
Abstract
Cardiomyocyte apoptosis is thought to play an important role in sepsis-induced cardiodepression. Previous studies mainly focused on the role of exogenous TNF-α in sepsis-induced cardiac damage, however, the role of endogenous TNF-α is rarely known. Therefore, we hypothesized that endogenous TNF-α also contributed to sepsis-induced cardiomyocyte apoptosis. Primary neonatal rat cardiomyocytes were time- and dose-dependently stimulated with BLP and TNF-α. In separate experiments, cells were treated with TNF-α antagonist and IL-10, respectively, to determine effects of endogenous TNF-α and exogenous IL-10 on BLP-induced cardiomyocyte apoptosis. After treatment, apoptosis was evaluated by nuclear condensation, membrane permeability change, caspase-3 activation, and pro- to anti-apoptotic protein (bax to bcl-2) expression. Treatment of cardiomyocytes with BLP and TNF-α both significantly induced caspase-3 activation in a time- and dose-dependent manner and caused apparent nuclear condensation and increased membrane permeability. TNF-α antagonist pretreatment attenuated BLP-induced caspase-3 activation, and downregulated bax/bcl-2 ratio. In addition, administration of IL-10 inhibited TNF-α production and suppressed cardiomyocyte apoptosis induced by BLP. Our data suggest that endogenous TNF-α play an important role in BLP-induced cardiomyocyte apoptosis and IL-10 protect cardiomyocytes from BLP-induced apoptosis, an effect partially through inhibition of endogenous TNF-α production.
Keywords
Sepsis is one of main causes of death in the intensive care unit. According to an American retrospective survey, the incidence of sepsis is rising at an annual rate of 8.7%. 1 Severe sepsis is defined as sepsis with organ dysfunction. 2 Almost 50% of septic patients have cardiac dysfunction, the case fatality ratio of which can reach 70%. 3 In our previous study, 4 we found that the application of low doses of bacteria lipoprotein (BLP) in septic mice, making mice immune system tolerance, could improve the cardiac function compared with non-tolerant mice. It is suggested that to alleviate cardiac inflammation could probably benefit to sepsis-induced cardiac dysfunction.
Once the body suffers from infections, microorganism ingredients are able to activate immunocytes to induce rapid release of various inflammatory factors, such as TNF-α, interleukin (IL)-1 β and IL-6. Parrillo 5 in 1985 found that septic patients’ serum could obviously reduce rat cardiomyocyte contraction force, for the first time, declared the existence of myocardial inhibitory substances (MDSs). Then following studies discovered that injection of TNF-α into mice could dose-dependently cause myocardial contraction, ejection fraction, and blood pressure reduction, and induce systemic vascular resistance, with a similar performance of sepsis. Thus, the production of inflammatory cytokines such as TNF-α was considered as an important mechanism in sepsis-induced myocardial dysfunction. However, in several clinical studies, anti-circulatory TNF-α therapy had no benefit or even had harmful effects on patients with heart dysfunction.6–9 The role of TNF-α generated from the heart itself in sepsis-induced cardiac dysfunction is rarely known and searches for effective measures involved can offer potential therapeutic targets to alleviate cardiac dysfunction.
Materials and methods
Animal and chemicals
Animal
Sprague-Dawley (SD) rats, aged 1–3 days, were purchased from Experimental Animal Center of Nanjing Medical University (Nanjing, PR China). The use of the animals reported in this study was approved by the Institutional Animal Care and Use Committee of Nanjing Medical University.
Chemicals
Bacterial lipoproteins (Pam3CSK4.3HCl) were purchased from Alexis Biochemicals (San Diego, CA, USA). Recombinant rat TNF-α and IL-10 were purchased from PeproTech (Rocky Hill, NJ, USA). Dulbecco’s modified eagle medium(DMEM), fetal calf serum(FCS), hoechst 33342 reagent, collagen II, and trypsin 1:250 were obtained from Invitrogen Life Technology (Carlsbad, CA, USA). Primary antibodies for caspase-3, bax, and bcl-2 were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-α-tubulin, anti-β-actin, and antibodies against IgG conjugated with horseradish peroxidase (HRP) were purchased from Boster (Wuhan, PR China).TNF-α antagonist (WP9QY) was from Santa Cruz Biotechnology (Santa Cruz, CA, USA). TNF-α ELISA kits were purchased from Beijing 4A Biotech (Beijing, PR China). Bicinchoninic acid (BCA) protein assay kit and SuperSignal West Pico Chemiluminescent substrate were obtained from Pierce (Rockford, IL, USA).
Neonatal rat cardiomyocytes culture
Primary cardiomyocytes were isolated from neonatal SD rats using modified methods described previously. 10 In brief, 1–3-day-old SD rats were killed and their hearts were quickly removed into the cold D-Hank’s liquid to squeeze out the residual blood. After scalpel homogenization into approximately 1–3 mm3 pieces, hearts were subjected to a series of trypsin and collagenase combinization digestions at 37°C until the tissue blocks completely disappeared. Each of the supernatant except the first time was collected and neutralized by 4 mL FCS. Then the mixed supernatant was filtered, centrifuged, and resuspended. Cells were plated into a dish for 1.5 h. Then, the unattached cells were collected and placed on different culture plates in DMEM medium supplement with 10% heated-inactivated FCS, 100 U/mL penicillin, and 100 mg/mL streptomycin in a humidified atmosphere (5% CO2 and 95% air) in order to obtain a high purity of cardiomyocytes. Subsequently, 0.01 mmol/L 5-Brdu was added to the medium for the first 48 h of culture to inhibit the growth of fibroblasts.
Cytokine ELISA
Neonatal rat cardiomyocytes (0.5 mL of cell suspension per well) were seeded at a density of 2 × 105 cells/mL in 24-well plates for 72 h. Cells were incubated in serum-free medium for 12 h before treatment. For TNF-α protein assay, ELISA estimations were done on 100 μL of cell free culture medium from the collected supernatant using a commercially available kit following the manufacture’s instruction (n = 9 in each group).
Hoechst 33342 staining and Annexin V-FITC/PI staining
Nuclear condensation, which is obvious with Hoechst 33342 staining, is an important phenomenon marker during the process of apoptosis. Apoptosis of cardiomyocytes was determined by Hoechst 33342. Cardiomyocytes were cultured on 12-well plates for 72 h and divided into several groups. After treatment, the former medium was discarded and cells were fixed with 50% methanol and 50% acetone for 15 min and then incubated with 1 μg/mL Hoechst 33342 for 15 min at room temperature in the dark. Cells were then viewed under a phase contrast fluorescence microscope. Nuclear condensation was expressed as a percentage of total nuclei (n = 3 in each group).
Annexin V-FITC/PI staining – cardiomyocytes (2 × 105cells/well) were seeded in 12-well plates. After treatment, cells were washed with PBS and treated with 1× assay buffer, annexin-fluorescein isothiocyanate, and propidium iodide (PI) as per the protocol described in the Annexin V apoptosis detection kit. After 20 min, the cells were washed with PBS and apoptotic cells were viewed using a fluorescent microscope and photographed (n = 3 in each group).
Western blot
To determine effects of BLP and TNF-α, TNF-α antagonist, and IL-10 on caspase-3 activation and bax/bcl-2 protein expression, cell extracts were prepared using lysis buffer contained 20 mM pH 7.4 HEPES, 4mM EDTA, proteinase inhibitor cocktail, 2 mM Na3VO4, and 20 mM pNPP. Protein concentration of the extracts was measured by BCA Protein Assay. Proteins were loaded and run on 15% SDS-PAGE gel. Proteins from the gel were transferred to a nitrocellulose membrane electrophoretically. To avoid unspecific binding, the membrane was incubated in 5% skim milk for 2 h at 4°C and then immunobloted with specific antibodies against caspase-3, bax, bcl-2, α-tubulin, or β-actin overnight at 4°C. The membrane was subsequently washed and exposed to horseradish peroxidase-conjugated secondary antibodies for 2 h at 4°C. Finally, proteins were detected with an ECL kit and blots were quantified with scanning densitometry (n = 3 per group).
Statistical analysis
The data were presented as mean ± standard deviation. Comparison data between groups was performed using one-way analysis of variance (ANOVA). LSD’s procedure for multiple range tests was performed. P <0.05 was considered to be significant.
Results
Both BLP and TNF-α induced neonatal rat cardiomyocyte apoptosis
Caspase-3 is a critical executioner of apoptosis, as it is either partially or totally responsible for the proteolytic cleavage of many key proteins such as the nuclear enzyme poly (ADP-ribose) polymerase (PARP). First, we observed the time point at which apoptosis developed obviously. Cardiomyocytes were exposed to 1 μg/mL BLP for 0, 12, 24, 36, and 48 h. As shown in Figure 1a, cleaved caspase-3 was significantly higher from 24 to 48 h (P <0.01 VS 0 h). Second, cells were treated with BLP at concentrations of 0, 0.25, 0.5, 1, 2, and 5 μg/mL for 24 h. As shown in Figure 1b, BLP dose-dependently increased cleaved caspase-3. The level of cleaved caspase-3 was increased by 102%, 313.6%, 357.9%, and 423.8% in cardiomyocytes treated with 0.5, 1, 2, and 5 μg/mL BLP, respectively, when compared with time-matched 0 μg/mL BLP groups (P <0.01). In addition, as presented in Figure 2, in 5 μg/mL BLP alone treated groups, the pro- to anti-apoptotic protein (bax to bcl-2) ratio was significantly increased by 350.4% compared with untreated control groups (P <0.01).


We also confirmed that TNF-α led to an increase of cleaved caspase-3 in a time- and dose-dependent manner in cardiomyocytes (Figure1c and d). Twenty-four hours after TNF-α stimulation, the level of cleaved caspase-3 increased significantly by 22.3%, 40.7%, and 61.7% in cardiomyocytes treated with 20, 40, and 60 ng/mL TNF-α, respectively, when compared with 0 ng/mL TNF-α groups (P<0.01). Furthermore, 20 ng/mL TNF-α caused an apparent increase by 204.0% in bax/bcl-2 ratio after 48 h of treatment compared with untreated control groups (P <0.01) (Figure 2).
Furthermore, we examined effects of BLP- and TNF-α-induced cell apoptosis as indicated by nuclear condensation and membrane permeability change. As shown in Figure 1e, exposure to BLP and TNF-α both resulted in evident nuclear condensation and increased membrane permeability compared with that in control groups, of which the cell nucleuses were evenly stained with nattier blue and there were less Annexin V-FITC(+)/PI(+) staining cells.
Neonatal rat cardiomyocytes secreted TNF-α after BLP stimulation
In order to know whether neonatal rat cardiomyocytes was able to secrete TNF-α after BLP stimulation, cardiomyocytes were incubated with 5 μg/mL BLP for 4 to 48 h in serum-free medium. The results presented in Figure 3a indicated that cardiomyocytes could secrete basic level of TNF-α, which were in the range of 20–30 pg/mL. After BLP stimulation, the concentration of TNF-α rose and achieved the peak (52.75 ± 3.52 pg/mL) at 12 h. Then cells were treated with BLP at concentrations of 0, 0.5, 2, 5, and 10 μg/mL for 12 h. As shown in Figure 3b, TNF-α level ascended in a dose-dependent manner, at a highest concentration of (59.35 ± 3.47) pg/mL in 10 μg/mL BLP groups.
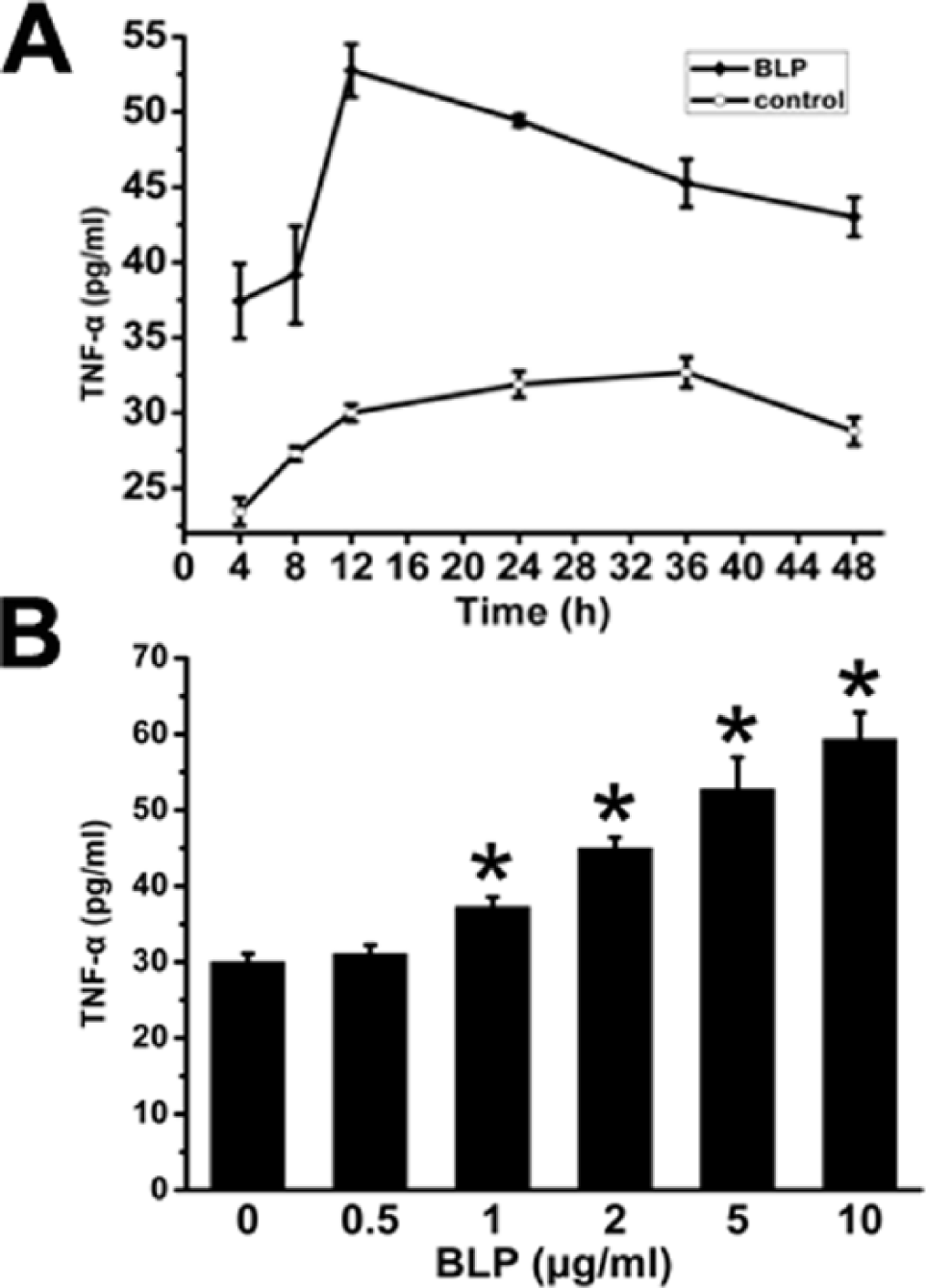
Endogenous TNF-α was involved in BLP-induced cardiomyocyte apoptosis
In order to investigate whether endogenous TNF-α generated from cardiomyocytes themselves was involved in the process that BLP induced cardiomyocyte apoptosis, WP9QY, which was able to combine with TNF-α to antagonist TNF-α effects, was used in the following experiments. 11 WP9QY (10 μM, 20 μM) was pretreated in cardiomyocytes 30 min before BLP treatment for 48 h. The results presented in Figure 2 showed that in WP9QY (10 μM, 20 μM) pretreated groups, the cleaved caspase-3 decreased by 18.9%, 44.1%, and the bax/bcl-2 ratio reduced by 5.1% and 34.8%, respectively, compared with BLP alone treated groups. In TNF-α positive control groups, WP9QY (10 μM, 20 μM) led to a remarkable decrease of cleaved caspase-3 by 44.0% and 44.5%, and bax/bcl-2 ratio by 51.3% and 56.3%, respectively, compared with TNF-α alone treated groups. Therefore, the pro-apoptotic effect of BLP was attenuated by TNF-α antagonist. It was suggested that endogenous TNF-α promoted to BLP-induced cardiomyocyte apoptosis.
IL-10 suppressed BLP-induced cardiomyocyte TNF-α production and cardiomyocyte apoptosis
First, as shown in Figure 4a, 2.5 ng/mL of IL-10 was enough to antagonist the production of TNF-α induced by 5 μg/mL BLP. In Figure 4b, cardiomyocytes were divided into five groups. In IL-10+BLP groups, cardiomyocytes were treated with 20 ng/mL IL-10 2 h prior to 5 μg/mL BLP administration for 48 h. In contrast, in BLP+IL-10 groups, 20 ng/mL IL-10 was added 12 h after cells were incubated with 5 μg/mL BLP and then co-incubated for 36 h. Surprisingly, as shown in Figure 4b, no matter whether cardiomyocytes were pretreated or post-treated with IL-10, the cleaved caspase-3 was notably reduced by 78.6% and 73.2%, and bax/bcl-2 ratio decreased by 76.5% and 66.9%, respectively, compared with BLP alone treated groups.

Discussion
Cardiac myocytes are terminally differentiated phase cells. Any cause of myocardial apoptosis, autophagy, or necrosis will reduce the number of myocardial cells, which may eventually affect the contractility of the whole heart. Of the three, apoptosis may be the main reason in sepsis.12,13 In the present study, we observed that both BLP and TNF-α were able to directly induce cardiomyocyte apoptosis. When the bacteria invade into circulation, BLP, widely existing on bacterial cytomembranes, is released to initiate inflammation. A total of 1 μg/mL of BLP caused an obvious cleavage of caspase-3 after stimulating cardiomyocytes for 24 h and then gradually increased with the extension of time. Moreover, different concentrations of BLP dose-dependently promoted expression of cleaved caspase-3. We also found that BLP led to visible cardiomyocyte nuclear condensation and significant rise of bax to bcl-2 ratio. Therefore, not only lipopolysaccharides, which just exist on gram negative bacteria, but also BLP could directly induce cardiomyocyte apoptosis. In addition, the experiment found that TNF-α could significantly activate caspase-3 and improve bax/bcl-2 ratio, thus inducing apoptosis of cardiomyocytes. BLP has been observed to have similar effects of TNF-α, showing that the pro-apoptotic mechamism of BLP has internal consistent mechanism with TNF-α. Our previous studies confirmed that BLP could induce human monocytes (THP-1) rapid release of TNF-α. 14 The question that whether BLP was able to stimulate cardiac cells to produce TNF-α and promote cardiomyocyte apoptosis was answered by the following study.
We have previously reported that 1 μg/mL of BLP was able to stimulate THP-1 cells (number of 2 × 105) for 4 h to a rapid release of TNF-α, a concentration of about 600 pg/mL. However, stimulation of cardiomyocytes (a cell count of approximately 0.5 × 105) with 5 μg/mL BLP in this experiment also resulted in visible expression of TNF-α, peaking at 12 h after the stimulus. There were several different results on the expression level of TNF-α in cardiomyocytes, but the secretion phase of TNF-α is basically identical, which reached a peak during 8–14 h. For example, Xiao-Qiang Li 15 used 100 ng/mL of LPS to stimulate neonatal rat cardiomyocytes and cells were found to release a gradual increase of TNF-α, peaking at 12 h, with a concentration of approximately 100 pg/mL. Then, we used different concentrations of BLP to stimulate cardiomyocytes, and found that TNF-α level increased gradually, but not in a proportional ratio. This is similar to the findings of Wang HD: 16 after 100 μg/mL of LPS stimulus to cardiomyocytes, TNF-α level just increased by 10 pg/mL than 10 μg/mL of LPS. Collectively, these data show that cardiomyocytes are also able to produce inflammatory cytokine TNF-α after stimulation. Therefore inferred from this, the impact of sepsis on cardiac function can be not only from the systemic inflammatory response, but also from endogenous inflammatory cytokines deriving from myocardial cells themselves.
In this part of the experiment, we used the WP9QY, a TNF-α receptor analogues, to antagonize the effects of endogenous TNF-α. The experiment observed that 10 μM WP9QY pretreatment attenuated 5 μg/mL BLP-induced caspase-3 activation. Decrease of bax/bcl-2 protein ratio further confirmed the anti-apoptotic property of WP9QY. In order to prove the effect of endogenous TNF-α was really inhibited by WP9QY, we used the TNF-α- stimulated cardiomyocytes as positive control group. We observed that 10 μM WP9QY pretreatment was able to not only antagonize 20 ng/mL TNF-α-stimulated caspase-3 activation and but also reduce bax/bcl-2 ratio. The influence of 20 μM WP9QY was more evident. Thus, through the use of WP9QY to inhibit endogenous TNF-α, BLP-induced cardiomyocyte apoptosis was obviously weakened, which suggested that endogenous TNF-α played an important role in cardiomyocyte apoptosis. Meanwhile, in this study, we observed that effects of BLP-induced apoptosis had not been completely inhibited by WP9QY. There are several possible reasons: first, WP9QY is a kind of competitive inhibitor that cannot fully antagonize effects of endogenous TNF-α. Second, except for TNF-α, BLP could induce other inflammatory cytokines or reactive oxygen species to induce cardiomyocytes apoptosis. Third, BLP possibly has direct pro-apoptotic effect on cardiomyocyte apoptosis not via inducing TNF-α production. Influence of endogenous inflammatory cytokines on heart function has hardly been discussed by other scholars.17,18 This study possibly declared another important mechanism in sepsis-induced cardiac dysfunction.
IL-10, a pleiotropic cytokine, is able to suppress a variety of inflammatory factors production, such as TNF-α and IL-1β. IL-10 knockout endotoxemic mice showed excessive inflammatory response, 19 indicating that IL-10 played an important role in resistance to infection. Our preliminary study 4 found that use of low-dose BLP to induce septic mice immune tolerance can significantly improve the heart function and observed to have high level of IL-10 in heart homogenate, suggesting that IL-10 may participate in improvement of sepsis-induced cardiac dysfunction. But, the specific mechanism of IL-10 is not clear. The present results showed that no matter whether IL-10 was pretreated or post-treated, it was able to reduce BLP-induced cardiomyocyte apoptosis. In a recent study, 20 intravenous injection of IL-10 improved left ventricular function followed by a decrease of pro-inflammatory cytokines and infiltration of macrophages in rats subsequent to myocardial infarction. However, the biological effects of IL-10 for clinical prevention and treatment of sepsis-induced cardiac dysfunction need more researches.
In summary, this study demonstrated that both BLP and TNF-α could induce neonatal rat cardiomyocyte apoptosis. Endogenous TNF-α generated from cardiomyocytes themselves played an important role in bacterial lipoprotein-induced cardiomyocyte apoptosis. In addition, IL-10 was found to have anti-apoptotic property to antagonize cardiomyocyte apoptosis. This study provided experimental evidence for prevention of sepsis-induced cardiac dysfunction.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was supported by the Program for Development of Innovative Research Team in the First Affiliated Hospital of Nanjing Medical University (no. 20113012).