
Abstract
Advanced glycation end products (AGEs) are a family of compounds of diverse chemical nature that are the products of nonenzymatic reactions between reducing sugars and proteins, lipids, or nucleic acids. AGEs bind to one or more of their multiple receptors (RAGE) found on a variety of cell types and elicit an array of biologic responses. In this review, we have summarized the data on the nature of AGEs and issues associated with their measurements, their receptors, and changes in their expression under different physiologic and disease states. Last, we have used this information to prescribe lifestyle choices to modulate AGE-RAGE cycle for better health.
‘Exogenous AGEs [advanced glycation end products], for the most part, are derived from food that we consume.’
Advanced Glycation End Products: Origin and Metabolism
Advanced glycation end products (AGEs) are a family of compounds that are the products of nonenzymatic reactions between reducing sugars and proteins, lipids, or nucleic acids.1-4 Roasting and broiling food at high temperatures is a common practice in cooking. These high temperatures facilitate chemical reactions between primary and secondary amino groups of amino acids in proteins and carbonyl groups of reducing sugars, resulting in formation of AGEs; this reaction is commonly referred to as Maillard reaction. 5 While there are at least over 3 dozen known AGEs, only about half of these have been identified in foods. Some typical AGEs in food include, Nϵ-carboxymethyl-lysine (CML), Nϵ-carboxyethyl-lysine (CEL), pyrraline, crossline, pentosidine, imidazolium cross-link derived from glyoxal and lysine-lysine (GOLD), and imidazolium cross-link derived from methylglyoxal and lysine-lysine (MOLD). 6 These compounds play a very important role by giving special aroma, color, and taste to different foods.7-9
AGEs are also produced in vivo as a result of normal metabolic processes, or can come from diet. Methylglyoxal (MG) is the most common endogenous mediator of AGEs synthesis that is present ubiquitously in all cells. MG is largely derived as a result of carbohydrate, lipid, or amino acid metabolism involving both enzymatic and nonenzymatic reactions.10-13 MG synthesis is catalyzed by MG synthase, cytochrome P450 2E1, myeloperoxidase, and amino oxidase, participating in glycolytic bypass, acetone metabolism, and amino acid breakdown, respectively. 12 The nonenzymatic pathways of MG synthesis include the spontaneous decomposition of dihydroxyacetone phosphate, the Maillard reaction, the oxidation of acetol, and lipid peroxidation. 12 Exogenous AGEs, for the most part, are derived from food that we consume. The total body AGEs burden is the sum total of AGEs from dietary sources and endogenous synthesis. However, the relative contribution of endogenous versus exogenous AGEs in determining total body AGEs burden and its physiological relevance is difficult to assess. This is due to the multiplicity of AGE molecules, their differing biopotencies, and lack of reliable data on their metabolism, absorption, and distribution in body compartments.14-24
AGEs in Health and Disease
Interest in the role of AGEs in health and disease was sparked by initial reports of progressive rise of in vivo AGEs with normal aging,25-27 by their ability to cross-link proteins in an irreversible fashion,28-30 and by their modulation of extracellular-signal-regulated kinases (ERK) signaling.31-36 A series of reports demonstrating rise in circulating AGEs in people with diabetes37-41 and chronic kidney disease42,43 stimulated further interest in health implications of AGEs. This followed a plethora of empirical studies exhibiting an association between AGEs and a variety of conditions such as decline in memory with age,44-46 pathophysiology of eye diseases,47-49 polycystic ovary syndrome,50-55 wound healing,56-61 cardiovascular complications,62-68 bone health,69-71 periodontitis,58,72 erectile dysfunction,73-76 anemia in older community-dwelling women, 77 slow walking speed in older adults, 78 peripheral neuropathy,79,80 peripheral artery disease,81-85 obstructive sleep apnea,86-89 islet β-cell dysfunction,90-96 cancer,97-99 elevated cellular oxidative and inflammatory state,100-105 schizophrenia, 106 Alzheimer’s disease,107-113 and risk for metabolic syndrome in adults and children.114,115
In a recent study, Foster et al examined racial (European ancestry vs African American) differences in serum level of AGE by ELISA and prostate tissue expression of RAGE (receptor for AGE) by immunocytochemistry in subjects who underwent surgery for prostate cancer. 97 The results of this study showed higher expression of both AGE and RAGE in African Americans compared with those of European ancestry, suggesting the possibility that AGE-RAGE axis may be a marker for cancer health disparity. 97 The relationship between AGE and its receptors is described in detail under “The AGE-RAGE Cycle” section. While most of these empirical studies do not address a cause-and-effect relationship between AGEs and a disease condition, there are observations supporting altered physiologic responses following administration of exogenous AGEs, foods rich in AGEs, or perturbation of their endogenous levels.116-128 Some examples of such observations from animal and human studies include reproductive abnormalities and prostatic disorders in mice, 116 induction of inflammatory mediators,116,117 promotion of insulin resistance in mice following oral administration of AGEs, 119 acute state of impaired endothelial function, 120 acute impairment of vascular function after a high AGEs meal, 121 enhancement of low-density lipoprotein (LDL)–induced vascular toxicity by high AGEs diet, 122 increased proteinuria,123,124 and increase in lung level of high mobility group box protein 1 (HMGB1). HMGB1 is a nuclear protein that acts like an agonist for RAGE. 125 In a recent pilot study, we examined whether the effect of dietary AGE on circulating AGE may be controlled by fat content (low-fat vs high-fat) of the diet. 126 In this study, CML was measured as a surrogate marker of AGE. 126 Results of this study show that while there was no change in serum CML following consumption of a low-fat, high-AGE breakfast, there was a small but significant rise in CML after the high-fat, high-AGE breakfast. These data are suggestive that perhaps high dietary fat may increase health risk associated with AGE. A recent human study shows a very weak reduction in circulating AGE following 3 months of low-calorie Mediterranean diet. 127 In a double-blind, randomized, crossover human trial, a diet low in AGE resulted in improved insulin sensitivity and decreased urinary AGE excretion. 128
As described above, while there is a large volume of published data implicating the role of AGEs in a variety of chronic diseases, there are also publications that contradict many of the above observations.129-131 Data from the cross-sectional Reykjavik Study did not show any significant association between AGEs and age-related macular degeneration. 132 While evaluating the efficacy of Irbesartan in 450 patients with type 2 diabetes and nephropathy, Busch et al observed a lack of any role for AGEs in predicting cardiovascular events and renal outcomes in these patients, 133 an observation contrary to earlier reports.42,43,62-68 In another study, Somoza and colleagues observed a diet high in AGEs enhanced antioxidant capacity as well as chemopreventive enzymes—glutathione-S-transferase and UDP-glucuronyl-transferase, 129 again an observation contradicting a prooxidant role for AGEs.100-103 Another example is the observation that intake of bread crust rich in AGEs did not negatively affect calcium bioavailability and bone metabolism, 130 an observation unlikely to explain positive correlation between bone fracture and AGEs.69-71 A large randomized controlled trial that demonstrates no effect of dietary AGEs on endothelial function and inflammation 131 contradicts earlier reports associating AGEs with inflammation.104,105
While reasons underlying these apparent contradictory observations are not clear, few appear very apparent. AGEs refer to a collection of structurally different molecules that may have different affinity for receptors for AGE. Hence, there are possibilities for different physiological responses. Yet we almost always measure single chemical entities (eg, CML, CEL, GOLD, or pentosidine), or administer a single chemical entity and assume it to represent all AGEs. The measurements of AGEs employ multiple methods utilizing physicochemical (high-performance liquid chromatography [HPLC]) and immunological (enzyme-linked immunosorbent assay [ELISA]) on extracted as well as unextracted samples. The use of different methods will most certainly yield different values for AGE. The specificity and affinity of the antibody used as well as exposure of antigenic epitopes in samples affect the outcome of ELISA assay. These are some of the very obvious measurement-associated variables that may explain the observed differences in association studies. Additionally, choice of AGEs (eg, CML vs pentosidine vs CEL) for in situ, in vivo, or in vitro studies may yield different results. In many association studies, skin autofluorescence has been used as a surrogate measure of AGEs39,42,70,81; however, almost none of these studies has ever ascertained whether skin autofluorescence correlates with circulating AGEs. Furthermore, it is well known that only select AGEs produce autofluorescence. 7 The observation of lack of correlation between skin autofluorescence readings and circulating AGEs in patients with systemic lupus erythematous 134 further emphasizes the need for validation. This line of thinking is supported by the observation from Stirban and colleagues where they reported postprandial increases in skin autofluorescence in both diabetic and healthy subjects. 135 Our lack of understanding of how AGEs interact with their receptors vis-è-vis how AGEs regulate their receptors further complicates the interpretation of such studies.
Since the ultimate goal is to manage levels and actions of AGEs for better health, the following section summarizes the processes and mediators involved in action(s) of AGEs. This would enable investigators to plan how AGEs or downstream mediators of AGEs and their actions could be managed.
The AGE-RAGE Cycle
There are 4 known receptors for AGEs: full-length RAGE, N-truncated RAGE (Nt-RAGE), and C-truncated RAGE, which has 2 isoforms, secretory RAGE (sRAGE) and endogenous secretory RAGE (esRAGE). RAGEs belong to a member of the immunoglobulin superfamily of receptors. 136 RAGE, an approximately 45-kDa protein, has an extracellular component consisting of a variable (V) immunoglobulin-like domain followed by 2 constant domains (types C and C′). 137 It has a single transmembrane domain followed by a cytosolic tail. 138 The N-terminus of the V domain is the ligand-binding site, and the cytosolic tail is essential for RAGE-induced intracellular signaling. 138 Nt-RAGE resides in the plasma membrane, but its function is poorly understood. However, expression of cellular Nt-RAGE, like other RAGE variants, is regulated differentially by different RAGE ligands.139-142 For example, receptor engagement by distinct AGEs (CML-HSA, MG-HSA, diabetic RBC) differentially enhances expression of RAGE isoforms (RAGE, Nt-RAGE) in HUVEC cells.143,144 C-truncated isoforms lack cytosolic and transmembrane domain and circulate in the blood. There are 2 isoforms of C-truncated RAGE: total sRAGE and esRAGE. sRAGE is formed by ectodomain shedding of RAGE catalyzed by membrane-bound proteases including matrix metalloproteases and the closely related ADAM (a disintegrin and metalloprotease domain).141-143 esRAGE, a variant of RAGE lacking transmembrane domain, is formed from alternative splicing of native membrane receptor. 138 Serum levels of sRAGE are 5 times higher than esRAGE in healthy subjects. 145 Both sRAGE and esRAGE act as decoy for RAGE ligands by sequestering RAGE ligands or competing with full RAGE for ligand binding and thus have cytoprotective effects against AGEs-RAGE interaction. 146
RAGE is widely expressed in a variety of tissues (heart, lung, skeletal muscle, and vessel wall) and cell types (neurons, microglia, astrocytes, cerebral endothelial cells, pericytes, smooth muscle cells, monocytes/macrophages, and lymphocytes).138,147 It is a pattern recognition receptor. 146 and it has a large repertoire of ligands, enabling it to participate in the etiology of chronic diseases associated with cellular stress and inflammation. Some of these ligands include AGEs; amyloid-β peptide (which accumulates in Alzheimer’s disease); amyloid A (which accumulates in systemic amyloidosis); S100/calgranulins, a family of closely related calcium-binding polypeptides that accumulate extracellularly at sites of chronic inflammation; DNA-binding protein HMGB1 (amphoterin), which is released by cells undergoing apoptosis; and surface molecules on bacteria and leukocytes (macrophage-1 antigen or Mac-1).138-140 Binding of AGEs to RAGE regulates transcription factors, such as nuclear factor kappa B (NF-kB), activator protein 1 (a Jun-Jun homodimer or a Jun-Fos heterodimer), and forkhead box protein O4 via various signal transduction cascades, such as mitogen-activated protein kinases (MAPK), c-Jun N-terminal kinases, extracellular signal-regulated protein kinases 1 and 2, and Janus kinase/signal transducers and activators of transcription (JAK-STAT).148-153 It is noteworthy to mention that the common cellular response associated with all of these signal transduction cascades is inflammation. As mentioned earlier, 2 isoforms of RAGE—sRAGE and esRAGE—bind to AGEs but fail to initiate intracellular signal transduction cascade and, therefore, elicit a protective anti-inflammatory response.154,155
Like sRAGE and esRAGE, there are other receptors that bind AGEs but do not transduce cellular signals; thus, physiologically they serve as antagonists of AGEs action. 155 Such receptors include macrophage scavenger receptor types I and II (SR-A), 155 oligosaccharyl transferase-4 (OST-48 or AGE-R1), 156 galactin-3 (AGE-R3), 157 protein kinase C substrate (AGE-R2), 158 and lectin-like oxidized LDL receptor-1 (LOX-1).159-162 AGE-R1 or OST belongs to a family of proteins complexes (commonly known as translocon) responsible for the translocation of polypeptides across membranes in eukaryotes. It is a single transmembrane protein that has a small extracellular N-terminal domain and a cytoplasmic C-terminal domain. 163 AGE-R2, an 80 to 90 kD protein containing a tyrosine-phosphorylated section anchored in the plasma membrane of the cell, participates in the intracellular signaling of various receptors, like the fibroblast growth factor receptor. 158 AGE ligands bind at the C-terminus of AGE-R3 with high affinity. 157 AGEs also bind to the class E scavenger receptor, LOX-1, and have been shown to increase LOX-1 expression in diabetic rats.159-162
The Lifestyle Choices Affecting AGE-RAGE Cycle
Lifestyle choices have a significant effect on total body AGEs load, expression/action of RAGE and its isoforms, and resulting metabolic consequences. Figure 1 summarizes effects of lifestyle on AGE-RAGE cycle and plurality of pathophysiologic consequences. Summarized below are the role of lifestyle choices such as smoking, exercise, diet, and dietary supplements on the status of AGE-RAGE cycle and its consequences.
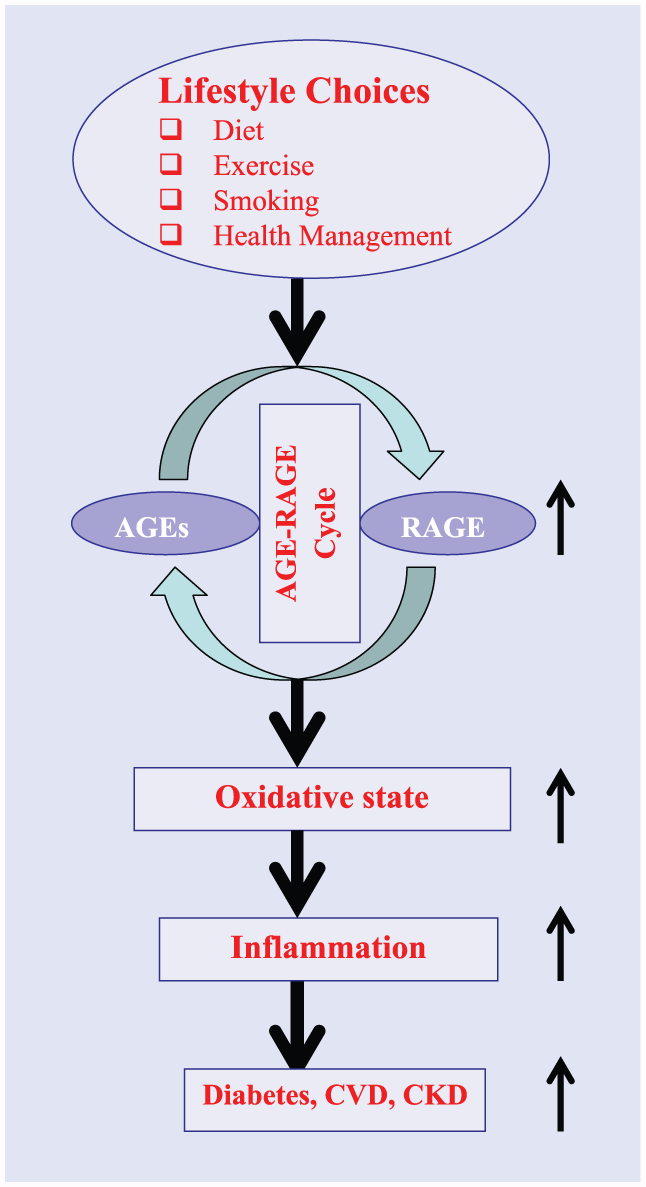
A schematic representation of possible steps between lifestyle choices and emergence of chronic diseases.
Smoking
Certain components of cigarette smoke can react with plasma and extracellular matrix proteins to form covalent adducts with many properties similar to AGEs.164,165 This has been suggested as a possible explanation for higher incidence of cardiovascular disease and cataracts in smokers than in nonsmokers. 165 This is further supported by observations that in smokers, tobacco-derived AGEs accumulate in plasma LDL, structural proteins present within the vascular wall, the lens proteins of the eye, and the collagen in the skin.165,166
Exercise
The data on the regulation of AGE-RAGE cycle by exercise in human remains controversial. Aging rats on an exercise regimen have shown a decrease in circulating AGEs with concomitant attenuation of cardiac fibrosis. 167 These observations were further supported by another animal study in which Steppan et al suggested that exercise combined with Alagebrium (an AGEs breaker) prevented formation of new AGEs as well as breakdown of already formed AGEs. 168 This may represent a therapeutic strategy for age-related ventricular and vascular stiffness. 168
In a recent human study, 17 middle-aged sedentary nonsmoking healthy females (35-70 years) free of overt metabolic, cardiovascular, and renal disease underwent a 3-month-long lifestyle modification program that included educational sessions for healthy eating and exercise. At the end of the study, there was a significant decrease in serum levels of CML and pentosidine. 169 However, other human studies where measured outcomes were sRAGE or esRAGE, known to attenuate action of AGEs, have yielded conflicting results. 170 For example, Choi et al reported aerobic exercise to increase sRAGE levels along with improvement of various cardiometabolic risk factors in patients with type 2 diabetes. 171 These observations were further supported by Santilli et al, who reported a significant increase in plasma esRAGE following an 8-week standardized aerobic high-amount–high-intensity training program in 22 sedentary human subjects. 170 In contrast, Kotania et al found a decrease in plasma sRAGE with increase in physical activity in an elderly population. 169 It is conceivable that these results might have suffered from the fact that the sample size was very small and there was no adequate description of subjects recruited for this study. 169 This observation warrants further studies on the clinical relevance of sRAGE changes with physical activity.
Diet
Western diet is high in AGEs. Two large studies have attempted to quantify AGEs in a variety of foods.4,172 These studies were performed by the same group using the same method (ELISA with anti-CML antibody). Another smaller study by Hull et al used HPLC-mass spectrophotometry (MS) to determine CML concentrations in foods. 173 In most studies, CML has been the surrogate marker to estimate AGE content of foods. This is based on earlier studies indicating that CML levels directly correlate with levels of other protein- or lipid-derived AGEs. 174 Reliability of measures of AGEs in foods is complicated by lack of specificity of measurements and methods used to calculate their concentrations.4,172
Many recent dietary intervention studies have used the tables developed by Goldberg et al to calculate AGE in their test diets rather than measuring AGE in the test foods. 4 To develop this table, Goldberg et al obtained a variety of foods from convenience stores and fast food restaurants and also prepared foods using standard cooking times with variation in cooking methods: boiled, broiled, deep fried, oven fried, and roasted. All foods were analyzed for AGE using ELISA that quantifies CML. However, this study did not report whether multiple samples/trials were performed in order to determine AGE. By using this method, concentrations of AGEs were lowest in carbohydrate foods with the lowest levels within this group being found in milk, followed by vegetables and fruits. Broiled beef, chicken, oils heated at high temperature, and roasted nuts were among the highest CML foods. Using these charts the researchers analyzed 3-day food records from healthy participants and found mean daily AGE intake to be about 16 000 ± 5000 kU AGE. 4
A follow-up study by the same group repeated the study using greater variations in cooking techniques—marinating, various temperatures, use of AGE inhibitors—and reported that fats have higher levels of AGEs per gram. 172 Beef and cheese were found to have the highest levels of AGEs followed by poultry, pork, fish, and eggs. In addition, higher fat and aged cheeses were found to have more AGEs than lower fat cheeses. 172
The study by Hull et al, which used HPLC-MS to determine CML in foods, found that AGEs were highest in cereal and lowest in fruits and vegetables expressed as mg/100 g of food. CML remained highest for meat products when expressed as milligram per serving. However, levels of CML in oils were very low. 173
An analysis of European liquid infant formulas found elevated concentrations of various AGEs including CML with 3- to 8-fold higher concentrations in liquid infant formula compared with cow’s milk. In the same study, the level of CML in powder infant formula was 2.5 to 5 times higher in concentration than standard powdered milk. 175 Also, the levels of CML in infant formula have been found to be 70 times higher compared with breast milk. Plasma levels of CML in infants were also consistently higher in the formula-fed than those who were breast-fed. 176
A study of CML levels in a variety of common foods published in 2009 found it to vary from as low as 0.3 mg/kg of raw milk and 0.35 mg/kg of skim milk to as high as 46.1 mg/kg of whole meal bread crust (compared to just 4.45 mg/kg of bread crumb). 177 Commercial breakfast cereals, ice cream, and barbecue sauces also appear to be sources of AGE.178-180 An evaluation of a variety of processing methods for nuts and seeds found that CML levels were increased by roasting. 181 Thus, consumption of cooked foods compared to raw foods increases AGE ingestion. 181 While low-fat vegan diets are lower in AGEs, the research indicates that plasma AGE is actually higher in vegans. 182
Intermediate glycation metabolites which may or may not form AGEs such as MG and glyoxal have also been evaluated in various foods. In general, products containing high fructose corn syrup were found to be higher in MG in comparison to diet drinks. 183 In contrast, a study by Uribarri et al found high quantities of MG in diet coke and low MG in regular Coke and low quantities of CML in both. 172 For Pepsi, the reverse was true. 172 Drinks with caramel additives such as Coke Classic or Diet Coke were found to contain 8500 and 9500 units/cup compared to 475 units per cup in Sprite, 600 in orange juice, and around 2000 in coffee and tea. 20 The problem with these studies is that they vary in the method of quantifying AGE—some quantify CML versus other AGE products. There are multiple concerns with the reported values of AGEs. These include the following: what is measured, how is it measured, and how much is absorbed.4,172,173,177-180,184
Dietary Supplements
Steps associated with synthesis and breakdown of AGEs is summarized in Figure 2. The major goals of research in the use of dietary supplements including herbals in regulating AGEs are to either block/delay its synthesis or enhance degradation of existing AGEs. These studies can generally be divided into 3 types in order of preponderance in published literature: (1) in vitro and cell culture studies, (2) animal studies, and (3) human studies. In in vitro screening, glycation reaction is initiated by incubating high concentrations of a reducing sugar (hexose or pentose) with a model protein (eg, lysozyme, bovine serum albumin, collagen type 1). The reaction is slow and it takes days to weeks to plateau. Test substances (supplements or plant extracts) are added to the incubation mixture and anti-glycation activity is evaluated by assessing nature of model protein used by sodium dodecyl polyacrylamide gel electrophoresis (SDS-PAGE), ELISA, and HPLC.
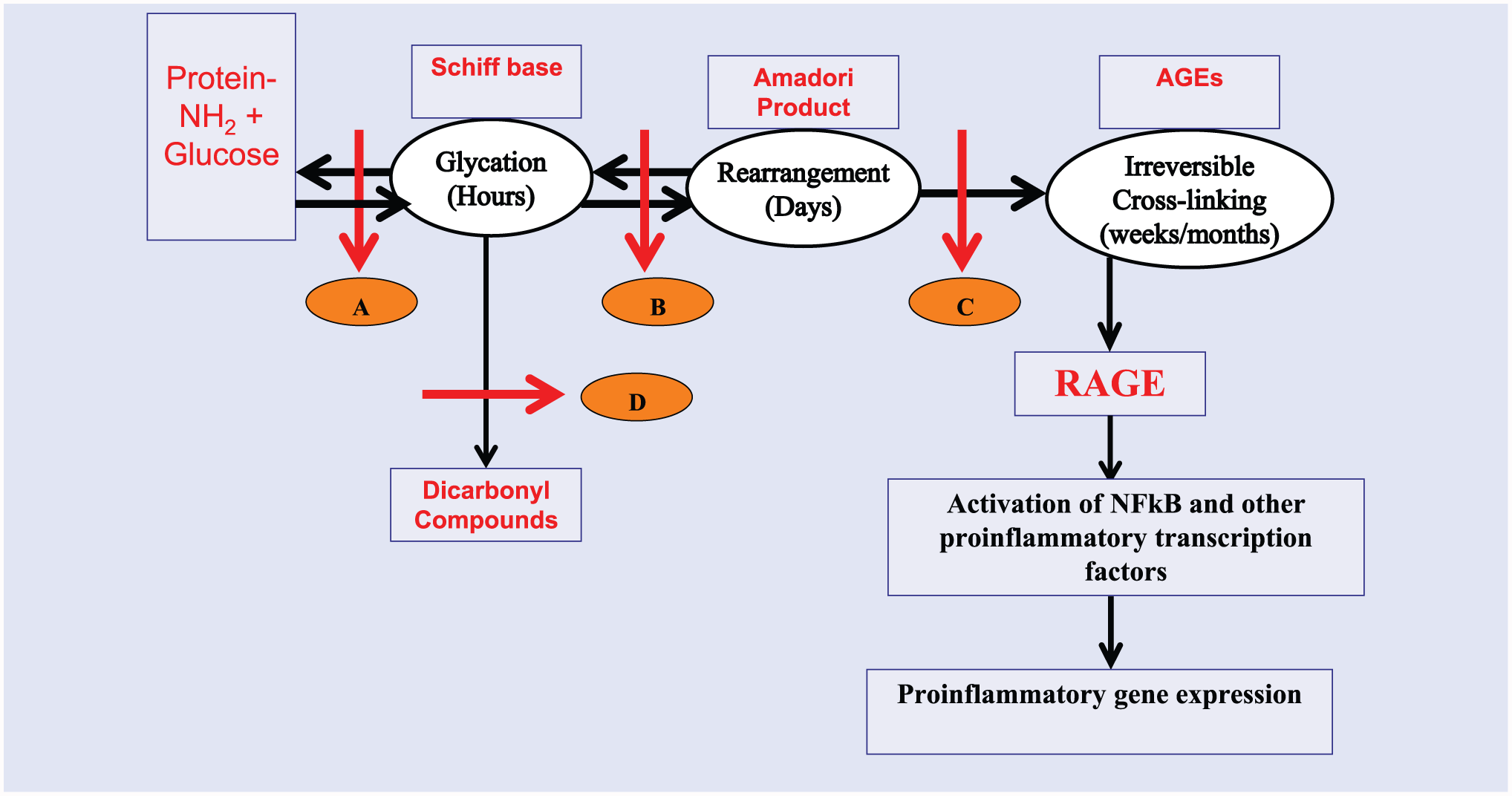
A diagrammatic representation of steps in the formation of irreversible advanced glycation end products. Letters A to D denote sites where AGEs formation could be intervened through lifestyle changes or use of pharmacologic agents. A, Schiff’s base formation leading to glycation; B, Rearrangement of glycated proteins into Amadori adducts; C, Cross-linking of Amadori adducts into irreversible AGEs; D, Conversion of glycated products into reactive dicrbonyl compounds.
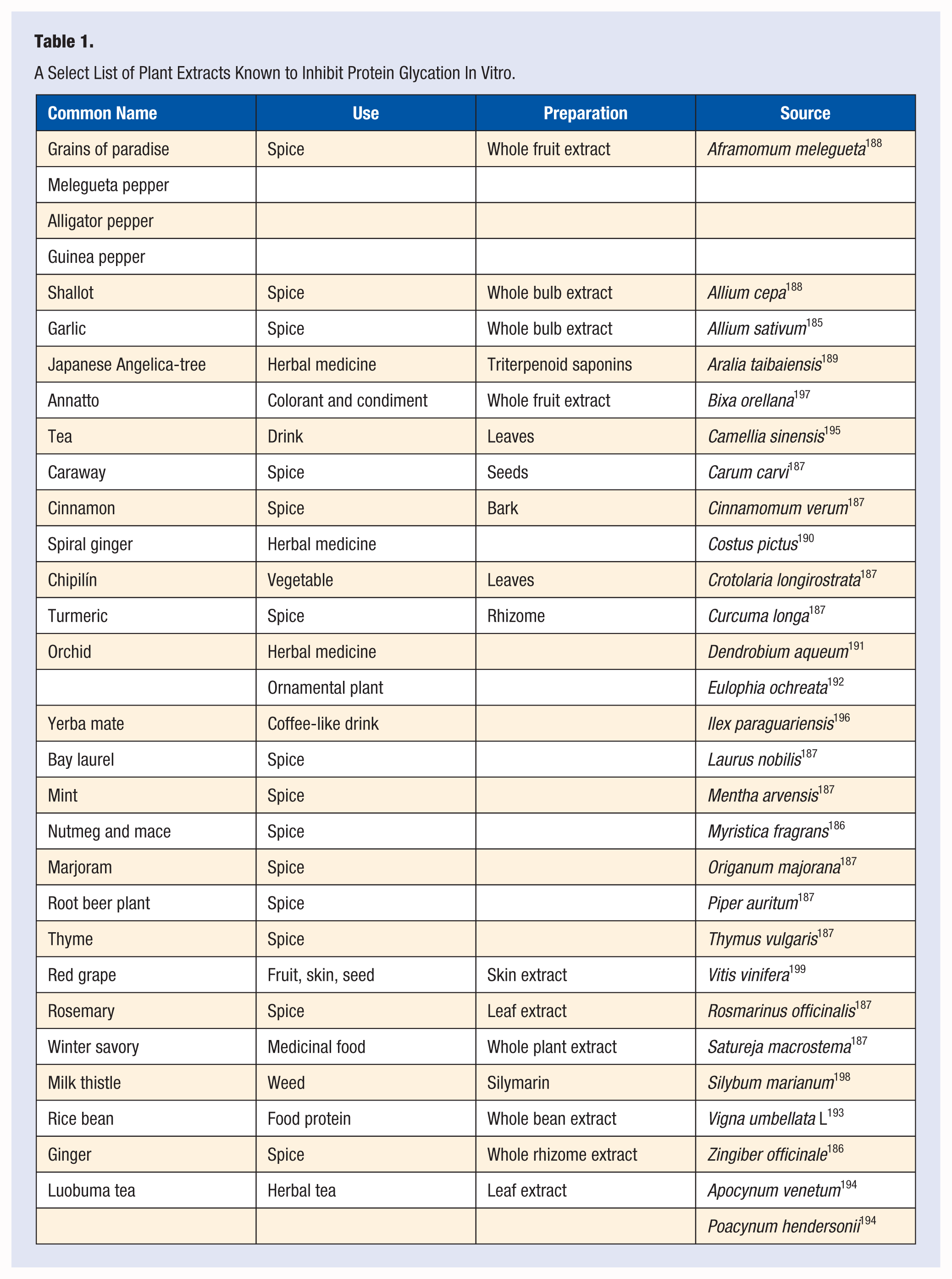
Multiple investigators have tested a variety of dietary supplements and aqueous or organic extracts of medicinal and food plants for their ability to inhibit protein glycation in in vitro or in animal studies (Table 1). While these observations are interesting, establishing their relevance to health outcomes would require standardization of the extraction method, stability of bioactivity, evaluation of toxicity, and finally human studies to document safety and efficacy. Many of these extracts come from commonly consumed foods, and thus may have a lower possibility of toxicity. Unfortunately, most of these initial observations have not progressed to the next step in the process. In contrast to in vitro and animal studies, there are few cell culture studies.
200
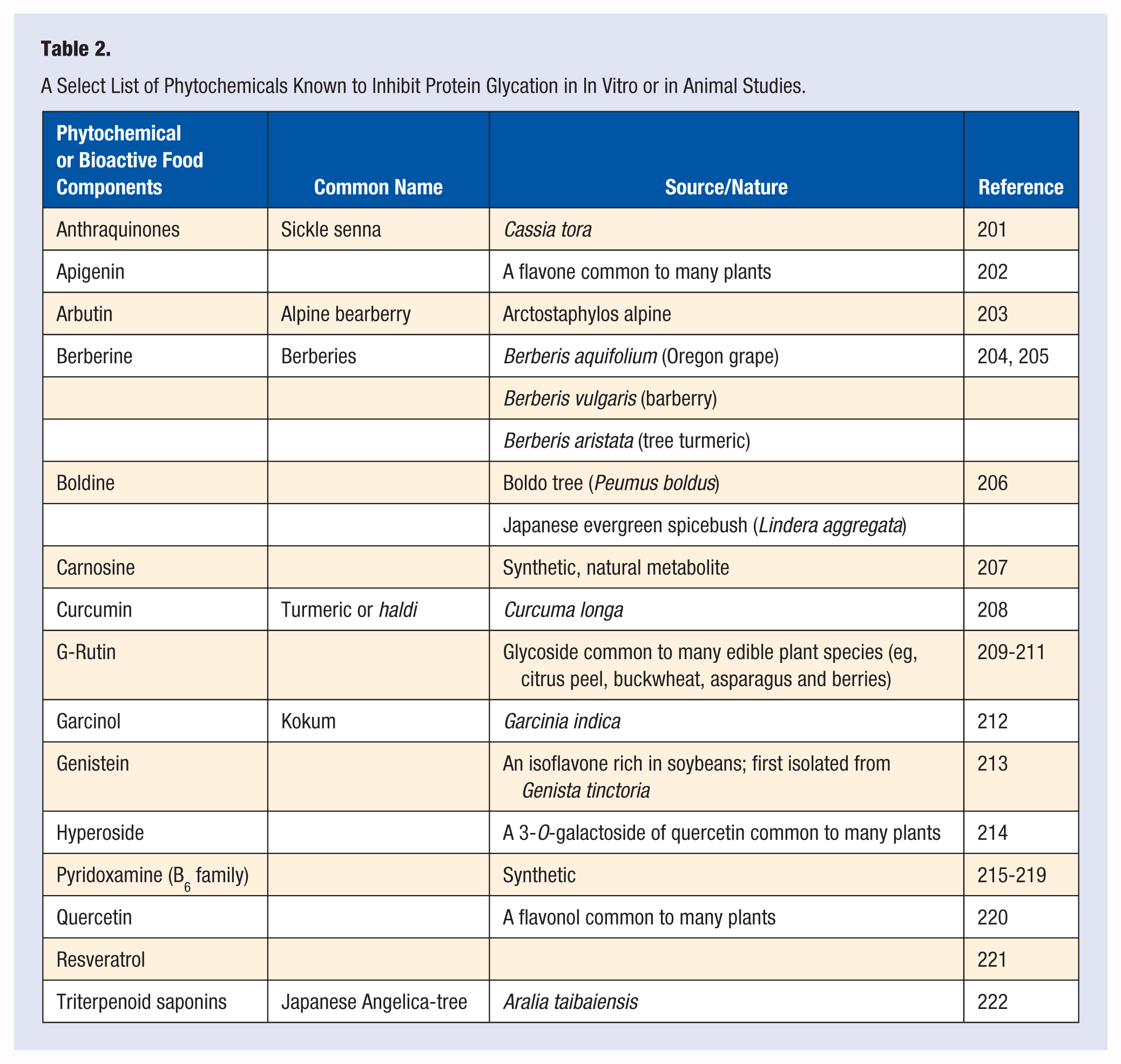
Encouraging results from the tests using crude preparations led many investigators to examine the effects of purified/semipurified plant components, bioactive food components, and their synthetic counterparts on protein glycation resulting in several candidate agents for possible human study (Table 2). Unfortunately, human studies to assess safety and efficacy of such preparations are far fewer.
223
Of the agents tested so far in human studies, pyridoxamine and thiamin—2 B-vitamins—have provided clinical data of uncertain value.215-219 Pyridoxamine, for example, has no established safe human dosage, and at high dosage, it produces profound sensory loss and sensory neuropathy with axonal degeneration.
224
Benfotiamine, a synthetic S-acyl derivative of thiamin (vitamin B1) widely used in Germany for sciatica and other painful nerve conditions, has shown inconclusive results in every study when glycation-related outcomes were evaluated.225-228 There have been a series of human studies examining role of aged garlic in managing coronary artery calcification.229-231 The results of these studies show aged garlic alone or in various combinations with B vitamins, folic acid, coenzyme Q10, and
A Select List of Plant Extracts Known to Inhibit Protein Glycation In Vitro.
A Select List of Phytochemicals Known to Inhibit Protein Glycation in In Vitro or in Animal Studies.
Pharmacologic Interventions
Although a discussion on the development and use of pharmacological agents for improving glycation-related outcomes is beyond the scope of this review, a brief overview of our experience with a few compounds studied so far is worth mention. Aminoguanidine (trade name: pimagedine) is a diamine oxidase and nitric oxide synthetase inhibitor that reduces circulating AGEs. 232 A human trial of pimagedine was terminated in 2000 due to an unfavorable risk-to-benefit ratio.233,234 Alagebrium (trade name: ALT-711) was another compound that underwent clinical trial for glycation-related outcomes. While there were some interesting clinical outcomes,235-237 again this trial was terminated in 2007 due to unfavorable risk-to-benefit ratio. Metformin (biguanidine) is a first line agent for suppressing glucose production by liver in obese-overweight insulin-resistant patients that also decreases AGEs production perhaps secondary to decrease in plasma glucose.238,239
Conclusion: Where Do We Go From Here?
AGEs include a diverse group of a family of compounds (exceeding over 3 dozens) that are products of nonenzymatic reactions between reducing sugars and proteins, lipids, or nucleic acids. Except for methylglyoxal10,12,13—a product of normal metabolism—all other AGEs are exogenous in nature derived from food. Although there is a progressive rise of in vivo AGEs with normal aging,25-27 in healthy individuals, the in vivo homeostasis of circulating AGEs is maintained through regulation of renal clearance of AGE-peptides. 240
Results of almost all of the human studies examining role of AGEs in health and disease are limited by the fact that they are cross-sectional in nature with great variances in ethnicity, gender, age, sample size, and end point measurements (see Table 3 and 4). The interventional studies are limited by exposure duration of hours to 6 weeks at the most. It is difficult to imagine how a short duration may have a meaningful consequence to a chronic health condition such as cardiovascular disease or diabetes. However, with few exceptions, the results are qualitatively similar. These results are summarized as follows:
Administration of exogenous AGEs to healthy adults does not lead to increased inflammation or endothelial dysfunction.126,131,132
Elevated AGEs are associated with higher risk of all-cause mortality, severity of coronary atherosclerosis, and cardiovascular disease mortality as well as chronic kidney disease.25,43,66,67
Elevated AGEs are associated with cognitive decline in elderly. 44
Elevated AGEs are associated with metabolic abnormalities in polycystic ovarian syndrome.50,54
Exogenously administered AGEs have deleterious metabolic consequences in diabetic as well as patients with compromised renal function.117,120-122
A diet that is low in AGEs may reduce the risk of type 2 diabetes by increasing insulin sensitivity. 128
A Summary of Human Studies Examining Role of Advanced Glycation End Products in Noncommunicable Diseases.
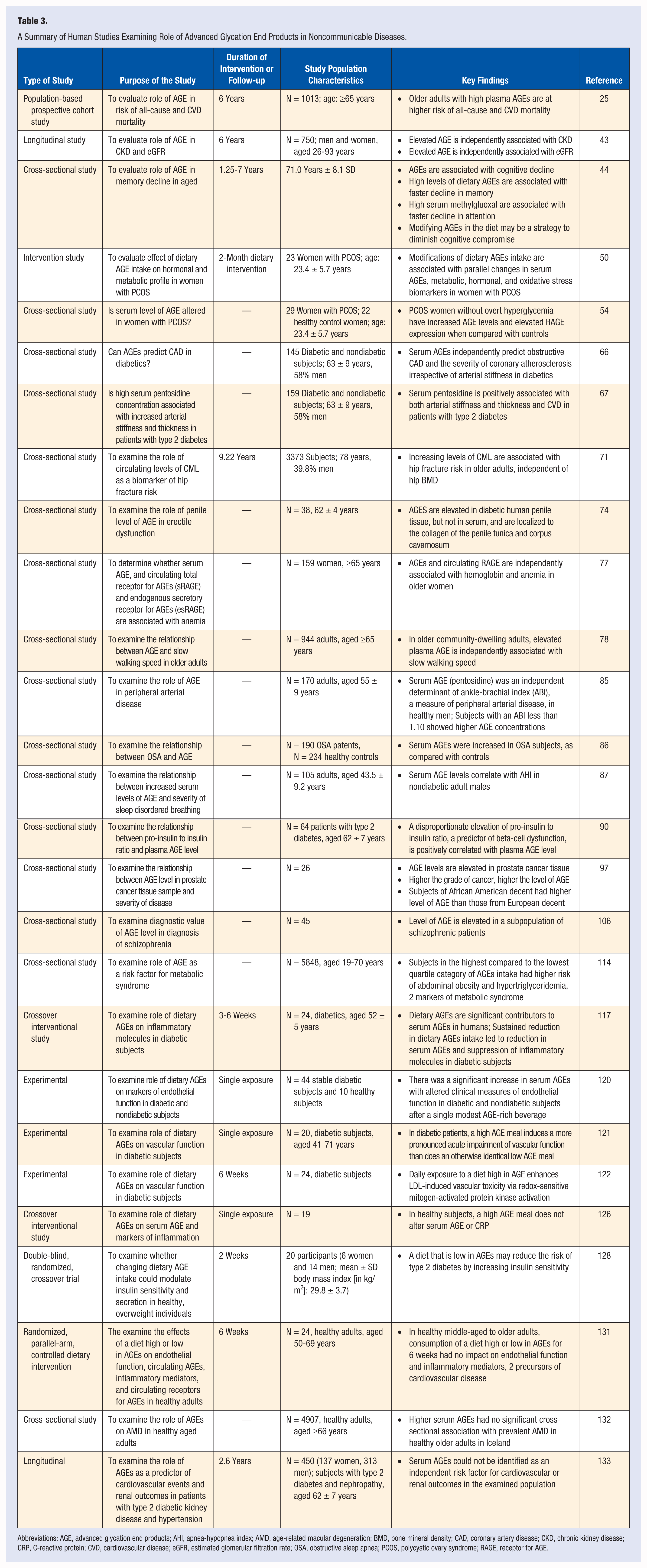
Abbreviations: AGE, advanced glycation end products; AHI, apnea-hypopnea index; AMD, age-related macular degeneration; BMD, bone mineral density; CAD, coronary artery disease; CKD, chronic kidney disease; CRP, C-reactive protein; CVD, cardiovascular disease; eGFR, estimated glomerular filtration rate; OSA, obstructive sleep apnea; PCOS, polycystic ovary syndrome; RAGE, receptor for AGE.
Summary of Animal Studies on the Impact of Advanced Glycation End Products in Noncommunicable Diseases.
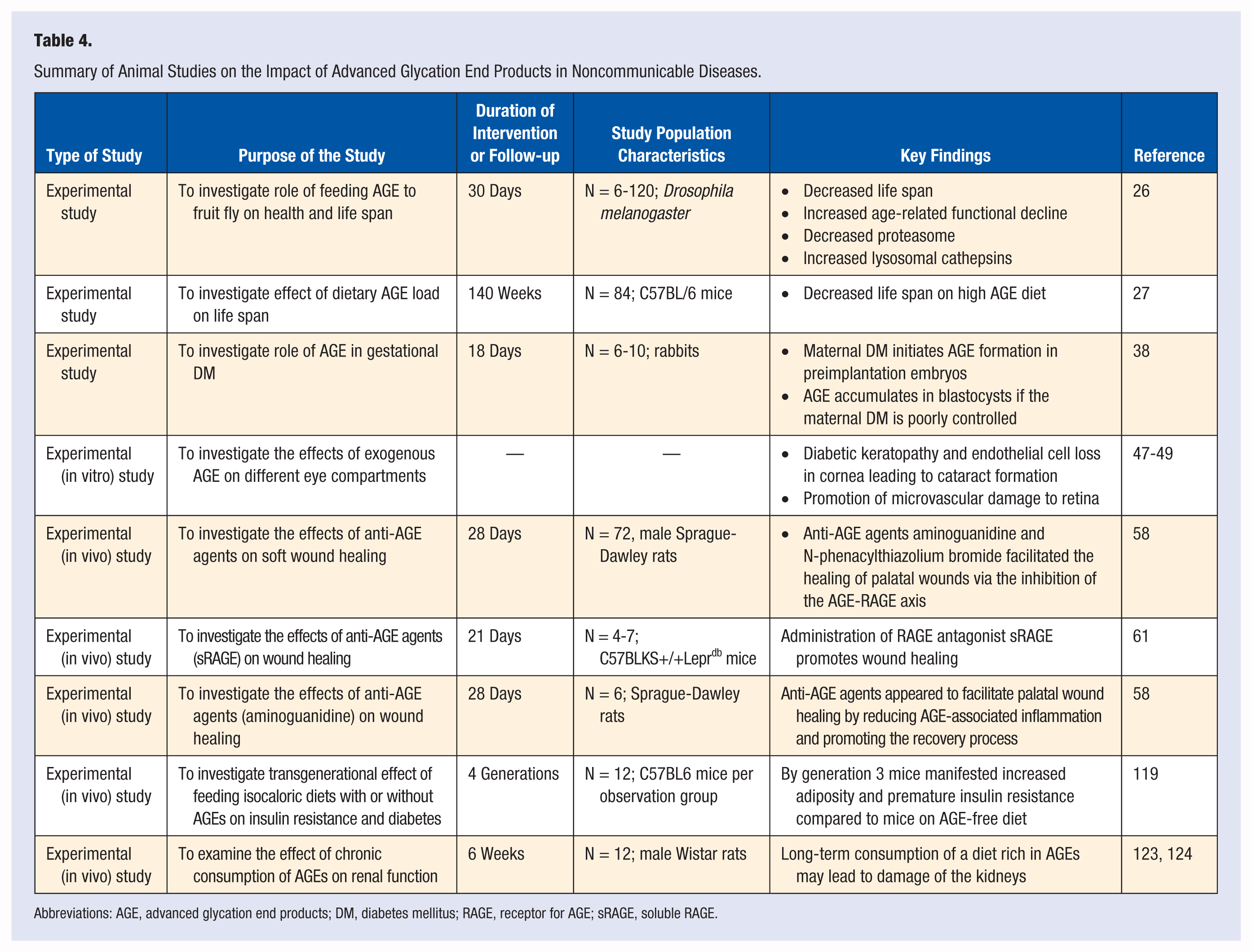
Abbreviations: AGE, advanced glycation end products; DM, diabetes mellitus; RAGE, receptor for AGE; sRAGE, soluble RAGE.
Irrespective of the role of AGE in initiation or progression of metabolic disorders, reduction in its level is certainly of health benefit. This has sparked great interest in defining ways to reduce circulating AGE levels. Approaches to reduction in deleterious effects of AGE include reduction in its formation, inhibition of its action, or increase in its metabolism by pharmacologic and dietary means. Pharmacologic agents, no matter how effective, generally, if not always, tend to carry undesired side effects. Therefore, approaches that may use dietary supplements or dietary modification in achieving a reduction in level or effect of AGEs would be preferable. In this review, we have summarized data on biology and chemistry of AGEs as well as surveyed available data on modification of its formation and action by lifestyle changes. Finally, we have made some lifestyle change recommendations to manage glycation load (Table 5).
Lifestyle Modification Recommendations to Lower Glycation Overload.

Footnotes
Acknowledgements
We thank Ms Amanda Bertucci (MS student) for help with literature search.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by The Human Nutrition Research Fund, the State of Texas.