
Abstract
Ionizing radiation, though indispensable in medicine, energy, and space exploration, imposes complex biological stresses that disrupt redox homeostasis, inflammation control, and genomic integrity, ultimately contributing to fibrosis, immunosuppression, and carcinogenesis. Despite advances in pharmacological radioprotectors and technological safeguards, their incomplete efficacy and safety concerns highlight the need for integrative and sustainable mitigation strategies. Emerging evidence supports that nutrition- and lifestyle-based interventions can attenuate radiation-induced injury and enhance intrinsic defense mechanisms without compromising therapeutic efficacy. Nutritional modulation through balanced energy intake, micronutrients, and bioactive compounds such as antioxidants, vitamins, omega-3 fatty acids, and phytochemicals helps stabilize redox balance, suppress inflammatory signaling, and promote DNA repair and mitochondrial stability. Micronutrients including selenium and zinc stimulate antioxidant enzymes and DNA repair systems, while phytochemicals such as resveratrol, curcumin, and epigallocatechin gallate activate Nrf2 and inhibit NF-κB pathways, restoring cellular homeostasis. Complementary lifestyle factors including regular physical activity, adequate sleep, circadian rhythm alignment, and stress management, further enhance immune surveillance, genomic stability, and metabolic adaptation. Collectively, these integrative approaches provide synergistic, multi-pathway protection against radiation-induced damage. Future directions should focus on multi-omics–guided mechanistic studies and well-designed clinical trials to establish personalized, evidence-based frameworks for radiation mitigation.
Keywords
Introduction
Background of Ionizing Radiation and Its Health Risks
Radiation plays a pivotal role across multiple domains, such as medicine, research, and industry, driving technological and scientific progress in areas including medical imaging, cancer therapy, and energy generation. While non-ionizing radiation (including visible light, infrared, radiofrequency, and microwaves) lacks sufficient energy to remove electrons from atoms, ionizing radiation (IR) possesses enough energy to generate ions and reactive free radicals capable of disrupting molecular stability and cellular integrity. 1 Sources of IR include radioactive isotopes, medical imaging devices, radiotherapy (RT) equipment, and nuclear reactors, among others. Although indispensable for diagnostic and therapeutic purposes, IR exposure remains a public health concern because of its capacity to induce molecular, cellular, and tissue-level injury. The scientific community continues to investigate both the beneficial applications of IR and its biological hazards stemming from medical, environmental, and occupational exposures. 2 Despite the development of numerous anthropogenic radiation sources, natural background radiation remains the principal contributor to total human exposure. It comprises three major components: terrestrial radiation, emitted by naturally occurring radionuclides in the soil and rocks; cosmic radiation, originating from high-energy particles from outer space; and internal radiation, which arises from the incorporation and decay of naturally occurring radionuclides within biological tissues.2,3 The biological effects of IR, whether electromagnetic waves such as X-rays and gamma (γ) -rays or particulate for instance alpha (α), beta (β) particles, and neutrons, depends largely on the radiation type, its linear energy transfer (LET), and tissue penetration depth. Owing its strong ionizing capacity, IR can eject electrons from atoms, break covalent bonds, and induce cascading molecular and biochemical alterations. 4
Among the radiation types, α particles exhibit the highest ionizing potential but limited penetration. Even a single α-particle can deposit substantial energy within a single cell. 5 β radiation, composed of electrons or positrons, has lower ionization density but greater penetration range. In contrast, γ radiation, with its high penetration power and low LET, can traverse the entire thickness of biological tissues, delivering diffuse energy throughout the body. 6 High doses, total-body or partial-body exposure may result in acute radiation syndrome (ARS), a potentially fatal condition characterized by nausea, vomiting, diarrhea, hematopoietic suppression, immune disfunction, and, at extreme doses, mortality. 7 Additional acute manifestations include cutaneous and soft tissue injury, epilation, and ocular complications such as radiation-induced cataracts, often observed following nuclear accidents, RT mishaps, or another occupational overexposure. 8 In contrast, chronic or stochastic effects arise from low-dose or prolonged exposure and may develop for years or decades. These include an elevated risk of carcinogenesis (e.g., leukemia and solid tumors), fibrotic and vascular degeneration, neurocognitive decline, and developmental abnormalities following prenatal exposure.9,10 IR is a well-established carcinogen, capable of inducing malignancies across nearly all tissues, species, and life stages. The latency period for radiation-induced leukemias typically ranges from 2 to 9 years, while solid tumors may emerge after 10–40 years post-exposure. 11 Furthermore, epidemiological data from the life span study of Japanese atomic bomb survivors revealed that even moderate or low radiation doses significantly increases the risk of cardiovascular diseases, including stroke and ischemic heart diseases. 12
Collectively, these findings underscore the dual nature of IR, as both a cornerstone of modern medicine and a pervasive environmental hazard, necessitating continuous evaluation of its biological impacts and the development of strategies to mitigate associated health risks.
Current Countermeasures: Pharmacological and Technological Approaches
Efforts to counteract the health risks posed by IR have largely advanced along two major avenues: pharmacological interventions and technological solutions. While both have made meaningful contributions to radioprotection and mitigation, their translation from preclinical success to clinical effectiveness remains limited, reflecting ongoing gaps in safety, cost-efficiency, and therapeutic breadth. Radioprotectors are administered prior to radiation exposure to prevent tissue injury by scavenging free radicals and shielding critical macromolecules from ionization events occurring during the early radiochemical phase. In contrast, radiomitigators are administered shortly after exposure, aiming to minimize toxicity and facilitate repair processes before overt clinical symptoms manifest. 13 These agents, collectively termed radiomodifiers, act through a spectrum of mechanisms, including antioxidant and free radical scavenging, modulation of cell death pathways, enhancement of DNA repair, suppression of inflammatory signaling, and stimulation of hematopoietic recovery. 14 Among pharmacological countermeasures, amifostine (WR-2721) remains the first and only Food and Drug Administration (FDA)-approved radioprotective agent with clinical validation. Initially developed under the U.S. Department of Defense’s radioprotectant program, amifostine functions as a cytoprotective adjuvant in RT and chemotherapy (CTx), notably reducing radiation-induced injury to the salivary glands, kidneys, and bone marrow. 15 However, despite its efficacy in mitigating hematopoietic injury, clinical adoption has been hindered by severe adverse actions, including transient ischemic attacks, hypersensitivity responses, nephrotoxicity, and cardiovascular complications. Moreover, its narrow therapeutic window, systemic toxicity, parenteral delivery requirement, and high cost have collectively constrained widespread use.16,17
Beyond amifostine, radiomitigators and immunomodulatory agents such as 5-androstenediol, granulocyte colony-stimulating factor for hematopoietic acute radiation syndrome, interleukin-6 (IL-6), CBLB502 (a truncated flagellin-derived Toll-like receptor 5 agonist), and angiotensin-converting enzyme inhibitors like captopril have shown efficacy in reducing tissue damage and improving survival in preclinical and early clinical studies.18,19 The U.S. FDA has approved ten pharmacological agents for mitigating radiation-associated injuries. Among these, amifostine and palifermin (keratinocyte growth factor) are approved for limited RT indications, though only amifostine qualifies as a true radioprotector, both constrained by toxicity and poor tolerability. 16 Additional cytokines such as erythropoietin, interleukin-3 (IL-3), and interleukin-11 (IL-11), initially developed for therapy-induced neutropenia, are under investigation as potential radiation countermeasures.17,20 Furthermore, many candidate compounds face challenges including adverse side effects, toxicity, unfavorable inability pharmacokinetics, to cross physiological barriers (e.g., blood-brain barrier), and substantial developmental and regulatory costs. 21 Consequently, ongoing research is directed toward developing next-generation countermeasures with improved safety profiles, broader radioprotective spectra, and optimized translational frameworks to bridge the gap between laboratory discovery and field deployment, while minimizing unintended toxicities.
Rationale for Exploring Nutrition and Lifestyle Interventions as Complementary Strategies
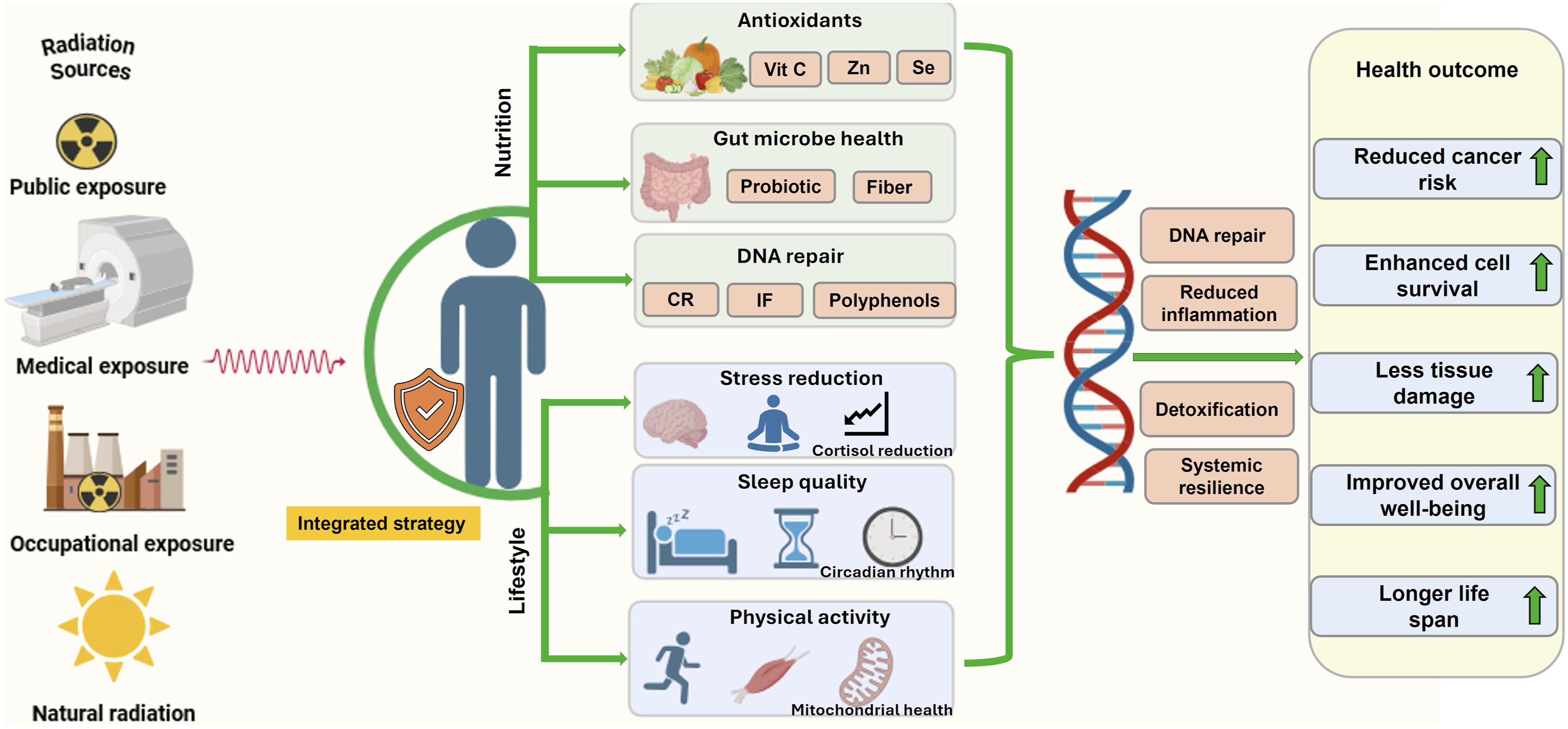
The limitations of existing pharmacological and technological countermeasures underscore an urgent need for safe, accessible, and low-toxicity alternatives capable of mitigating both the acute and delayed biological consequences of IR exposure. Increasing evidence suggests that micronutrients and bioactive natural compounds including polyphenols, antioxidants, phytochemicals, and trace elements, represent promising leads for next-generation radioprotective therapeutics due to their favorable safety profiles and capacity to modulate multiple biological pathways simultaneously. 22 Nutritional interventions generally exhibit excellent tolerability, can be administered orally, and are suitable for chronic or sustained dosing, making them advantageous over many pharmaceutical agents. Because they are derived from natural sources including foods, supplements, or plant extracts, nutritional radioprotectors tend to cause few adverse effects and do not require invasive delivery methods such as injections. Moreover, their wide natural availability and biological compatibility further enhance their translational potential as practical and sustainable radioprotective strategies. Maintaining a balanced macronutrient composition including appropriate proportions of carbohydrates, proteins, and fats, supports metabolic flexibility, enabling the organism to adapt to various physiological and oxidative stresses. 23 Micronutrients such as vitamins, carotenoids, probiotics, polyphenols, and Omega-3 polyunsaturated fatty acids (ω-3PUFAs) have demonstrated the ability to neutralize free radicals, enhance endogenous antioxidant defense, and stabilize redox homeostasis, thereby attenuating radiation-induced molecular and cellular damage.22,24 Beyond nutrition, lifestyle factors including regular physical activity, effective stress management, psychological well-being, adequate sleep, and circadian rhythm alignment play synergistic roles in fortifying the body’s natural defense systems. These interventions support metabolic efficiency, immune robustness, and neuroendocrine balance, which collectively reduce oxidative stress, enhance DNA repair, and may lower long-term cancer risk following radiation exposure. 25 Consequently, nutrition- and lifestyle-based interventions emerge as cost-effective, accessible, and multi-pathway adjunctive approaches for radiation risk mitigation. Recent studies indicate that these factors can substantially attenuate RT-induced toxicities, including site-specific inflammation, loss of lean body mass, and muscle strength deterioration, while improving treatment tolerance and recovery.22,26 Integrating tailored nutritional strategies with structured physical activity enhances overall physical fitness, while formal dietary guidance improves mental health, self-efficacy, and treatment adherence among patients undergoing cancer therapy. 27 Taken together, this combined and integrative framework supports the development of safe, sustainable, and multi-targeted approaches to protect both high-risk populations, such as RT patients, astronauts, and nuclear industry workers, and the general population from the health risk of IR.25,28 By reinforcing endogenous defense networks through natural physiological modulation, nutrition and lifestyle strategies offer a vital complementary dimension to conventional radioprotective paradigms.
Scope of This Review and Future Directions
This narrative review synthesizes recent advances in the integration of nutritional and lifestyle interventions for mitigating radiation-induced biological damage, emphasizing the underlying molecular mechanisms, key findings from experimental and clinical research, and the translational potential of these approaches in practical contexts. Despite encouraging progress, significant challenges persist, including an incomplete understanding of the mechanistic pathways, species-specific variability in experimental models, and limited translating success from preclinical data to human application. Furthermore, the scarcity of long-term, large-scale clinical trials continues to constrain definitive conclusions regarding efficacy, dosing, and population-specific recommendations, issues that are critically examined throughout this review.
Looking ahead, this review highlights the importance of multidisciplinary collaboration among the fields of radiobiology, nutrition science, pharmacology, and clinical medicine to bridge current knowledge gaps. Integrating pharmacological radioprotectors with personalized dietary strategies and evidence-based lifestyle interventions may yield synergistic, safe, and cost-effective countermeasures to enhance resilience against radiation-induced injury. Ultimately, such combined and holistic strategies hold promises for providing accessible, sustainable, and biologically sound solutions to safeguard both high-risk groups and the general population from the diverse health effects of IR.
Materials and Methods
This article was conducted as a structured narrative review aimed at synthesizing current mechanistic and translational evidence regarding the integration of nutrition- and lifestyle-based interventions in mitigating radiation-induced biological damage. A comprehensive literature search was performed across the following electronic databases PubMed, Scopus, Web of Science, and Google Scholar (for supplementary searches and cross-referencing) and included publications available up to December 2025. The core search terms included radiation mitigation; radioprotection; ionizing radiation; radiation-induced damage; nutrition, dietary intervention, micronutrients, antioxidants, caloric restriction, intermittent fasting on radioprotection; exercise, sleep, lifestyle modification on radiation response.
Peer-reviewed preclinical and clinical studies examining the molecular mechanisms of radiation-induced tissue damage (RITD) were included, with particular focus on dietary and lifestyle interventions such as polyphenols, circadian regulation, and physical activity. Studies lacking a clear mechanistic basis, non–peer-reviewed opinion articles, and interventions with high toxicity or limited translational relevance were excluded. Articles were screened for relevance to the “Integrated Radiation Protection Framework.” Eligible studies were categorized into four domains: (1) biological basis of radiation response, (2) nutritional interventions, (3) lifestyle factors (sleep, exercise, stress), and (4) translational challenges. A qualitative synthesis was conducted to evaluate potential synergistic effects of combined modalities compared to single-agent approaches. A total of 1200 records were identified through databases and manual searches. After removal of duplicates, 950 articles underwent title and abstract screening. Of these, 350 articles were assessed in full text, and 276 studies met the predefined inclusion criteria and were included in the final qualitative synthesis. The study selection process is summarized in Figure 1. Study design for literature review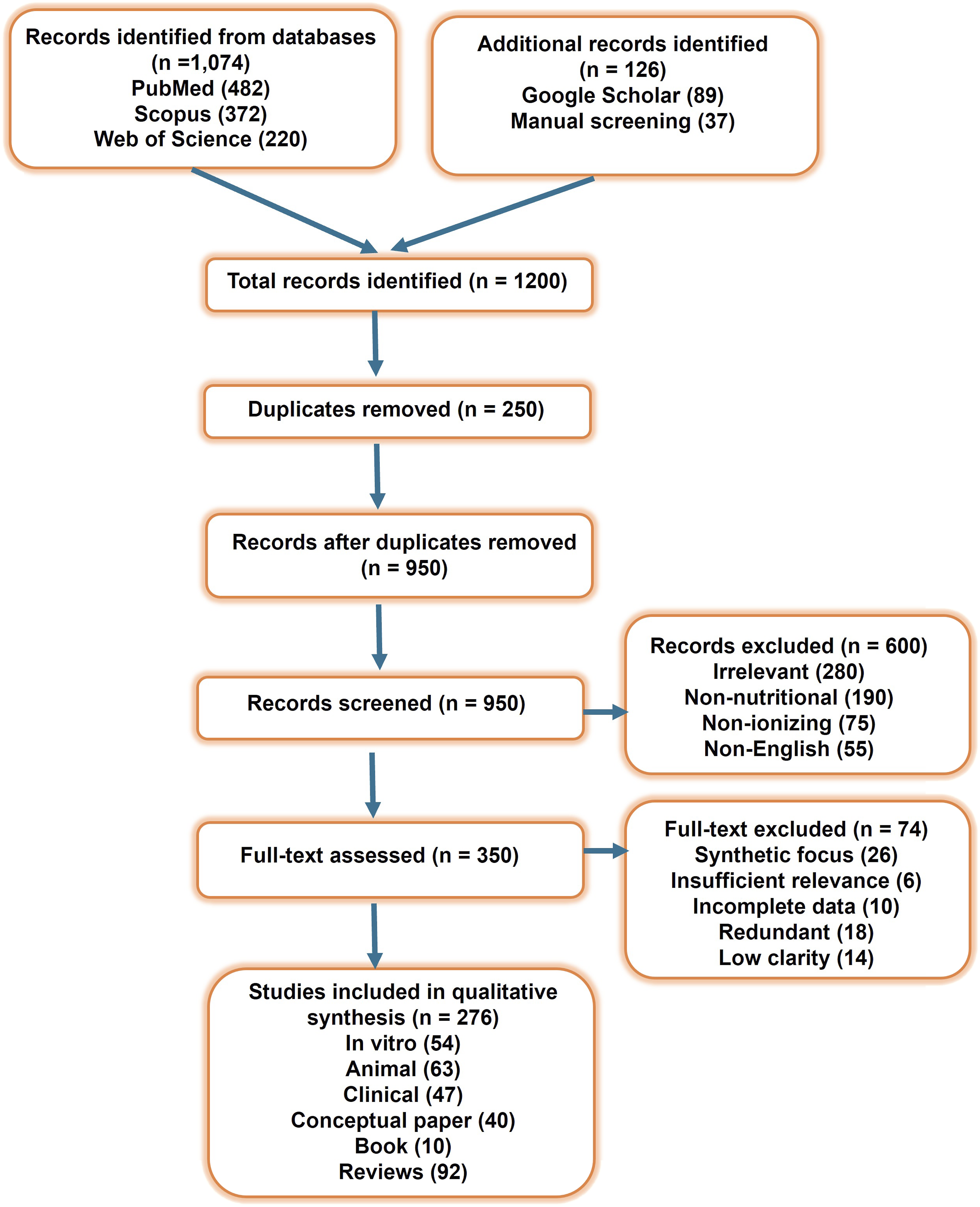
Biological Basis of Radiation Response
Mechanisms of Radiation-Induced Damage
IR, including X-rays, γ-rays, and charged particles, threatens cells, tissues, and organisms through both direct ionization of biological macromolecules and indirect damage caused by reactive oxygen species (ROS) generated from water radiolysis within cells. These processes initiate a cascade of biological events that result in acute tissue injury and long-term effects such as carcinogenesis. Radiation-induced tissue damage (RITD) arises primarily from DNA injury, oxidative stress, and microenvironmental remodeling, driven by the synergistic activation of inflammatory and damage-repair signaling pathways.6,8 IR generates a broad spectrum of DNA lesions, including single-strand breaks (SSBs), DNA double strand breaks (DSB), base modifications, DNA-protein crosslinks (DPCs), locally multiply damaged sites, and complex clustered damage, via direct or ROS-mediated interactions.21,29,30 Among these, DSBs are the most lethal and mutagenic lesions because unrepaired or misrepaired DSBs cause chromosomal translocations, deletions and mutations, leading to cell death, genomic instability, and a range of diseases including cancers, neurological, immunological, and developmental disorders.24,31 Upon DSB formation, sensor complexes such as the MRN complex rapidly detect the damage and activate the DNA damage response (DDR) through the ATM/ATR signaling pathways, these kinases phosphorylate downstream effectors including ATM-CHK2-p53 and ATR-CHK1-WEE1 axes, to orchestrate DNA repair, cell cycle arrest, or apoptosis. 32 Cells repair DSBs through several pathways: homologous recombination (HR), an error-free process active during the S/G2 phases, and nonhomologous end joining (NHEJ), a faster, but error-prone repair mechanism operating throughout the cell cycle.33,34 Repair pathway choice is regulated by end resection, cell-cycle stage, chromatin state, and DDR factor availability (e.g., ATM, DNA-dependent protein kinase catalytic subunit, BRCA1/2).35-37 Base excision repairs (BER), primarily active in the G1phase, removes oxidized or alkylated bases via sequential enzymatic steps, base excision, backbone cleavage, nucleotide replacement, and ligation, to restore genomic integrity.29,38
Beyond the initial oxidative burst, IR triggers prolonged ROS production by damaging mitochondrial DNA (mtDNA) and disrupting the respiratory chain, which increases electron leakage and mitochondrial ROS (mtROS) generation, creating a self-amplifying cycle of mitochondrial dysfunction and ongoing oxidative injury.39-42 Additionally, IR upregulates inducible nitric oxide synthase (NOS) and enhances nitric oxide (NO) levels. NO reacts with superoxide (O2•-) to form peroxynitrite (ONOO-), a potent reactive nitrogen species (RNS) responsible for nitrosative stress.43-45 IR also activates NADPH oxidases (NOX family) via downstream effectors such as Protein Kinase C (PKC) and cytokine receptors, producing secondary ROS that sustain oxidative stress and drive chronic inflammation.29,39 Figure 2 schematically depicts the mechanisms of IR–induced cellular damage, highlighting DNA injury, ROS generation, and downstream DNA damage response and inflammatory signaling pathways. Mechanism of IR-induced damage. IR (X-rays, γ-rays, and charged particles) cellular injury through both direct ionization of DNA and indirect effects mediated by radiolytically generated ROS. These processes produce SSBs, DSBs, and base modifications, collectively activating the DDR. The ensuing signaling cascades determine cell fate including accurate repair, cell-cycle arrest, senescence, apoptosis, or long-term genomic instability leading to malignant transformation. Additionally, radiation-induced ROS activate NF-κB signaling, promoting the release of pro-inflammatory cytokines such as IL-1β, TNF-α, and IL-6, which drive acute and chronic inflammation contributing to tissue injury and late radiation effects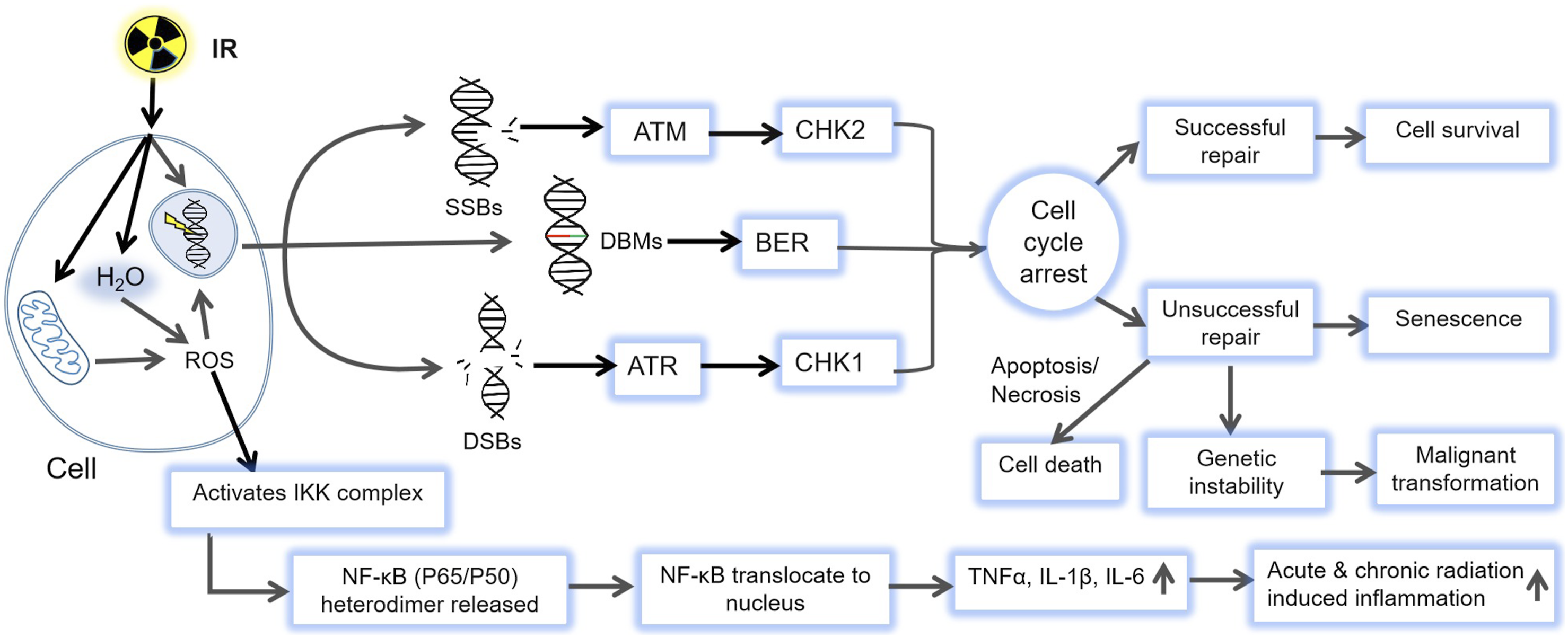
Besides, IR profoundly alters the immune landscape by triggering the release of damage-associated molecular patterns, such as high-mobility group box 1 (HMGB1), and mtDNA, which activate innate immunity via Toll-like receptors (TLRs) and the cyclic GMP-AMP synthase–stimulator of interferon genes (cGAS–STING) pathway.46-48 The release of Interleukin-1 beta (IL-1β) tumor necrosis factor-alpha (TNF-α) IL-6, transforming growth factor beta (TGF-β) further promotes vascular permeability, fibroblast activation, and fibrotic remodeling. 49 Through activation of NF-κB and MAPK, these cytokines coordinate immune and repair responses, establishing a chronic inflammatory milieu.47,50 Among T cells, regulatory T cells promote tissue repair, whereas T helper 1 (Th1) and Th17 subsets intensify chronic inflammation and fibrosis.47,51 Although IR can transiently enhance antitumor immunity by increasing antigen availability and T-cell priming, high or repeated doses often deplete T and B-cell, ultimately shifting the immune response toward suppression and increasing the risk of chronic tissue damage rather than regeneration.49,52
Inter-Individual Variability in Radiation Sensitivity: Genetics, Age, Comorbidities and Environment
Inter-individual variability represents a critical challenge in RT optimization, radiation protection, and risk assessment. This variability arises from the intricate interplay of genetic predispositions, age at exposure, pre-existing conditions, and environmental or lifestyle factors such as diet, microbiome composition, smoking, and pollution exposure. 53 These factors collectively determine the efficiency of DNA repair, antioxidant defense, immune reactivity, and overall tissue resilience to IR.
Genetic Determinants of Radiosensitivity
Genetic factors profoundly influence baseline cellular capacity for DNA damage recognition, repair fidelity, chromosomal stability, cell-cycle control, and oxidative stress regulation. Individuals with genetic syndromes such as ataxia-telangiectasia, Nijmegen breakage syndrome, and Phelan–McDermid syndromes display marked radiosensitivity, characterized by defective DNA repair and chromosomal instability.53,54 Single-nucleotide polymorphisms in TGF-β1, SOD2, XRCC3, XRCC1, and APEX have been associated with increased risk of radiation-induced fibrosis and telangiectasia following post-mastectomy RT, with combined risk alleles correlating with greater radiosensitivity, supporting the development of multi-gene predictive models for tissue responses.55,56 Genetic variants in cytokine-related genes such as TGF-β1, IL6, and TNF-α further modulates fibrotic and inflammatory responses. The TGF-β1 C-509T polymorphism, for example, has been linked to a higher incidence of radiation-induced fibrosis in breast cancer patients.54,56 Collectively, these findings underscore the potential of genomic profiling in predicting normal tissue toxicity and guiding personalized RT strategies.
Age-Dependent Radiosensitivity
Age is a dominant determinant of biological response to IR. Children exhibit increased susceptibility to long-term malignancies and developmental abnormalities, particularly in radiosensitive tissues such as the brain and thyroid. 57 Conversely, older individuals show heightened radiosensitivity due to accumulated genomic instability, mitochondrial dysfunction, immune decline, and impaired DNA repair capacity. 58 Aging diminishes the efficiency of both NHEJ and HR pathways in human fibroblasts, associated with reduced expression of XRCC4, Lig4, and Lig3, and delayed Rad51 recruitment to DSB sites. The activity of key signaling kinases such as ATM and ATR also declines with age, impairing H2AX phosphorylation and downstream DDR activation. 59 Concurrently, age-associated mitochondrial dysfunction elevates ROS generation, while diminished activities of antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx), reduce redox buffering capacity, rendering aged tissues more vulnerable to radiation-induced oxidative injury. 60 In addition, aging is accompanied by chronic low-grade inflammation (inflammaging), characterized by elevated IL-6, TNF-α, and TGF-β levels. 61
Impact of Pre-Existing Comorbidities
Comorbid conditions significantly modulate radiation responses by altering systemic metabolic and inflammatory states. Metabolic disorders such as diabetes and obesity are associated with elevated levels of IL-6, TNF-α, and TGF-β, increased baseline oxidative stress, endothelial dysfunction, and prolonged inflammation, all of which exacerbate radiation-induced tissue injury. 62 Similarly, cardiovascular and vascular diseases amplify microvascular damage, fibrosis, and ischemic injury in irradiated organs such as the heart, brain, and kidneys. 63 Patients with chronic pulmonary or cardiovascular disease exhibit reduced RT tolerance and higher risks of radiation pneumonitis and cardiac complications. 64
Sex Differences in Radiation Response
Emerging evidence indicates that sex-based biological differences also affect radiosensitivity. Studies suggest that females experience greater long-term tissue toxicity and higher risks of radiation-induced malignancies than male at equivalent doses. 65 These differences are thought to arise from hormonal modulation, immune response variability, and sex-specific gene expression patterns. Recognizing sex as a biological variable is therefore crucial in radiation risk assessment and personalized mitigation strategies.
Environmental and Lifestyle Influences
Environmental and lifestyle factors, such as dietary patterns, gut microbiome composition, stress, smoking, and exposure to pollutants, profoundly influence radiation response by modulating oxidative stress, immune regulation, and DNA repair efficiency. 66 The nutritional status determines both the baseline antioxidant defense and the capacity for tissue regeneration following radiation injury. Certain medications, particularly those influencing DNA repair or vascular function, may further modify radiosensitivity, either aggravating damage or conferring partial protection depending on their pharmacodynamics. 67 Chronic smoking and exposure to environmental pollutants elevate baseline oxidative stress and disrupt vascular and immune homeostasis, thereby intensifying radiation-induced injury. Environmental stressors often act synergistically, for instance, tobacco exposure combined with IR, or malnutrition under chronic psychological stress, to amplify DNA adduct formation, impair immune competence, and accelerate disease progression. These observations highlight the multifactorial and context-dependent nature of radiosensitivity and underscore the importance of integrative, lifestyle-based countermeasures in radiation protection. 68
Targets for Nutritional and Lifestyle Modulation
Nutritional modulation, through antioxidants, dietary micronutrients, bioactive compounds, fasting, caloric restriction, and microbiome-targeted strategies, can substantially influence the cascade of biological events triggered by IR.21,24 A growing body of research indicates that natural dietary radiomitigators represent a promising approach for enhancing antioxidant enzyme activity, protecting cells from oxidative stress, and improving RT outcomes, largely via activation of the Keap1-nuclear factor erythroid 2-related factor 2 (Nrf2) pathway. Under oxidative stress conditions, dietary compounds such as vitamin C, curcumin, and genistein activate the Keap1–Nrf2 axis by modifying Keap1 cysteines residues on Keap1, thereby preventing Nrf2 ubiquitination and degradation. Stabilized Nrf2 translocates to the nucleus, dimerizes with small Maf proteins, and induces transcription of antioxidant and detoxification genes, including SOD, CAT, GPx, and heme oxygenase-1.22,23,69 This mechanism enhances intrinsic antioxidant defenses, and reduces oxidative injury, and improves cell survival following radiation exposure. IR also activates NF-κB–dependent inflammatory signaling via ATM–NEMO interactions, ROS-driven inhibitor of kappa B kinase (IKK) activation, and DAMP/TLR pathways, leading to IκB-α degradation and nuclear translocation of NF-κB. Polyphenols, flavonoids, polyunsaturated fatty acids, and phytonutrients such as resveratrol can counteract this radiation-induced inflammation.24,66 These compounds inhibit IKKβ phosphorylation, prevent IκB-α degradation, and restrict nuclear translocation of NF-κB (P65/P50). Consequently, NF-κB remains cytoplasmic, resulting in reduced transcription of pro-inflammatory mediators such as TNF-α, IL-1β, and IL-6. This suppression limits cytokine overproduction, alleviates secondary tissue injury, and supports post-irradiation recovery. 70 Certain nutritional agents also modulate the expression of DNA damage-repair genes, enhancing the activity of enzymatic complexes involved in BER and nucleotide-excision repair (NER) pathways. 71 Moreover, functional foods and targeted supplementation including prebiotics, probiotics, and ω-3PUFAs, can reduce radiation-induced toxicity by correcting gut dysbiosis and balancing immune and inflammatory responses. 72
Beyond nutritional factors, lifestyle behaviors also play a crucial role in shaping the biological response to radiation exposure. Complementary lifestyle factors, such as regular physical activity, adequate sleep and circadian rhythm alignment, and stress management, also contribute to maintaining metabolic homeostasis, mitochondrial efficiency, and immune resilience, thereby improving the body’s capacity to tolerate and recover from radiation exposure.25,26 Collectively, these interconnected biological mechanisms highlight multiple nutrient-sensitive molecular targets, including Nrf2, NF-κB, AMP-activated protein kinase (AMPK), and DNA repair enzymes, through which nutrition and lifestyle interventions may mitigate RITD. This understanding provides a mechanistic foundation for integrative approaches that combine dietary modulation and healthy behavioral practices as complementary strategies to enhance resilience and support radioprotection.
Nutrition-Based Approaches
Energy Balance and Macronutrient Composition Modulate Radiation Sensitivity
Modifying energy intake and macronutrient composition profoundly influences the body’s response to radiation, making nutritional and metabolic regulation a promising yet underexplored avenue for personalized radioprotection and cancer therapy. Beyond total caloric intake, the quality and distribution of macronutrients including carbohydrates, proteins, and fats, play a pivotal role in modulating these biological responses. Adequate nutrient status, particularly amino acids such as glutamine, and precursors for antioxidant molecules like glutathione, supports redox homeostasis and detoxification of radiation-induced ROS. Similarly, lipid composition affects membrane stability, mitochondrial function, and inflammatory mediator synthesis, thereby influencing tissue resilience after radiation exposure. 73 Collectively, maintaining optimal energy balance and macronutrient composition not only supports DNA repair and cellular recovery but also provides a foundation for developing metabolic interventions that enhance both radioprotection and therapeutic efficacy.
Calorie Restriction and Intermittent Fasting Enhance Stress Resistance
Caloric restriction (CR) and intermittent fasting (IF) elicit profound metabolic and physiological adaptations that enhance organismal stress resistance and mitigate radiation-induced injury. CR, defined as a sustained reduction of energy intake by approximately 20–40% without malnutrition, is one of the most robust interventions known to delay aging, extend lifespan, and attenuate the onset of age-associated diseases across diverse species, including flies, rodents, and primates. 74 IF, encompassing time-restricted feeding or alternate fasting cycles, similarly promotes systemic stress resilience and has demonstrated protective effects against RITD in preclinical models. 75 Both CR and IF exert radioprotective effects by modulating key nutrient- and stress-sensing pathways, including sirtuins (SIRTs), forkhead box O transcription factors, mechanistic target of rapamycin (mTOR), AMPK, and Nrf2, all central mediators of antioxidant defense, DNA repair, and longevity regulation.74-76
CR enhances cellular and systemic resistance through multiple mechanisms: reduction of oxidative stress, improved protein homeostasis and autophagy, enhanced mitochondrial efficiency, and suppression of chronic inflammation.75,77 By decreasing proton leak and promoting mitochondrial biogenesis, CR reduces mtROS generation and protects lipids, proteins, and DNA from oxidative injury. 78 It also upregulates antioxidant and chaperone systems, such as mitochondrial Mn-SOD, cytoplasmic Cu/Zn SOD, alpha-tocopherol (AT) transfer protein, and heat shock protein 72, fortifying cells against radiation stress.75,79 Mechanistically, CR suppresses insulin-like growth factor-1 (IGF-1)/PI3K/AKT/mTOR signaling, thereby promoting DNA repair, autophagy, and metabolic flexibility while enhancing radiosensitivity in cancer cells and radioprotection in normal tissues.80,81 CR also increases p62 expression, which promotes autophagy and prevents mtROS accumulation, thus inhibiting radiation-induced liver carcinogenesis radiation-induced liver carcinogenesis. 82 Furthermore, CR attenuates pro-inflammatory cytokines such as IL-6, TNF-α, IL-8, macrophage inflammatory protein-1β, potentially reducing radiation pneumonitis and fibrosis.83,84 Studies suggest that pre-irradiation CR can prevent genotoxic initiation, while post-irradiation CR mitigates epigenetic and leukemogenic processes, jointly extending tumor-free survival. 85 However, the effects of CR in humans remain variable. Habermann et al reported no significant changes in DNA repair capacity following one year of CR (1,200–2,000 kcal/day) in postmenopausal women. 86 Overall, CR before or after irradiation improves survival and reduces cancer incidence, though its efficacy depends on the timing, duration, and magnitude of restriction. 87
IF, characterized by alternating periods of fasting and refeeding, has emerged as a promising and easily implementable approach shown in both preclinical and clinical studies to help prevent or manage aging-related diseases, cancer, neurodegenerative disorders, and metabolic dysfunctions. 75 Increasing evidence, particularly from animal models, suggests that IF enhances host resistance to radiation-induced injury by reprograming cellular metabolism and stress responses, thereby functioning as a biological mitigator of radiation damage. The radioprotective effects of IF involve several interconnected mechanisms, including metabolic reprogramming, activation of autophagy, modulation of the gut microbiome, improved DNA damage repair, attenuation of oxidative stress and inflammation, and regulation of immune responses — all of which synergistically enhance resilience to radiation exposure.77,88,89 A previous study reported that a 24-hour fast significantly reduced intestinal radiotoxicity by protecting small intestinal stem cells marked by Lgr5, Bmi1, and HopX expression, thereby improving survival in mice subjected to lethal abdominal radiation. 90 Similarly, a 3-day fasting regimen enhanced cellular stress resistance, largely through robust activation of the Nrf2 signaling pathway, a central regulator of antioxidant defense. 91 By suppressing insulin and IGF-1 activity and downregulating the mTOR pathway, IF reduces nutrient signaling, decreases the proliferation of normal cells vulnerable to radiation-induced injury, and enhances their stress resistance capacity.77,88,92 Fasting also promotes hepatic autophagy and mitophagy, which stimulate mitochondrial biogenesis and reduce the accumulation of ROS, thereby mitigating oxidative damage after irradiation. 93 During IF, nutrient sensing pathways such as SIRT1 and AMPK are activated to promote autophagy, whereas mTOR activity is inhibited. 94 Chen et al demonstrated that hepatic autophagy during fasting in mice is accompanied by activation of hypothalamic agouti-related protein neurons, linking systemic metabolic regulation with hepatic homeostasis. 95 The induction of hepatic autophagy under IF helps maintain energy balance, enhance mitochondria efficiency, preserve liver physiological and pathological integrity, sustain cellular homeostasis, suppress of tumor growth, and improve treatment sensitivity. 96 Experimental studies further support these findings. Li et al showed that short-term fasting markedly reduced radiation- and drug-induced toxicity in mice, with survival rates improving from 0% (control and 12 h fasting) to 12.5% (48 h) and 50% (72 h) after 7.5 Gy of 60Co irradiation, indicating that fasting enhances resistance to both radiation and chemical stress. 97 Likewise, Bonilla et al found that a 24-hour fast completely protected C57BL/6J mice from lethal total abdominal irradiation (11.5 Gy) by promoting intestinal stem cell regeneration and improving host survival, without diminishing tumor radiosensitivity. 98 Collectively, studies using IF have consistently reported improvements in survival rates and reduced mortality following IR exposure, particularly in rodent models, underscoring its potential as a non-pharmacological radioprotective strategy.
High-Fat, High-Sugar, and Protein-Modified Diets Modulate Radiation Sensitivity
The composition of dietary macronutrients constitutes a major modifiable determinant of metabolic and cellular responses that influence both the susceptibility to and mitigation of RITD. High-fat diets (HFDs), high-sugar diets (HSD), and diets with modified protein content, either increased or restricted, have been shown to affect radiation outcomes in preclinical and limited clinical settings.
A HFD, typically defined as providing ≥35–60% of total caloric intake from fats including both saturated and unsaturated fatty acids, has become increasingly prevalent worldwide. 99 Chronic consumptions of HFDs promotes the activation of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, which subsequently stimulate intracellular signaling pathways including MAPK, NF-κB, and JAK-STAT. 100 Activation of these cascades induces chronic low-grade inflammation and oxidative stress, leading to serine phosphorylation of insulin receptor substrates, disruption of insulin signaling, under ultimately insulin resistance. 101 Experimental studies combining HFDs with irradiation demonstrate synergistic adverse effects. For instance, C57Bl/6J mice exposed to 6 Gy of radiation and subsequently maintained on a HFD for 12 weeks exhibited exacerbated metabolic dysfunction, inflammation, gut dysbiosis, and insulin resistance. 102 Imaoka et al reported that HFD-induced obesity accelerates radiation-induced mammary tumorigenesis in rats by elevating insulin and leptin levels and increasing available energy for protein synthesis, conditions favoring rapid cancer development. 103 The combined metabolic stress of HFD and radiation exposure amplifies oxidative damage, mitochondrial dysfunction, and chronic inflammation, thereby heightening the risk of secondary malignancies and impairing the body’s capability to recover from radiation injury. 104 Interestingly, Khalil et al observed that fractionated whole-body irradiation (WBI) (0.5–2 Gy, three times weekly for two months) in HFD-fed Wistar rats induced an anti-inflammatory hepatic profile, characterized by enhanced IL-10 expression and reduced TNF-α, IL-1β, and IL-6 levels. 105 Nonetheless, most evidence indicates that HFDs increase oxidative stress, metabolic inflammation, hormonal imbalances, and impaired DNA repair, thereby intensifying radiation-induced cellular damage. 106 When combined with IR, HFDs exacerbate atherosclerosis, vascular inflammation, and lipid deposition at radiation-injured sites, worsening cardiovascular outcomes. 107 In female mice, an 8-week HFD (35% fat) significantly increased radiation-induced toxicity following whole-abdominal irradiation, 108 HFDs also potentiate genotoxicity and epigenetic reprogramming, leading to persistent insulin resistance and metabolic impairment in skeletal muscle and adipose progenitors. 109 Similarly, HFD synergizes with IR to aggravate liver injury through oxidative stress, miRNAs dysregulation, and epigenetic alterations, thereby increasing susceptibility to fibrosis, metabolic disorders, and carcinogenesis. 110 Maternal HFD exposure has been shown to heighten offspring radiosensitivity, shortening post-irradiation lifespan after total-body-irradiation (3.8 Gy at postnatal day 7), particularly in males, due to early mortality associated with bone marrow depletion and thymic lymphoma. 111 Current evidence remains insufficient to recommend HFDs for radiation protection and well-controlled studies are needed to evaluate their long-term safety and effects on normal tissue responses.
High-carbohydrate diets (HCD), particularly those high in sucrose, glucose, or fructose, have been reported to enhance radiation sensitivity by amplifying oxidative stress, promoting non-enzymatic protein glycation, activating inflammatory signaling, and inducing gut microbiota dysbiosis, all of which aggravate tissue injury following IR. 112 HCD consumption increases blood glucose and stimulates glycolytic flux, promoting excessive ROS generation via mitochondrial and enzymatic pathways. This oxidative overload depletes antioxidant defenses (e.g., GPx and SOD), disrupts redox homeostasis, and increases vulnerability to oxidative damage. Since IR itself generates ROS, the combined metabolic and radiation-induced oxidative stress can synergistically intensify cellular injury, causing more DNA strand breaks, lipid peroxidation, and genomic instability.113,114 In addition, combined HSD and HFD exposure enhances antibiotic-resistance gene expressions in the gut promoting inflammation, altering microbial composition, and facilitating gene transfer through increased bacterial permeability and ROS-mediated response, all of which are risk factors for radiation injury. 115 HCD also reduces microbial diversity and selectively enrich saccharolytic taxa, such as Bacteroides and Enterobacteriaceae. 116 Since IR disrupts gut epithelia integrity and tight junctions, concomitant HCD–induced dysbiosis permits translocation of bacteria and endotoxins into circulation, amplifying inflammation and increasing radiosensitivity in intestinal and bone marrow tissues. 117 Chronic HCD intake also leads to hyperglycemia and formation of advanced glycation end products which interact with their receptors to trigger oxidative stress and activate pro-inflammatory MAPK and NF-κB signaling, further elevating IL-6 and TNF-α levels. 118 Interestingly, D-allose suppressed tumor growth by upregulating thioredoxin-interacting protein expression, elevating ROS, and enhancing apoptosis while sparing normal tissues and reducing radiation-induced dermatitis and mucositis.119,120
Protein-modified diets also exert substantial influence on radiosensitivity in both normal and tumor tissues. Adequate and balanced protein intake is essential for radioresistance and post-irradiation recovery, as protein deficiency impairs DNA repair and tissue regeneration. Preclinical studies have provided important mechanistic insights into the role of amino acids in radiation response. Specific amino acids can modulate oxidative stress, inflammation, cellular repair processes. 121 As early as 1949, Patt et al demonstrated that L-cysteine protected rats against radiation-induced hematopoietic death and chromosomal damage, largely through its role as a glutathione (GSH) precursor mitigating free-radical–induced injury. 122 Conversely, methyl-deficient amino acid-defined diets lacking methionine and choline enhanced hepatocarcinogenesis and metastasis in diethylnitrosamine-treated rats, illustrating the tumor-promoting potential of methionine deficiency. 123 Moreover, diets containing enzymatically hydrolyzed casein enhanced intestinal regeneration after 9.0 Gy of 137Cs gamma-rays), improving DNA synthesis, crypt cell proliferation, and mucosal recovery compared with whole-casein or chow diets. 124 Similarly, whey hydrolysate peptides conferred radioprotective effects in BALB/c mice by reducing oxidative stress, enhancing immunity, and preserving intestinal and hematological function. 125 Whey protein supplementation also mitigated oxidative and inflammatory damage in irradiated mouse tongue tissue by restoring antioxidant capacity and suppressing NF-κB/p65 expression, suggesting its potential use as an adjunct in RT. 126 A high-protein diet was shown to attenuate radiation-induced immune and lipid disturbances in rats by normalizing cytokine profiles and lipid levels, including triglycerides, total cholesterol, and very low density lipoprotein cholesterol. 127
Clinical and translational studies have also begun to evaluate the impact of protein and amino acid supplementation in cancer patients undergoing RT. Arginine-enriched immunonutrition combined with ω-3PUFAs and nucleotides improved nutritional status and functional capacity in patients with head and neck cancer (HNC) or esophageal cancers undergoing chemoradiotherapy (CRT). 128 Supplementation with arginine, glutamine, and fish oil during concurrent CRT significantly reduced severe hematologic toxicity and improved treatment adherence. 129 However, Levine et al reported that individuals aged 50–65 who consumed >20% of calories from protein had a fourfold increase in cancer mortality and 75% higher overall mortality compared to those consuming <10% protein, though this risk was lower when protein was plant-based. 130 The ATHENA Project found that among breast cancer patients undergoing RT, higher carbohydrate intake was associated with increased ≥grade 2 acute skin toxicity, whereas higher total and animal protein consumption correlated inversely with this risk, suggesting that macronutrient balance may influence radiation-induced toxicity. 131
In summary, dietary macronutrient composition exerts profound influence on radiation response through modulation of oxidative stress, inflammation, energy metabolism, and DNA repair. While excessive fat or sugar intake generally exacerbates radiation injury, balanced or selectively restricted macronutrient strategies, particularly those emphasizing high-quality proteins and controlled carbohydrate intake, may enhance resilience and treatment outcomes.
Micronutrients and Bioactive Compounds Influence Oxidative and Inflammatory Pathways
Antioxidants Mitigate Oxidative Injury
Administration of antioxidants can substantially diminish radiation-induced damage in radiosensitive organs and improve overall survival. Antioxidants serve as effective radiomitigators by neutralizing radiation-generated ROS, preventing DNA strand breaks, lipid peroxidation, and mitochondrial dysfunction. They also enhance endogenous defense systems including SOD, CAT, GPx, to support cellular repair, improve tissue regeneration, and reduce post-exposure inflammation, thereby alleviating both acute and chronic radiation injury.21,24 Owing to their minimal toxicity and lack of interference with tumor control, naturally derived antioxidant compounds are regarded as promising radioprotective agents for individuals exposed to IR.22,23
Vitamins
Vitamins, obtained through diet or supplementation, have demonstrated protective effects against radiation-induced injury in both in vitro and in vivo systems. Among the thirteen known vitamins, only four, E, C, A, and D, show consistent radioprotective activity, with vitamin E being the most potent, followed by vitamins C, A, and D. 132
Vitamin E, encompassing tocopherols (α-, β-, γ-, δ-) and tocotrienols (α-, β-, γ-, δ-), acts as a potent lipid-soluble antioxidant that neutralizes radiation-induced free radicals through hydrogen atom donation. 133 AT monoglucoside protects DNA from radiation-induced SSBs. 134 Preclinical studies have demonstrated significant radioprotective effects of vitamin E and its derivatives. In animal models, vitamin E protected the protect jejunum, ileum, and colon from radiation injury, enhancing nutrient absorption via free-radical scavenging action. 135 Injectable AT (100 IU/kg) significantly prolonged survival in 60Co-irradiated mice and enhanced the radioprotective efficacy of WR-3689, increasing its dose reduction factor (DRF) from 1.35 to 1.49 when co-administered pre-irradiation. 136 Kumar et al demonstrated that subcutaneous administration of AT (400 IU/kg), 24 h pre-10.5 Gy WBI), provided superior post-irradiation survival compared to oral dosing. 137 Tocopherol succinate also conferred protection by preserving intestinal crypts, reducing apoptosis and DNA damage, and modulating antioxidant and oncogene expression.138,139
Clinical studies have shown that supplementation with AT (400 IU/d) and β-carotene (30 mg/d) mitigated adverse laryngeal effects in HNC patients 3 years after RT. 140 Clinically, combined oral supplements of vitamin E and pentoxifylline reduced the severity and duration of RT-induced oral mucositis and dysphagia in HNC patients, likely through antioxidant, antifibrotic, and microcirculatory mechanisms. 141
Vitamin C (VC) (ascorbic acid) functions as a potent water-soluble antioxidant that scavenges free radicals, thereby protecting DNA strand breaks and oxidative injury.24,27 Preclinical evidence shows that orally VC administered before or shortly after 1 Gy WBI reduced micronuclei and chromosomal aberrations in mouse bone marrow. 142 Pretreatment with VC (150mg/kg/day) improved 42% survival and alleviated intestinal injury, following 14 Gy WBI with bone marrow transplantation, whereas post-treatment offered limited benefit. 143 Oral 6-palmitoyl ascorbic acid-2-glucoside (80 mg/kg, 1 h pre-irradiation) significantly mitigated γ-radiation–induced genomic and oxidative damage in spleen, bone marrow, and blood, improving overall survival. 144 In clinical settings, concurrent supplementation of vitamin E (100 IU) and VC (500 mg) twice daily during RT helped reduce xerostomia and protect salivary glands, although without affecting overall survival in HNC patient. 145
Intraperitoneal administrations of Vitamin D (200 ng/kg/day) demonstrated anti-inflammatory, antioxidant, and radioprotective effects in rat lacrimal glands, supported by histopathological and cytokine analyses. 146 Similarly, vitamin A was shown to prevent gamma-radiation-induced intestinal injury by preserving brush border enzyme activity and reducing oxidative stress. 147
Carotenoids
Carotenoids act as powerful antioxidants due to their conjugated double bonds, which efficiently scavenge peroxyl, alkoxyl, hydroxyl, and superoxide radicals.23,69 Di Mascio et al identified lycopene (LY), abundant in red fruits and vegetables, as the most efficient carotenoid for quenching singlet oxygen and peroxyl radicals. 148 Pretreatment with LY significantly reduced gamma-radiation–induced oxidative stress and DNA damage in rat hepatocytes, confirming its radioprotective potential. 149 In vitro, LY protected human blood lymphocytes from X-irradiation (0.5-2 Gy) when administered immediately after exposure, though it did not enhance post-irradiation repair. 150 β-carotene and astaxanthin also preserved membrane lipid integrity under 60Co γ-irradiation (30 kGy). 151 Dietary intakes rich in VC, E, β-carotene, β-cryptoxanthin, and lutein–zeaxanthin has been correlated with reduced cumulative DNA damage in airline pilots chronically exposed to IR. 152 Histopathological studies confirmed LY’s protective role in reducing acute gastrointestinal radiation injury 153 and oral mucositis 154 in animal models, underscoring its potential to alleviate radiotherapy-induced complications.
Trace Elements Contribute to DNA Repair and Antioxidant Defense
Trace elements, even at trace concentrations (µg/g), play essential roles in mitigating IR–induced cellular injury by detoxifying ROS and facilitating DNA repair.
155
Their radioprotective effects are primarily mediated through antioxidant, anti-inflammatory, and DNA-stabilizing mechanisms.
156
Some clinical and experimental studies have demonstrated that supplementation with trace elements can serve as an effective adjunct radioprotection strategy during RT. The nutritional strategies in mitigating radiation-induced damages are illustrated in Figure 3. Nutritional strategies in mitigating radiation-induced damages. Dietary interventions help alleviate radiation-induced oxidative stress, DNA damage, inflammation, and gut dysbiosis while enhancing immune function, DNA repair, antioxidant defense, and cell homeostasis. CR, calorie restriction; IF, intermittent fasting; SOD, superoxide dismutase; CAT, catalase; GPx, glutathione peroxidase; ω-3 PUFA, omega-3 polyunsaturated fatty acid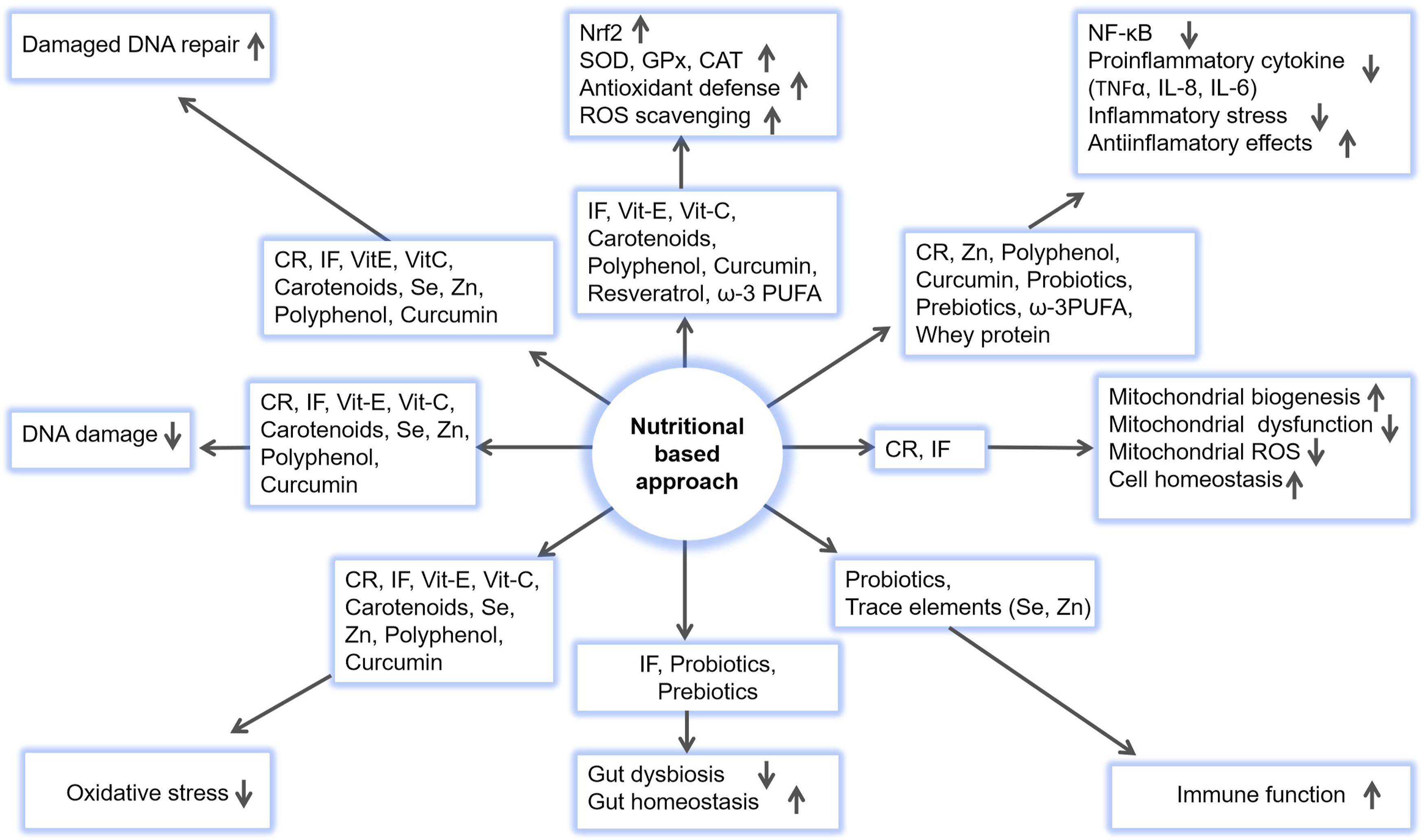
Selenium
Selenium (Se) is an essential trace element whose compounds and metabolites, such as selenomethionine (SeM), sodium selenite (SSe), and Ebselen, have long been recognized for their radioprotective potential.155,157,158 Se functions as a cofactor for numerous selenoproteins and enzymes, including thioredoxin reductase and GPx, which collectively reduce oxidative stress and inflammation while safeguarding genomic integrity. 159 Mechanistically, SeM promotes p53-mediated BER by enhancing APE1 activity and modulating Gadd45a–protein interactions, thus accelerating the resolution of oxidative DNA lesions following radiation exposure. 160 Experimental studies show that intraperitoneal injections of SeM (4mg/kg) or SSe (0.8 mg/kg) in mice significantly improved 30-day survival when administered before (1 or 24 h) or shortly after (within 15 min) 9 Gy exposure, with minimal toxicity. 161 Similarly, the selenocystine derivative 3,3′-Diselenodipropionic acid conferred radioprotection against WBI in Swiss albino mice by suppressing hepatic lipid peroxidation, protein carbonylation, splenic apoptosis, while preserving gastrointestinal and hematopoietic integrity. 162 Selenium-L-methionine also protected rat lung tissues from IR-induced pneumonitis and fibrosis by modulating immune and redox signaling. 163
Clinically, oral Se supplementation alleviated RT-induced diarrhea, ageusia and dysphagia in HNC patients without compromising tumor control. 164 In a Phase 1 clinical trial involving 15 metastatic cancer patients, oral SSe (5.5–49.5 mg) administered 2h before each RT fraction was well tolerated, with most patients experiencing disease stabilization and significant pain relief during palliative RT. 165 However, given Se’s narrow therapeutic window, dose optimization is crucial to balance efficacy and safety in clinical use.
Zinc
Zinc (Zn) is another essential trace element that serves as a cofactor for more than 300 enzymes including Cu/Zn-SOD, a key antioxidant enzyme.155,156 Zn protects cells by stabilizing sulfhydryl-containing proteins and antagonizing redox-active metals such as iron and copper, thereby limiting hydroxyl radical formation and mitigating site-specific oxidative injury following radiation exposure.156,166 Notably, Zn-bound metallothionein provided greater protection against 30 Gy γ-radiation-induced DNA damage than other thiol-containing molecules such as glutathione or albumin. 167 Zn deficiency disrupts the structure and binding efficiency of Zn-dependent transcription factors such as p53, an essential DDR regulator, thereby impairing DNA-repair processes. 168 Moreover, Zn depletion weakens both BER and NER pathways by destabilizing DNA repair proteins. 169
Preclinical studies have demonstrated radioprotective effects of Zn across several experimental models. Zinc sulfate preserved thyroid functions by maintaining T3, T4, TSH balance under 131I exposure. 170 In mice, co-administration of Zn and silymarin significantly alleviated 2 Gy γ-radiation–induced testicular injury by improving sperm quality, testosterone levels, and testicular cell density while suppressing oxidative stress, apoptosis, and inflammation. 171
Clinically, oral zinc sulfate (50 mg, three times daily) markedly reduced RT-induced mucositis and oral pain in HNC patients without adverse effects. 172 Similarly, zinc L-carnosine supplementation alleviated oral mucositis, xerostomia, and dysgeusia, enhancing quality of life in HNC patients. 173 In breast cancer patients undergoing RT, oral Zn sulfate reduced radiation-induced dermatitis in a dose-dependent manner by promoting DNA repair, maintaining epithelial barrier integrity, and reducing oxidative stress. 174 Recently, Li X et al developed a zinc-releasing oral patch that effectively prevented radiation-induced ulcerative mucositis in HNC patients by activating the KLF5/FoxO signaling cascade, enhancing collagen synthesis, and suppressing oxidative stress in irradiated tissues. 175 Consistent with these findings, Zn supplementation broadly attenuates radiation-induced oral mucositis and epithelial damage by modulating oxidative and inflammatory pathways while supporting mucosal regeneration and wound-healing processes. 176
Phytochemicals Exert Protective Effects
Phytochemicals, including polyphenols (e.g., resveratrol, quercetin, and epigallocatechin gallate), curcuminoids (curcumin), isothiocyanates (sulforaphane), carotenoids (LY), terpenoids, and various alkaloids, have gained increasing attention as potential radioprotective and radiomitigative agents.24,177 These bioactive compounds attenuate IR-induced damage through multiple mechanisms, including scavenging of radiation-generated ROS/RNS, activation of endogenous antioxidant defenses via Nrf2 signaling, modulation of DDR pathways such as ATM/ATR and Poly (ADP-ribose) polymerase (PARP), stabilization of mitochondrial function, suppression of pro-inflammatory cytokine signaling (e.g., NF-κB), and regulation of apoptotic cell-death cascades.24,74,177,178
Polyphenols
Natural polyphenols, plant-derived secondary metabolites structurally characterized by the presence of two or more phenolic rings, exhibit consistent radioprotective efficacy through their potent antioxidant, free-radical-scavenging, anti-inflammatory, and DNA-damage-modulating properties.24,71,179 A single subcutaneous dose of the isoflavone genistein (GEN) (25–400 mg/kg) markedly improved 30-day survival in CD2F1 male mice when administered 24 h, but not 1 h, before a lethal 9.5 Gy γ-irradiation. 180 In CT26 tumor-bearing mice, GEN pretreatment (200 mg/kg, 1 day before irradiation) significantly reduced intestinal apoptosis after 5 Gy abdominal irradiation, ameliorated crypt survival and villus shortening, and delayed tumor growth. 181 Recent reviews highlight GEN’s ability to enhance tumor radiosensitivity via modulation of oxidative stress, DNA repair, cell-cycle control, and apoptosis across several cancer models, including breast, prostate, and lung cancers. 182 Similarly, oral naringenin (50 mg/kg for 7 days) protected normal tissues against radiation-induced oxidative, genotoxic, and apoptotic damage by enhancing endogenous antioxidant defenses, increasing spleen colony formation, and regulating p53, Bax, and Bcl-2 expression while inhibiting NF-κB-mediated pro-apoptotic signaling. 183 A polyphenol-rich extract blend (100 mg/kg for 7 days before WBI) preserved hematological and biochemical parameters, reduced TNF-α and oxidative stress, normalized stress- and apoptosis-related gene expression, and maintained liver and kidney morphology, demonstrating strong radioprotective efficacy. 184
In a phase II clinical study, daily administration of the green-tea polyphenol epigallocatechin-3-gallate (EGCG) (400 mg) significantly reduced radiation-induced intestinal epithelial injury in cervical and endometrial cancer patients, none developed grade 3 or higher toxicity, supporting its promise as a safe and effective radioprotective agent. 185 EGCG supplementation also decreased pain associated with acute radiation-induced esophagitis during thoracic RT for lung cancer. 186
Curcumin
Curcumin (CUR), a phenolic compound derived from Curcuma longa rhizomes, demonstrates potent antioxidant and radioprotective properties.21,24 In a rat skin irradiation model, oral CUR given at 150 mg/kg administered one day before and for three days after irradiation) significantly mitigated oxidative damage by lowering malondialdehyde (MDA) levels and enhancing the activities of SOD, CAT, and GPX. 187 In human lymphocytes, CUR pretreatment (0.5–100 μg/mL) prior to γ-irradiation (0.05–2 Gy) decreased γ-H2AX/53BP1 foci formation and chromosomal translocations in a concentration-dependent manner, indicating strong in vitro radioprotective effects. 188 Nano formulation of CUR enhances its bioavailability and radioprotective efficacy by further reducing MDA levels, increasing SOD activity, preventing aberrant DNA methylation, suppressing TNF-α, IL-6, IL-1β, and TGF-β1, and promoting differential effects in normal versus tumor tissues—protecting normal cells while sensitizing cancer cells to radiation through ROS accumulation, NF-κB inhibition, and activation of pro-apoptotic signaling (p53, p21, BAX). 189 In p53-mutant prostate cancer cells, CUR functions as a radiosensitizer by blocking TNF-α/NF-κB–mediated Bcl-2 upregulation, shifting the Bcl-2:Bax ratio, and activating cytochrome c–dependent caspases, thereby potentiating radiation-induced apoptosis. 190
A meta- analysis confirmed that CUR supplementation significantly reduced radiation-induced dermatitis in breast cancer patients, 191 and systemic reviews suggest that CUR alleviates CTx- and RT-induced oral mucositis by attenuating oxidative and inflammatory stress. 192
Resveratrol
Resveratrol (RSV), a stilbene-type polyphenol abundant in grapes, peanuts and red grapes, and mulberries, exhibits potent antioxidant, anti-inflammatory, and radioprotective actions.23,24 RSV protects hematopoietic stem cell from WBI by suppressing NOX 4-mediated ROS production and activating SIRT1-dependent antioxidant pathways (SOD2, GPx). 193 Pretreatment with RSV (10 mg/kg for 30 days) mitigated X-ray–induced oxidative injury by lowering MDA and enhancing total antioxidant capacity, SOD, and CAT activities. 194 Post-irradiation administration of RSV-loaded polymeric nanocapsules markedly reduced hematopoietic and intestinal injury in mice, preserved mucosal architecture, improved hematologic indices, and increased survival, suggesting efficacy as a radiomitigator. 195 RSV (25 mg/kg daily for two weeks before and three days after irradiation) also mitigated radiation-induced premature ovarian failure by preserving follicular reserve and suppressing PARP-1/NF-κB–driven inflammation via upregulation of SIRT1 and peroxisome proliferator-activated receptor γ. 196 RSV (0.1–1 mM/ml), administered immediately or 1hour post-irradiation, reduced radiation-induced DNA damage in human lymphocytes in a dose- and timing-dependent manner. 197 Combined RSV and alpha-lipoic acid therapy further attenuated radiation-induced pneumonitis and pulmonary fibrosis in mice, reducing alveolar inflammation, hemorrhage, and vascular damage. 198 Additionally, RSV protected against radiation-induced cutaneous injury by activating the AMPK/SIRT7/HMGB1 pathway, thereby enhancing DNA repair in keratinocytes. 199
Functional Foods and Nutritional Supplements as Translational Opportunities
Clinical Evaluation of Nutritional Dietary Intervention in Radiation Mitigation
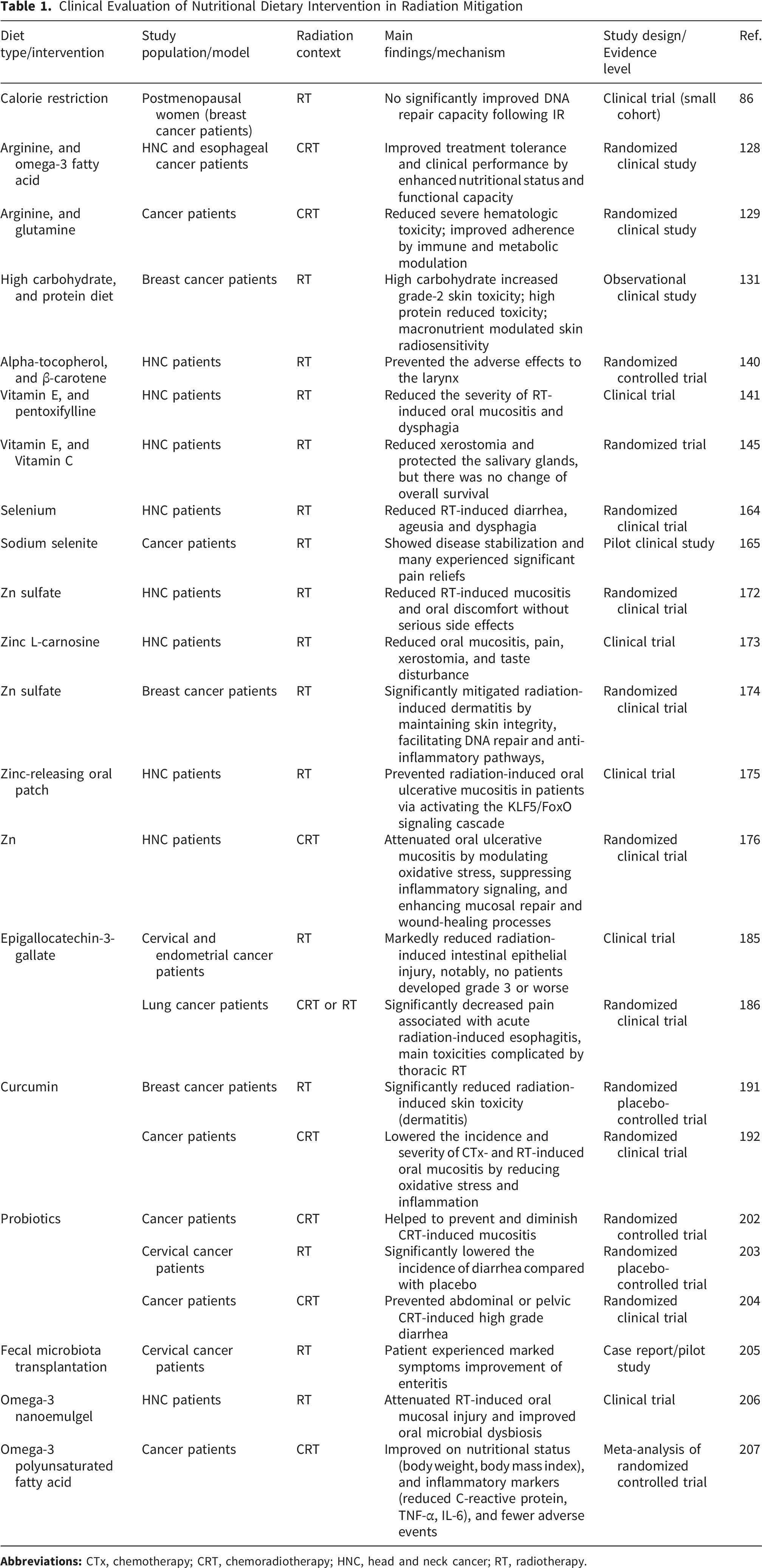
Gut Microbiota and Probiotics Shape Host Radiation Response
The gut microbiota, comprising trillions of bacteria, archaea, fungi, and viruses inhabiting the gastrointestinal (GI) tract, plays a crucial role in modulating the host’s systemic response to IR. RT and chemoradiation frequently induce gut microbiome dysbiosis, characterized by a disrupted balance between beneficial and pathogenic microbes. This dysbiosis activates innate immune receptors, particularly TLRs, stimulating NF-κB–mediated signaling and driving the release of pro-inflammatory cytokines like IL-1β, IL-6, and TNF-α.208,209 The gut microbiota thus emerges as a key determinant of radiation sensitivity by regulating both mucosal and systemic immune responses, the so-called microbiota–immune axis.200,210 Dysbiosis-induced radiation enteritis is exacerbated by overgrowth of pathogenic taxa such as Enterobacteriaceae, Bacteroides, and Citrobacter rodentium, which promote NF-κB–driven inflammation. Conversely, beneficial species, including Faecalibacterium prausnitzii, Bifidobacterium infantis, Akkermansia muciniphila, and Lactobacillus spp., produce metabolites that strengthen epithelial barriers, suppress inflammation, and facilitate mucosal repair.201,211 Therapeutic modulations of the gut microbiome, via probiotics, prebiotics, microbial metabolites, or fecal microbiota transplantation (FMT), has demonstrated significant mitigation of radiation injury in preclinical and clinical models.209,212
Probiotics, defined as live microorganisms conferring health benefits to the host, protect against IR injury by modulating microflora composition and immune responses. Lactobacillus plantarum and L. rhamnosus GG enhance intestinal stem-cell DNA repair and suppress DNA damage and inflammation through FXR-FGF15 and cGAS/STING pathways. 211 Oral administration of Akkermansia muciniphila sattenuates radiation-induced GI damage by improving colon length, crypt architecture, and goblet cell numbers while reducing IL-6, TNF-α, and IL-1β expression. 213 Engineered second-generation probiotics expressing interleukin-22 (Lactobacillus reuteri or Escherichia coli) significantly improved 30-day survival in irradiated mice by stimulating Lgr5 + intestinal stem-cell generation. 214
Clinically, probiotic combination containing Saccharomyces boulardii, Bifidobacterium, and Lactobacillus acidophilus have been proposed to prevent CRT-induced mucositis. 202 In a double-blind, placebo-controlled, trial, supplementation with L. acidophilus LA-5 and Bifidobacterium animalis subsp. lactis BB-12 (1.75 × 109 CFU) significantly reduced RT-induced diarrhea in cervical cancer patients. 203 Similarly, probiotic administration in patients undergoing abdominal or pelvic CRT markedly decreased the incidence of high-grade diarrhea compared with placebo. 204
Prebiotics, non-digestible food ingredients such as dietary fibers, selectively promote beneficial bacterial growth and modulate gut composition. Their metabolites, such as short-chain fatty acids (SCFAs), help restore microbial balance and reduce inflammation during RT. SCFAs derived from Ruminococcus spp. activate Wnt signaling via α-linolenic acid, promoting intestinal stem-cell proliferation and epithelial repair after mucositis. 215 Marine-derived prebiotics, such as alginate oligosaccharide, attenuate dysbiosis and oxidative stress by enhancing SCFA and tryptophan metabolite production. 216 Similarly, konjac glucomannan mitigates WBI-induced intestinal and hematopoietic damage by increasing microbiota diversity, SCFA production, and gut-barrier integrity. 217
Microbial-derived ω-3PUFAs inhibit NF-κB activation through TLR4/IKK signaling, attenuating inflammation and oxidative stress. 218 Gut microbial metabolites, such as propionate and tryptophan derivatives, reduce DNA damage markers (p53, 53BP1) and ROS in intestinal and bone marrow stem cells, thus protecting against hematopoietic and gastrointestinal injury. 219 FMT has also demonstrated mitigation of radiation-induced pneumonitis through microbiome restoration via the gut–lung axis. 220 In the first reported clinical case, FMT successfully alleviated radiation enteritis in a 59-year-old cervical cancer patient following stool transplantation from her healthy son donor. 205
Omega-3 Fatty Acids Modulate Inflammatory Signaling
ω-3 PUFAs, primarily eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), are derived from marine sources such as cod, halibut, mackerel, salmon, and menhaden. 221 EPA and DHA, the principal ω-3 PUFAs, suppress the synthesis of pro-inflammatory eicosanoids, prostaglandin E2 (PGE2) and leukotriene B4 (LTB4), while simultaneously serving as precursors for anti-inflammatory lipid mediators. This dual mechanism attenuates NF-κB activation and decreases the expression of inflammatory enzymes and cytokines such as cyclooxygenase-2, TNF-α, and IL-1β.222,223 Specialized pro-resolving mediators, derived from ω-3 PUFAs limit neutrophil infiltration, promote macrophage-mediated efferocytosis, and facilitate tissue regeneration, thus restoring homeostasis following IR injury. 224 Preclinical findings indicate that an ω-3-rich diet reduces systemic inflammation and preserved bone microarchitecture and mineral density in irradiated mice, primarily through TNFα downregulation. 225 Clinically, a randomized controlled trial (NCT05214495) demonstrated that topical ω-3 nanoemulgel effectively alleviated RT-induced oral mucosal and improved oral microbial balance in patients with head and neck cancer, providing translational evidence of ω-3 therapy. 206 ω-3 PUFAs-enriched oral nutritional supplements during CRT improved patients’ nutritional status (body weight, body mass index), and inflammatory markers (reduced C-reactive protein, TNF-α, IL-6), and fewer adverse events, although overall clinical evidence remains limited. 207
Lifestyle Interventions in Radiation Mitigation
RT remains a cornerstone of cancer treatment; however, it also inflicts collateral damage on healthy tissues, underscoring the importance of lifestyle-based interventions to mitigate radiation-induced injury.25,26,226 Recent studies highlight that, alongside dietary interventions, physical activity, stress management and sleep quality can ameliorate damage by modulating oxidative stress and inflammation.226,227 Considering the growing number of cancer survivors, particularly those treated for breast and prostate cancers, there is an urgent need to explore non-pharmacological strategies to minimize radiation toxicity and improve patient outcomes. Beyond improving physical performance, exercise and other lifestyle factors have been shown to regulate molecular and cellular processes that control ROS generation, DNA repair, and immune function.227,228 Thus, incorporating lifestyle modification, encompassing physical activity, psychological resilience, and circadian regulation—into survivorship and clinical care frameworks is crucial to target biochemical and systemic pathways associated with radiation injury.
Physical Activity Enhances Resilience Through Redox Regulation, DNA Repair, and Immune Modulation
Physical activity (PA) protects against ROS-induced DNA damage and decelerates cancer progression by elevating antioxidant enzymes such as CAT, SOD (particularly SOD2), and GPX. 229 Preclinical evidence shows that pre-exercise training significantly increased SOD and CAT activities while lowering pro-inflammatory cytokines (IL-1, IL-6, TNF-α, TGF-β) in irradiated mice. 230 Exercise interventions also reduce oxidative DNA damage markers such as 8-hydroxy-2′-deoxyguanosine, enhancing genomic stability via DNA repair pathways. 231 Furthermore, exercise also reduces γH2AX foci formation and micro-nucleated reticulocytes post-radiation exposure, indicating enhanced DNA repair capacity and reduced apoptosis. 230 Moderate exercise supports hematopoietic stem cell survival after irradiation via the carnosine/Slc15a2-p53 axis, alleviating myelosuppression. 232 Exercise further influences epigenetic regulation. Modulating DNA methylation in promoters of oxidative stress-related genes such as SOD2 and L3MBTL histone methyl-lysine binding protein 1 (L3MBTL1), thereby enhancing antioxidant expression and anti-tumor activity. 233 In cancer models, chronic exercise upregulates Ccl5 and Cxcl10 chemokines, promoting CD8+ T-cell infiltration and inhibiting tumor progression. 234 By lowering leptin and IGF-1 levels, exercise suppresses PI3K/Akt/mTOR signaling and activates AMPK, favoring pro-apoptotic tumor pathways.235,236
Clinical studies also support the beneficial effects of PA in cancer patients. In metastatic breast cancer patients, a six-month PA program enhanced GPx activity, reduced ROS production, and lowered NOX and myeloperoxidase (MPO) activity, indicating decreased oxidative stress. 237 Reductions in MDA, a lipid peroxidation marker, have been observed following exercise in cancer patients, reflecting improved membrane integrity. 238 Exercise also alters methylation of tet methylcytosine dioxygenase 1 (TET1), and DNA methyltransferase 3α and 3β, which regulate DNA methylation dynamics and tumor progression in breast cancer patients. 239 PA also modulates immune regulation through pattern-recognition receptors such as TLRs, which fine-tune inflammatory signaling via pathways such as NF-κB pathways. 240 Exercise-induced myokines, including IL-6 and interleukin 15 (IL-15), mediate immune resilience; IL-15 promotes proliferation of CD8+ T, CD19+ B, and natural killer (NK) cells, while IL-6 enhances tumor suppression via DNA damage and immune activation.241,242 PA strengthens NK cell–mediated antitumor activity by upregulating CD57, NKG2D, CXCR3, and TIM-3 surface markers and by stimulating IL-15–driven NK maturation. 243 Acute dynamic exercises transiently mobilize circulating leukocytes, especially NK and γδ T cells, followed by CD8+ and CD4+ T cells. 244 In addition to epigenetic changes, physical exercise affects the level of blood growth factors and hormones. For example, ovarian cancer patients have reported a reduced IGF-1 and leptin after physical activity, both hormones believed to be associated with tumor growth, suggesting the production of anti-proliferative signals. To the contrary, higher vascular endothelial growth factor level is indicative of improved oxygenation, though it should be explored to get the meaning in clinical sense. 245 Together, these findings underscore exercise as a systemic radioprotective agent acting through antioxidant, immune, and metabolic reprogramming.
Stress Management and Psychosocial Factors
Chronic psychological stress triggers sustained activation of the hypothalamic–pituitary–adrenal axis, resulting in elevated cortisol levels that suppresses immune activity, impairs DDR, and disrupt cell-survival signaling, thereby increasing radiosensitivity. 246 Persistent stress also promotes ROS accumulation and antioxidant depletion, exacerbating tissue damage and compromising recovery following irradiation. 247 These mechanisms suggest that prolonged psychological stress may aggravate radiation-induced cellular injury through immune dysregulation, oxidative imbalance, and impaired DNA repair processes.
In humans, higher perceived stress and radiation-related anxiety correlate strongly with increased fatigue, poorer self-reported health, and diminished quality of life following radiation exposure or RT. The mental health impacts observed after major nuclear accidents, such as Chernobyl and Fukushima illustrate how psychological effects can surpass physical injury in magnitude and duration.248,249 In these populations, anxiety and depression levels remained elevated regardless of the actual radiation dose received, leading to the conceptualization of radiation anxiety, a term referring to persistent fear of radiation-induced health risks.250,251 Moreover, public health countermeasures intended to reduce exposure—such as forced evacuations or restrictive policies—sometimes inadvertently heightened psychological distress and deteriorated quality of life. 251 The long-term mental health consequences may persist for decades; for instance, elderly residents in unexposed areas near Nagasaki continued to experience elevated anxiety about radiation risks many years after the atomic bombing. 252 The emotional burden of nuclear disasters is further compounded by social stigma, discrimination, and the profound community disruption caused by displacement. 253 Therefore, coordinated, community-based psychological interventions are crucial for reducing stress-induced immune suppression and enhancing resilience among radiation-exposed populations. Structured psychological support, public education, and transparent communication strategies can minimize anxiety and foster long-term adaptation. These psychosocial frameworks are as essential as biomedical countermeasures in mitigating the total health burden of radiation exposure. 254
Potential Roles of Mindfulness, Yoga, and Stress-Reduction Programs
Psychological and behavioral interventions, such as mindfulness, yoga, and structured stress-reduction programs, offer measurable benefits for individuals undergoing RT. These approaches not only alleviate distress and fatigue but also influence physiological pathways involved in oxidative stress, immune modulation, and DNA repair. 251 Mindfulness is defined as the intentional cultivation of non-judgmental awareness and focused attention to present-moment experiences.255,256 Initially developed as a clinical tool for anxiety and depression, mindfulness-based interventions are now applied widely across oncology to enhance emotional regulation and stress resilience. 257
Randomized and controlled trials (RCTs) demonstrate that mindfulness-based stress reduction effectively reduces anxiety and depressive symptoms while improving coping ability and emotional control among breast cancer patient undergoing RT. 258 Yoga, a mind–body practice integrating physical postures, breathing, and meditation, has also shown radioprotective and restorative effects. In a clinical study, participants practicing yoga demonstrated 14.5% less DNA damage compared with a 28.8% increase in the control group, suggesting a tangible protective effect against radiation-induced genotoxic stress. Furthermore, patients perceived stress scores dropped significantly, from a mean of 20.4 to 14.9 after RT, while anxiety scores were half, from 8.5 to 4.1, compared to negligible improvements in the control group. 259 These results support yoga’s dual psychological and physiological benefits during RT. A meta-analysis of RCTs confirmed that yoga, mindfulness, and psychosocial interventions effectively reduce cancer-related fatigue and improve sleep quality and overall functioning in patients under RT. 260 Sincere RT is associated with common side effects such as fatigue, skin toxicity, insomnia, depression, and anxiety, stress-reduction strategies are essential components of comprehensive cancer care.261-263 For example, structured stress-management programs for women with breast cancer undergoing RT were found to significantly decrease anxiety and depression, with some trials also reporting improvements in physical functioning and quality of life. 262 Educational interventions that provide information and emotional support before RT sessions, such as the radiation therapy prepare program, effectively reduce psychological distress, alleviate treatment-related fear, and enhance treatment adherence and readiness. 263 Collectively, these findings highlight that psychological well-being and emotional regulation are fundamental determinants of radiation resilience. Integrating mind–body therapies into radiation treatment protocols represents a cost-effective and evidence-based approach to complement molecular and pharmacologic countermeasures.
Circadian Rhythms and Sleep
Radiation Response and Circadian Biology
The circadian rhythm, a cell-autonomous 24-hour biological timing system, governs a wide array of physiological processes including metabolism, immune regulation, oxidative balance, and DNA repair. 264 When synchronized with environmental cues such as light and feeding schedules, it maintains cellular homeostasis and orchestrates metabolic and immune functions. Conversely, circadian disruption caused by shift work, trans-meridian travel, or irregular sleep patterns dysregulates immune-cell phenotype and cytokine production, leading to heightened inflammation and disease susceptibility. 264 At the molecular level, core circadian genes including circadian locomotor output cycles kaput (CLOCK), brain and muscle ARNT-like 1 (BMAL1), cryptochrome (CRY1-2), and period (PER1-3), control the timing of cell-cycle checkpoints (such as Wee1, p21, p16 and p53) and directly interact with DDR pathways such as NER, HR, and NHEJ.265-267 The CLOCK–BMAL1 complex regulates transcription of key DDR genes including ATM and BRCA1/2, thereby modulating DNA repair efficiency and influencing radiosensitivity. Circadian gating also governs antioxidant defenses via BMAL1-driven accumulation of Nrf2 protein, which regulates downstream antioxidant enzymes such as SOD, CAT, and GPx. 266 As a result, cells exhibit higher oxidative tolerance and more efficient ROS detoxification at specific circadian phases. In addition, melatonin, a hormone secreted in a circadian pattern, contributes further by regulating gut microbiota composition and attenuating inflammation through BMAL1-related mechanisms. 268
Clinical Evaluation of Physical Intervention in Radiation Mitigation
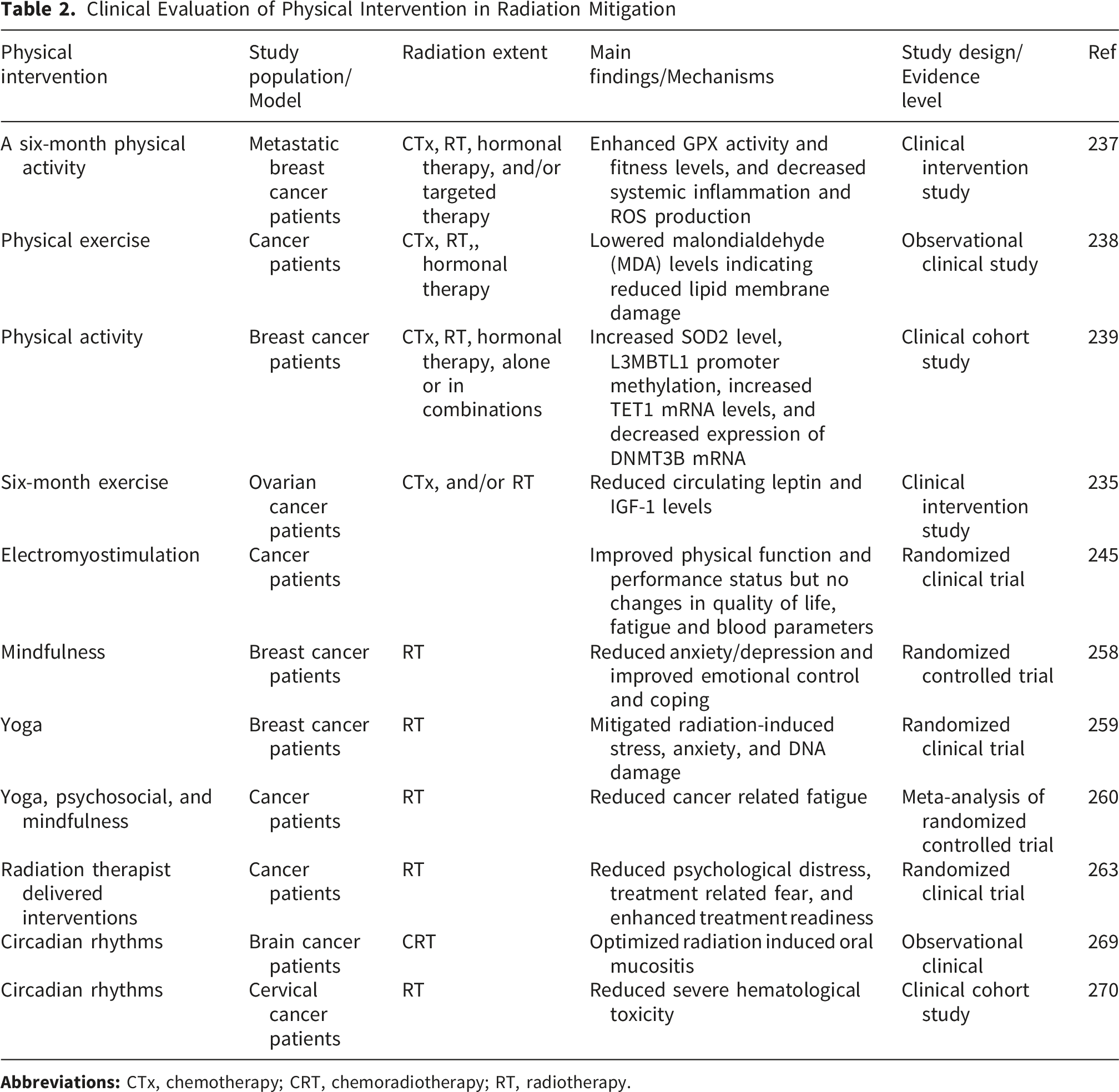
The Impact of Sleep Quality on DNA Repair and Immune Function
Sleep is a fundamental biological process that reinforces immune competence, promotes cellular repair, and maintains redox balance. During slow-wave sleep, cytokines such as IL-1 and TNF are upregulated, initiating immune responses and facilitating tissue recovery following irradiation. 271 Adequate sleep also enhances T-cell proliferation and cytotoxic activity, helping limit bystander damage and clear apoptotic or damaged cells. The neuroendocrine shifts that occur during sleep, characterized by reduced sympathetic activity and increased parasympathetic tone, lead to decreased cortisol and other immunosuppressive hormones. 272 Simultaneously, chromatin mobility and the activity of DDR proteins such as Rad52 and Ku80 increase, facilitating DNA lesion recognition and repair. 273 Sleep also supports melatonin-driven antioxidant and mitochondrial protection, mitigating oxidative and lipid peroxidation damage to DNA and cell membranes. 274
Conversely, sleep deprivation or poor-quality sleep impairs cytokine production, antibody formation, and NK cell function, reducing host defense against radiation-induced injury. 264 Inadequate sleep may compromise immune surveillance and delay recovery following radiation exposure. Hence, maintaining consistent, restorative sleep cycles contributes directly to radiation resilience by harmonizing immune, oxidative, and genomic repair mechanisms.
Summary of Lifestyle Interventions
Lifestyle factors, including physical activity, stress regulation, mindfulness, yoga, circadian alignment, and restorative sleep, work synergistically to reinforce endogenous defense systems against radiation-induced injury. These approaches mitigate oxidative stress, support DNA repair, restore immune balance, and optimize metabolic efficiency. Importantly, their benefits extend beyond cancer therapy to occupational, environmental, and space radiation exposure contexts. The concise overview of lifestyle interventions in radiation mitigation is presented in Figure 4. Lifestyle interventions in radiation mitigation. Lifestyle practices such as physical activity, stress management, and adequate sleep contribute to radiation recovery by enhancing DNA repair, strengthening antioxidant defense, modulating inflammatory responses, and promoting tumor cell death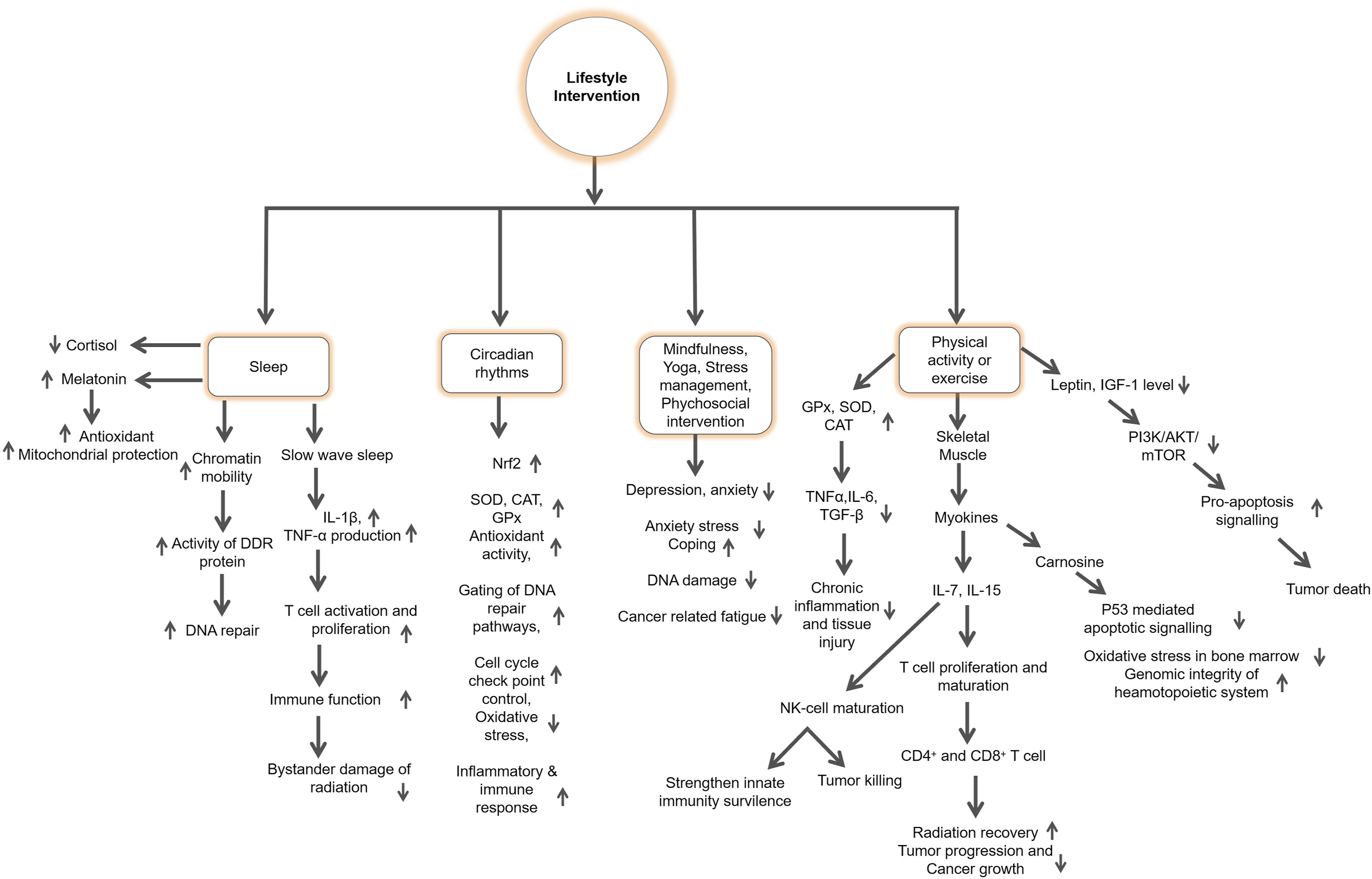
Integrating these lifestyle strategies into standard radioprotection frameworks promotes long-term physiological homeostasis and resilience. When combined with nutritional interventions, they provide a comprehensive, systems-level defense that enhances recovery, limits secondary damage, and supports overall health outcomes after radiation exposure.
Although emerging evidence indicates that lifestyle factors such as physical activity, adequate sleep, and stress management may contribute to improved resilience against radiation-induced damage, the current evidence base remains relatively limited. Much of the available data comes from preclinical studies or indirect clinical observations. Therefore, additional well-designed mechanistic studies and large-scale clinical trials are needed to better clarify their effectiveness and to develop stronger evidence-based recommendations.
Integrative Approaches
Synergistic Nutrition–Lifestyle Frameworks for Multi-Pathway Protection Against Radiation-Induced Injury
IR perturbs multiple interconnected biological networks, inducing oxidative stress, chronic activation of pro-inflammatory cascades, mitochondrial dysfunction, and immune-metabolic imbalance, which together drive tissue injury and long-term health risks. To comprehensively mitigate these effects, relying on single modality, such as nutritional or lifestyle intervention alone, may be insufficient. Instead, an integrative framework that combines dietary and lifestyle strategies offers a more effective and systems-level approach for protecting against RITD. Antioxidants and bioactive polyphenols such as RSV, EGCG, CUR, ω-3PUFA have shown potent radioprotective effects at the molecular level. These compounds act through upregulation of Nrf2-driven antioxidant gene expressions, suppression of NF-κB-mediated inflammatory signaling, and enhancement of DNA repair pathways, as supported by multi-omics analyses (metabolomics and transcriptomics).22-24,28,69,155-157,177-179,200,201 In parallel, systems biology evidence demonstrates that lifestyle factors including regular physical activity, sufficient sleep, and synchronized circadian rhythms, enhance mitochondrial biogenesis, immune resilience, and metabolic homeostasis.25,26,226-228,230,246,251,264-266,271-273 Although nutrition and lifestyle interventions act on overlapping biological pathways, each also targets distinct molecular mechanisms, therefore, neither can completely substitute for the other. When combined, their effects are often synergistic offering a more comprehensive defense by buffering oxidative stress, improving metabolic adaptability, and attenuating systemic inflammation (Figure 5).27,28 Emerging evidence suggests that integrative strategies, such as nutrition plus exercise, may attenuate RT-induced side effects by modulating inflammatory responses, preserving muscle metabolism, and improving treatment-related symptoms, though further RCTs are to confirm mechanistic insights.
26
Integrative frameworks of nutrition and lifestyle interventions in mitigating radiation-induced tissue damage. Nutritional and lifestyle interventions act synergistically to alleviate IR-induced injury by modulating overlapping yet distinct biological pathways. Nutritional approaches, including antioxidants, polyphenols, omega-3 fatty acids, and functional foods, enhance redox balance, DNA repair, and anti-inflammatory defenses, while lifestyle strategies such as physical activity, adequate sleep, stress reduction, and circadian alignment reinforce mitochondrial function, immune resilience, and metabolic homeostasis. Together, these complementary interventions provide a holistic and multi-pathway defense system that enhances recovery and reduces long-term radiation-related toxicity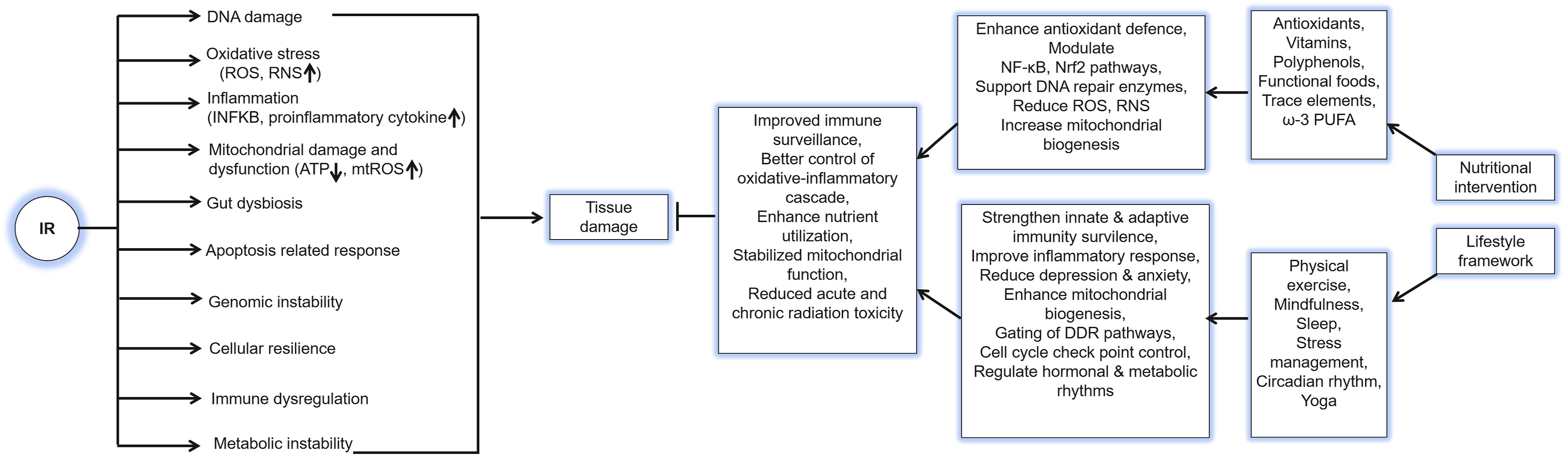
Moreover, physical activity, sleep quality, and a balanced gut microbiome (supported by prebiotics, probiotics, and IF) enhance nutritional absorption and utilization. Concurrently, dietary polyphenols, CR, IF, adequate sleep and regular exercise jointly promote mitochondrial biogenesis, increase ATP production, and counteract radiation-induced mitochondrial damage. Circadian rhythms, exercise, IF, vitamins, polyphenols such as RSV and CUR collectively strengthen antioxidant defense systems. By integrating these complementary approaches, the body achieves a more resilient physiological state, improving recovery from acute damage, minimizing long-term consequences, and maintaining systemic homeostasis. Together, these interventions enhance innate and adaptive immune surveillance, resistance to post-irradiation infections, and overall host resilience.
Importantly, aligning dietary timing and sleep
Challenges, Limitation, and Research Gaps
Although nutritional interventions and lifestyle modifications have demonstrated promising health benefits in in preclinical studies, substantial challenges remain in translating these findings to humans. High experimental variability limited clinical data, and the absence of standardized methodologies make it difficult to define optimal conditions or elucidate the precise mechanisms through which diet and lifestyle synergistically promote radiation recovery. Most current evidence is derived from rodent models, which differ considerably from humans in metabolic rate, microbiome composition, and radiation sensitivity. RCTs validating mechanistic outcomes in humans, such as redox modulation, DNA repair, and immune restoration, are largely lacking. Clinical trials face additional obstacles, including limited sample size, logistical and ethical constraints, and difficulty in rigorously controlling lifestyle variables. Maintaining consistent adherence to prescribed diets, sleep hygiene, and exercise regimens in real-world settings also poses practical challenges. While preclinical studies strongly support the radioprotective effects of lifestyle interventions, human data remains sparse and often inconclusive. For instance, one clinical trial reported that long-term CR, aerobic exercise, or their combination did not significantly alter DNA strand break repair capacity in overweight and obese postmenopausal women. 86 The use of antioxidants during RT presents another important challenge. Although antioxidant supplementation may alleviate radiation-associated adverse effects, concurrent use during RT could potentially reduce therapeutic efficacy, increase recurrence risk, and shorten survival. Thus, the safety and timing of antioxidant intake in cancer patients undergoing RT remain unclear and warrant further high-quality clinical investigation. 275 Interindividual variability adds another layer of complexity. Genetic polymorphisms, baseline inflammatory status, gut microbiome composition, and mitochondrial function can all significantly modify redox and inflammatory responses to dietary or exercise interventions. Personalized or stratified approaches remain underdeveloped. Moreover, many cancer patients receive concurrent chemotherapies, corticosteroids, or targeted agents that can alter nutrient metabolism and interfere with oxidative and inflammatory pathways, complicating clinical implementation. Practical challenges also exist. Fatigue, nausea, appetite loss, and psychological stress commonly experienced by RT patients make adherence to structured dietary or physical activity regimens difficult. The interplay among lifestyle components is often nonlinear, for example, excessive high-intensity exercise may elevate oxidative burden and counteract the benefits of dietary antioxidants, highlighting the need of integrated and well-balanced intervention strategies. Currently, there are no established consensus guidelines regarding the optimal dose, timing, or composition of dietary interventions, nor standardized clinical thresholds for lifestyle parameters such as exercise intensity, sleep duration, or circadian alignment. Future research should prioritize mechanistic, longitudinal, and personalized studies to establish evidence-based recommendations and ensure the safety and efficacy of combined diet–lifestyle interventions in radiation mitigation.
Conclusions and Future Perspective
This review underscores the integrative role of nutrient-rich diets and lifestyle interventions in mitigating RITD and promoting systemic recovery. Current evidence indicates that no single intervention can fully counteract the multifaceted biological disruptions caused by IR, which simultaneously perturbs redox homeostasis, DNA integrity, immune function, and metabolic regulation. Therefore, combining complementary dietary and lifestyle strategies provides a more comprehensive, physiologically coherent, and sustainable approach to radioprotection. Micronutrients, bioactive compounds, and functional foods including vitamins, minerals, polyphenols, and ω-3PUFA, stabilize redox balance, attenuate inflammatory signaling through NF-κB suppression, and stimulate DNA repair via Nrf2 and p53 pathways. Concurrently, lifestyle factors such as regular physical activity, sufficient sleep, circadian synchronization, and stress reduction promote mitochondrial biogenesis, enhance immune surveillance, optimize metabolic efficiency, and improve nutrient utilization. Together, these interconnected systems reinforce cellular defense mechanisms and resilience to radiation injury. Collectively, these interventions cultivate a more robust internal environment that mitigates radiation-induced oxidative and inflammatory injury, protects normal tissue from collateral damage, and minimizes secondary complications such as metabolic imbalance and immune suppression. Such strategies hold promises not only in clinical RT but also in occupational, environmental, and space radiation contexts, where chronic low-dose exposure poses long-term risks. Importantly, nutritional and lifestyle interventions should not be regarded as alternative but complementary modalities that act through distinct yet interlinked molecular networks. Their combination provides additive or synergistic across multiple pathways—enhancing redox stability, genomic maintenance, and immune recovery beyond what can be achieved by any single approach. From a translational standpoint, the integration of nutrition and lifestyle strategies with pharmacological radioprotectors may represent a future paradigm for holistic radiation countermeasures. Preliminary studies and systematic reviews support their potential to alleviate radiation-induced side-effects and improve patient well-being during and after RT. Nevertheless, mechanistic validation through rigorously controlled clinical trials remains limited. Future research should focus on elucidating the molecular mechanisms that underlie the synergistic effects of combined dietary and lifestyle interventions through advanced multi-omics and systems biology approaches. It is equally essential to determine the safe and effective doses, timing, and compositions of bioactive compounds and dietary patterns that yield optimal radioprotective effects. Establishing standardized experimental and clinical protocols, along with validated biomarkers, will enable a more accurate assessment of radioprotective efficacy and inter-individual variability. Moreover, developing personalized frameworks that integrate genetic, metabolic, and gut microbiome profiles could optimize adaptive responses to radiation exposure, ultimately leading to tailored prevention and recovery strategies. Ultimately, advancing this integrative model requires close collaboration between radiation biologists, nutrition scientists, clinicians, and behavioral researchers. By bridging mechanistic insight with clinical application, evidence-based dietary and lifestyle interventions may emerge as vital components of radiation mitigation, enhancing treatment tolerance, reducing long-term toxicity, and improving the quality of life for both patients and radiation-exposed populations.
Footnotes
Acknowledgments
The authors gratefully acknowledge Dr. Yi Shang for her careful reading and insightful comments on this manuscript, and Ms. Hiromi Arai for her outstanding technical assistance and invaluable administrative support. Polash Chandra Karmakar and Tusher- Al-Arafat were fellows of the Nuclear Researchers Exchange Program 2025 and 2023, respectively, supported by the Ministry of Education, Culture, Sports, Science, and Technology (MEXT) and the Nuclear Safety Research Association of Japan.
Author Contributions
Conceptualization, P.C.K., T.A.A., and B.W.; methodology, P.C.K., T.A.A. and B.W.; data curation, P.C.K. and T.A.A.; writing—original draft preparation, P.C.K. and T.A.A.; writing—review and editing, P.C.K., T.A.A., C.L., T.K. and B.W.; visualization, P.C.K. and B.W.; supervision, B.W.; funding acquisition, C.L., T.K. and B.W. All authors have read and approved the final version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially supported by the Japan Society for the Promotion of Science (JSPS) KAKENHI, Grant-in-Aid for Scientific Research (C), Grant Numbers JP22K12387, JP23K11431 and JP25K15450.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
All data discussed in this review were obtained from previously published studies; no new data were generated.