of GD.
Although several SNPs in FOXP3 have been reported, the present investigation included four common SNPs with functional relevance. Two promoter polymorphisms (C-2383T and C-3279A) and IVS9 +459T>C were associated with FOXP3 mRNA expression: individuals with the wild-type allele displayed increased FOXP3 expression relative to those with the heterozygous and/or homozygous mutant allele. These observations were also true for both GD cases and healthy controls. In contrast, another FOXP3 polymorphism (A-3499G) did not correlate with ITS expression. Consistent with our observations, a recent promoter assay showed decreased relative activity of the FOXP3 promoter in the mutant allele (A) compared with the wild-type allele (C) for the C-3279A polymorphism. 6 The exact mechanism by which these variants control mRNA expression is not known. A mutation in FOXP3 may hamper the binding of crucial transcription factors that would lead to diminished FOXP3 expression. An earlier study in psoriasis patients showed that rs3761548 variants of the FOXP3 gene abrogate the binding of E47/c-Myb, 12 also supporting the functional relevance of FOXP3 promoter polymorphisms.
Studies on an association between FOXP3 gene polymorphism (C-2383T) and human diseases are limited. Variants of the C-2383T polymorphism have been associated with susceptibility to lung cancer and systemic lupus erythematosus in Iranian 13 and Guangxi Zhuang populations. 14 In this study, we observed a significant association between the homozygous mutant (TT) and minor allele (T) and susceptibility to GD. Subjects with the minor allele had an eightfold higher chance of developing GD. However, a recent study on a Han Chinese population failed to demonstrate an association between this polymorphism and the development of GD. 6 One possible explanation for the inconsistent results could be the smaller sample size of the earlier report. For a more concrete conclusion, we examined a larger cohort. This study also demonstrated the functional relevance of the C-2383T polymorphism, where mutants express a significantly lower level of FOXP3 compared with wild-type (CC). Diminished FOXP3 expression in GD cases could be due to the higher prevalence of FOXP3 C-2383T mutants.
Although an association between the FOXP3 C-3279A polymorphism and the development of various autoimmune disorders, including Behcet’s disease, rheumatoid arthritis, nondermatomal vitiligo, and psoriasis, has been documented in different populations, 15 its association with GD development is limited and remain contradictory. FOXP3 C-3279A mutants are associated with susceptibility to GD in Japanese 8 and Chinese populations; 6 however, studies on individuals from the United Kingdom 9 and Poland 7 did not reveal an association. In this study, the heterozygous (CA) and mutant allele (CA + AA) frequencies were significantly higher in GD cases compared with healthy controls, corroborating earlier observations in Chinese patients. The IVS9+459 T/C polymorphism is located in intron 9 of the FOXP3 gene and is believed to affect the splicing process, leading to the production of a less functional protein. A recent report showed an association between the intron 9 polymorphism and susceptibility to GD in a Han Chinese population, 6 consistent with the findings reported herein: the mutant allele (TC + CC) and minor allele (C) were more prevalent in GD patients than in controls (1.7- and 15.26-fold increased risk for the development of GD, respectively). In addition, the FOXP3 (IVS9+459 T/C) polymorphism was associated with the expression of FOXP3. However, the precise mechanism by which intron 9 controls FOXP3 expression should be further investigated.
In conclusion, diminished FOXP3 expression is associated with GD. Furthermore, subjects carrying the minor allele and the mutant genotype for C-2383T, C-3279A, and IVS9 +459 T/C FOXP3 polymorphisms are more susceptible to GD development. Moreover, these genotypes are associated with FOXP3 expression, irrespective of clinical phenotype. Although a larger number of subjects were enrolled in the present investigation, the results should be validated in other populations with a sufficient sample size.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.