
Abstract
Background
In the present study, the therapeutic effects of Toosendanin (TSN) against Mycoplasma pneumoniae (MP)-induced pneumonia (MPP) in mice.
Research Design
Swiss albino mice were exposed to MP culture for 2 days, causing pneumonia, and then treated with TSN for 3 days. Lung weights, total protein, IgM, C-reactive protein, oxidative stress, and inflammatory cytokines were assessed. Histological alterations were evaluated in lung tissues. Molecular docking analysis was performed to test TSN’s interaction with inflammatory cytokines, including interleukin-1 beta (IL-1β), interleukin-6 (IL-6), transforming growth factor-beta 1 (TGF-β1), and nuclear factor kappa B (NF-κB) were evaluated targets.
Results
TSN treatment significantly reduced lung weight by approximately 25% compared to the MP-infected group (P < 0.05). Total protein and C-reactive protein (CRP) levels decreased by 30% and 40%, respectively. Malondialdehyde (MDA), was reduced by 35%, while antioxidant enzyme levels (e.g., SOD, CAT) increased by 20%–25%. Pro-inflammatory cytokines such as IL-1β and IL-6 were significantly lowered by 40%–50%. Histological analysis revealed a marked reduction in inflammatory cell infiltration and alveolar damage scores (P < 0.01). Molecular docking confirmed strong binding interactions between IL-1β, IL-6, TGF-β1, NF-κB and TSN.
Conclusions
The present findings confirm the beneficial effects of TSN in protecting mice from pneumonia.

Introduction
Pneumonia is a widespread contagious disease that involves the infection of the lung tissue by several pathogens. 1 There is a current consensus that the incidence and death rates associated with pneumonia are increasing worldwide. Pneumonia is the primary cause of mortality in children globally. Each year, this disease is responsible for around 1.3 million child fatalities worldwide. 2 Mycoplasma pneumoniae (MP) is a prokaryotic microbe without a cell wall and is a major cause of infections in both the lower and upper respiratory tracts. 3 Mycoplasma pneumoniae (MPP) is a condition characterized by inflammation of the respiratory and pulmonary systems caused by an infection with MP. It mainly impacts children aged 3 to 15, particularly those in school-aged categories. This disease is distinguished by its increased prevalence and vulnerability to recurrence. 4 Because of the swift expansion and progress at this stage, the immunity of children is not yet completely developed, making them highly susceptible to MP infection. Epidemiological research suggests that MPP exhibits periodicity, with outbreaks occurring at intervals of 3-7 years.
MP is the predominant pathogen causing community-acquired pneumonia (CAP) in children in specific areas, accounting for about 40% of cases. The hospitalization rate for affected children is approximately 18%, highlighting its importance as a significant world health issue in pediatric healthcare.5,6 CAP is linked to significant morbidity and mortality rates, and it poses a significant risk to public health globally. MP was responsible for causing 8-40% of cases of CAP in children who were hospitalized.7,8 MP can be spread and disseminated among individuals by sneezing and coughing as airborne droplets.9,10 MP infections can lead to several extrapulmonary symptoms, including meningoencephalitis, myocarditis, and mucocutaneous eruptions. 11 Furthermore, MP causes mucocutaneous complications. Those disorders often coincide with systemic inflammation and an increased likelihood of long-term complications. 12
While MP infection typically resolves on its own and does not necessitate antibiotic intervention, individuals of all age ranges have the potential to develop severe, life-threatening, or extrapulmonary conditions. Macrolide antibiotics are commonly used in current clinical practice to treat MPP in children. Tetracycline and fluoroquinolone antibiotics have demonstrated efficacy in eradicating MP infections. However, it is worth noting that tetracyclines can lead to discoloration of teeth and bones in children. 13 Fluoroquinolones can also impact the musculoskeletal system, including the muscles, joints, and tendons. Historically, macrolides have been the primary medication for treating MP infections because of their lower side effects. The failure of antibiotic treatment has led to a rise in the death rate in recent years. 14 The extensive utilization of macrolide medications has resulted in the progressive emergence of MP resistance, posing challenges in the realm of antibiotic therapy. There has been a worldwide surge in reports of MP resistance, with a particular focus on the Asia-Pacific region, in recent years. 15
TSN is a tetracyclic triterpene that is obtained from the bark and fruits of the Melia toosendan plant. The previous studies already highlighted that TSN has therapeutic potentials such as anti-inflammatory, antioxidant, ant-parasitic, and anticancer properties.16,17 Toosendanin (TSN), has shown potential anti-inflammatory and antimicrobial properties in respiratory diseases, including Mycoplasma pneumoniae pneumonia (MPP). TSN has been shown to alleviate acute lung injury by modulating inflammatory pathways, such as NF-κB and Nrf2/HO-1, which are implicated in MPP pathogenesis.18,19 It also exhibited broad-spectrum antimicrobial activity against respiratory pathogens like Staphylococcus aureus and Klebsiella pneumoniae, suggesting potential utility in mixed or secondary infections common in MPP.20,21 These findings support further investigation into TSN as a candidate for managing inflammation and microbial burden in MPP.
However, there are no reports to claim the therapeutic properties of the TSN against the MPP. Therefore, the present work was carried out to understand the therapeutic properties of TSN against the MP-induced MPP condition in mice.
Materials and Methods
Material and Methods
The drugs and chemicals obtained from Sigma Chemicals, USA, include TSN, etc. The estimation of biochemical markers was conducted using test kits supplied by Abcam, USA, and Elabscience, USA, respectively.
Experimental Animals
The present investigation utilized Swiss albino mice with an average body weight of 24 ± 6 g. The mice were housed in sterile polypropylene confines and sustained in organized settings at a temperature of 23 ± 2°C, with air humidity ranging from 50% to 60%, and a 12-hour light/dark cycle. Every mouse in the study was provided with unlimited access to pellet food and sterile water during the experimental periods. Before commencing the tests, a period of seven days was allotted for the mice to adapt to the laboratory environment.
Animal Grouping and Treatment Protocol
After a 1-week acclimatization period, mice were randomly divided into three experimental groups (n = 6 per group):
Group I (Control): Mice received no treatment and served as the healthy control group.
Group II (MP group): Mice were intranasally administered 100 µL of Mycoplasma pneumoniae (MP) culture daily for 2 consecutive days to induce pneumonia.
Group III (MP + TSN group): Mice received 100 µL of MP culture intranasally for 2 days, followed by intraperitoneal administration of Toosendanin (TSN) at a dose of 20 mg/kg once daily for 3 consecutive days.
Following the completion of the treatment period, all mice were euthanized via cervical dislocation under light anesthesia. Blood and lung tissue samples were collected immediately for further analysis. Lung wet and dry weights were measured to evaluate pulmonary edema.
Analysis of C-Reactive Protein (CRP) and Ig-M Levels
The concentrations of CRP and IgM in the serum of experimental mice were determined using the respective test kits. The assays were conducted in triplicate as per the instructions provided by the manufacturer of the kits (Abcam, USA).
Analysis of Oxidative Stress Marker Levels
Lung tissues harvested from the mice were homogenized in an ice-cold buffer and centrifuged at 15 000 rpm for 20 minutes at 4°C. The resulting supernatant was collected and analyzed for oxidative stress biomarkers, including malondialdehyde (MDA), superoxide dismutase (SOD), and glutathione (GSH). Quantification was performed using commercially available colorimetric assay kits (Elabscience, USA), following the manufacturer’s standardized protocols.
Analysis of Total Protein Content and Total Cell Count in the Bronchoalveolar Lavage Fluid (BALF)
Saline aliquots of 30 mL were injected into the right middle lobe of the experimental mice to collect the BALF samples. The BALF was thereafter centrifuged at 6000 rpm for 5 min. The resulting BALF sample was subsequently transferred to new vials for additional examinations. A hemocytometer was used to enumerate the total cell quantities in the collected BALF sample under a microscope. The total protein concentration was measured using the assay kit (Abcam, USA).
Analysis of Inflammatory Cytokine Levels
Bronchoalveolar lavage fluid (BALF) was collected from each mouse and used to measure the concentrations of inflammatory cytokines, including IL-1, IL-6, IL-8, NF-κB, and TGF-β1. Cytokine levels were determined using enzyme-linked immunosorbent assay (ELISA) kits provided by Elabscience (USA), according to the manufacturer’s instructions. All assays were commercially standardized to ensure consistency and reproducibility.
Histopathological Analysis
Histological analysis was performed on lung tissue samples to identify histological changes. In summary, the lungs were treated with 10% neutral formalin and subsequently paraffinized. The paraffin-embedded tissues were subsequently cut into sections of 5 µm in diameter. The tissues were stained using eosin and hematoxylin stains and then examined under a microscope to observe the histological changes in the lung tissues.
Molecular Docking Analyses
The target assignment, dataset retrieval, and validation: The targets for the molecular docking analysis, including IL-1β (PDB ID: 8I1B), IL-6 (UniProt ID: P08505), TGF-β1 (UniProt ID: Q3UNK5), and NF-kB (PDB ID: 1NFK), were selected. IL-1β and NF-kB were retrieved from the Protein Data Bank (PDB). At the same time, IL-6 (Model Confidence Score (MCS): 80.88%) and TGF-β1 (MCS = 76.56%) were obtained from the AlphaFold Protein Structure Database (https://alphafold.ebi.ac.uk/; Accessed on: 27 June 2024) with high structure modeled confidence. The structures were then rigorously validated using the Ramachandran Plot (RM), available in the MolProbity database (https://molprobity.biochem.duke.edu/; accessed on 25 June 2024), to ensure the accuracy of our methods.
Preparation of Protein Structures and Ligands
The protein structures of IL-1β (PDB ID: 8I1B) and NF-kB (PDB ID: 1NFK) retrieved from the PDB were processed using Discovery Studio Visualizer (DSV) v19.1.0.18287 (https://www.accelerys.com/), which involved the removal of water molecules and their associated ligands. The structure of TSN, downloaded from the PubChem compound database in SDF format with provided 3D atomic coordinates, 22 was then converted into PDB format using DSV.
Molecular Docking and Visualization
The molecular docking process was initiated for the defined targets, such as IL-1β, IL-6, TGF-β1, and NF-kB, with TSN by implementing Autodock Vina, which offers an integrated scoring algorithm and is integrated into PyRx software. 23
The Autodock vina uses a new scoring mechanism,

C- Sum of intermolecular and intramolecular distance; ∑ - Over all of the pairs of atoms; ftitj - Symmetric set of interaction functions; rij - Interatomic distance defined targets and ligand were converted into PDBQT format. The PyRx virtual screening tool identified the interacted active site residues of the targeted biomarkers and the grid box properties were set as for IL-1β (size_x = 58.46 Å; size_y = 53.70 Å; size_z = 55.99 Å), IL-6 (size_x = 61.83 Å; size_y = 56.23 Å; size_z = 54.84 Å), TGF-β1 (size_x = 25 Å; size_y = 25 Å; size_z = 25 Å), and NF-kB (size_x = 25 Å; size_y = 25 Å; size_z = 25 Å). After molecular docking, the interactions were analyzed for the targets with the TSN in 3D and 2D by implementing Discovery Studio Visualizer v19.1.0.1828 (Dassault Systèmes BIOVIA, Rue Marcel Dassault, Vélizy-Villacoublay-78140, France (https://www.accelerys.com/; Accessed on 26 June 2024).
Statistical Analysis
The findings are statistically analyzed using SPSS software V20, and the data are given as the mean ± SD of three measurements. The data are analyzed using a one-way ANOVA and Tukey’s post hoc assay to measure the significance level. A significance level of P < .05 is used to determine statistical significance.
Results
Effect of TSN on the Lung Wet/Dry Weight Ratio in the Experimental Mice
Figure 1 shows that there was a notable increase in the lung wet/dry weight levels in the mice with MPP than the control. Notably, TSN at the 20 mg/kg concentration treatment successfully decreased these changes in the MPP mice. These findings revealed that TSN remarkably diminished the pulmonary edema and water content in the lung tissues of the MPP mice. Effect of TSN on the Lung Wet/Dry Weight Ratio in the Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group
Effect of TSN on the CRP and Ig-M Levels in the Serum of Experimental Mice
The concentrations of CRP and IgM in the serum of the experimental mice were assessed, and the results are presented in Figure 2. The mice that were induced with MPP exhibited a significant increase in both CRP and Ig-M levels when compared with the control. Nevertheless, the administration of 20 mg/kg of TSN successfully decreased these increases in the mice with MPP. Effect of TSN on the CRP and Ig-M Levels in the Serum of Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group
Effect of TSN on the Total Protein and Total Cell Counts in the BALF
The total protein concentrations and cell counts in the BALF of the control and treated mice were assessed, and the results are presented in Figure 3. An increase in both the total protein concentrations and total cell counts was seen in the mice with MPP as compared with the control. The TSN (20 mg/kg) treatment led to a remarkable decrease in both the total protein concentrations and total cell counts in the MPP mice. Effect of TSN on the Total Protein and Total Cell Counts in the BALF of the Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group
Effect of TSN on the Oxidative Stress Marker Levels in the Lung Tissues of the Experimental Mice
Figure 4 displays the concentrations of oxidative stress marker levels, including MDA, GSH, and SOD, in the lung homogenates of the experimental mice. A notable elevation in the level of MDA and a diminution in antioxidant GSH and SOD levels were seen in the BALF of mice with MPP, as compared with control. Notably, the treatment with 20 mg/kg of TSN remarkably diminished the level of MDA and elevated both SOD and GSH levels in the lungs of the MPP mice. These findings proved the antioxidant capacity of the TSN. Effect of TSN on the Oxidative Stress Marker Levels in the Lung Tissues of the Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group
Effect of TSN on the Inflammatory Marker Levels in the BALF of the Experimental Mice
Figure 5 displays the inflammatory cytokine levels in the BALF of experimental mice. A significant increase in the IL-1, IL-6, IL-8, and TGF-β1 levels was seen in the BALF of the MPP mice in comparison with the control. Notably, the treatment of 20 mg/kg of TSN considerably diminished these cytokine levels, including IL-1, IL-6, IL-8, and TGF-β1, in the BALF of mice with MPP. These results proved that TSN showed remarkable anti-inflammatory activity. Effect of TSN on the Inflammatory Marker Levels in the BALF of the Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group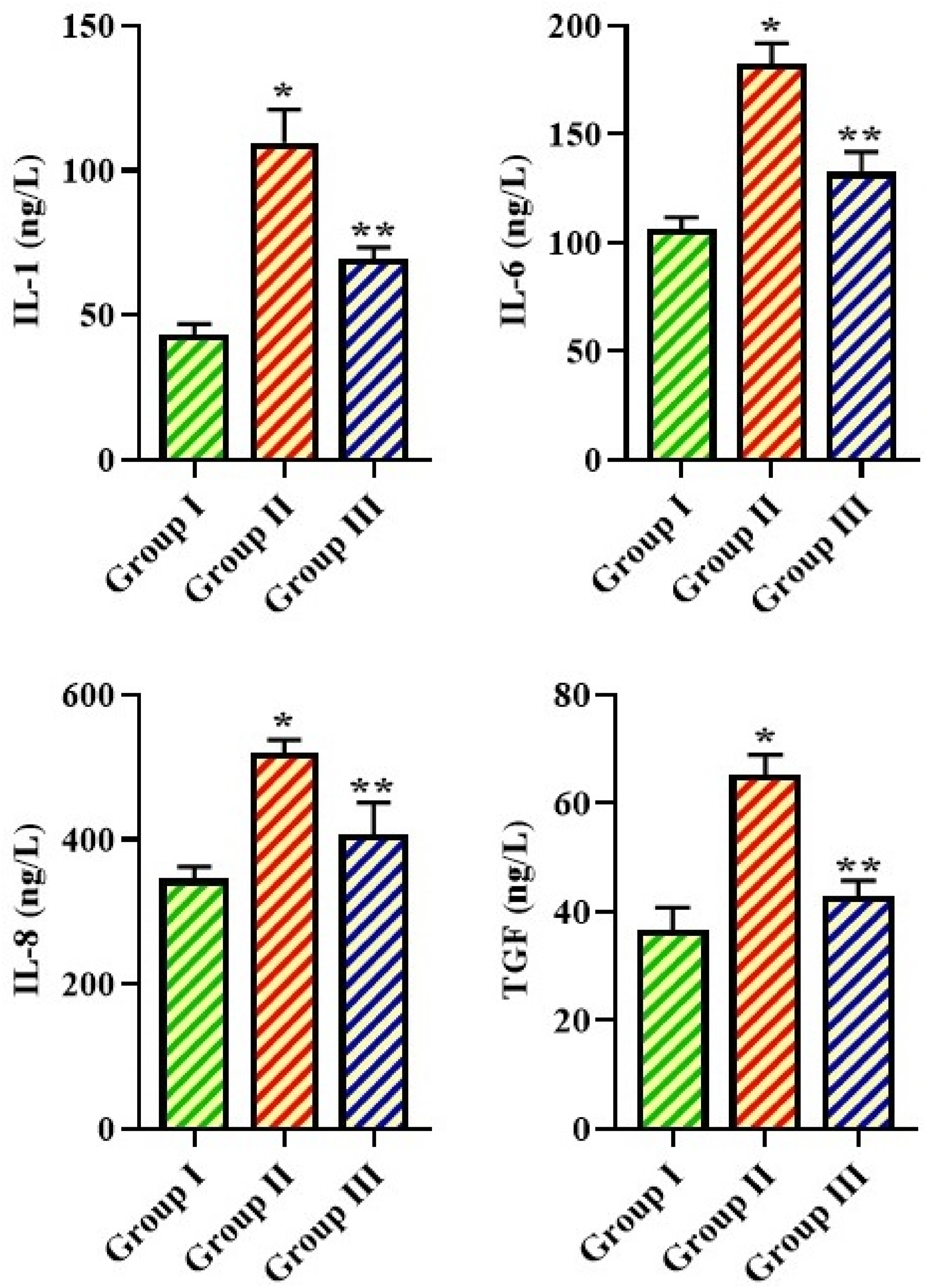
Effect of TSN on the Lung Tissue Homogenates
The NF-κB levels in the lung tissues of both control and MPP-induced mice were investigated using assay kits, and the outcomes are presented in Figure 6. A considerable increase in NF-κB levels was seen in the lungs of the MPP mice compared with the control. In contrast, the administration of 20 mg/kg of TSN remarkably diminished the NF-κB levels in the MPP mice, which further supports the anti-inflammatory properties of the TSN. Effect of TSN on the Lung Tissue Homogenates of the Experimental Mice. Each bar Displays the Mean ± SD of (n = 6) Assays. Statistical Studies are Conducted Using SPSS Software by One-Way ANOVA and Tukey’s Post Hoc Test. The Symbol '*' Indicates Statistical Significance at P < 0.01 Compared to the Control Group, whereas '**' Indicates Statistical Significance at P < 0.05 Compared to the Pneumonia-Induced Group
Effect of TSN on Lung Histopathology
The results of the histopathological analysis of the lungs of both control and treated mice are given in Figure 7. The control mice exhibited normal patterns of alveolar cells without any indications of inflammation or tissue damage in the lungs. In contrast, mice with MPP had significant histological alterations, including alveolar wall thickening, constriction of bronchial tubes, and increased inflammatory cell infiltrations. Remarkably, the administration of TSN (20 mg/kg) resulted in a significant reduction in lung histological alterations, such as the infiltration of inflammatory cells, alveolar wall thickening, and bronchial tube constrictions. These findings supported the therapeutic properties of TSN. Representative H&E-stained lung sections from each experimental group are shown in Figure 7. Group I (control) displayed normal alveolar architecture with intact bronchial walls and minimal cellular infiltration. In contrast, Group II (MP-infected) exhibited marked inflammatory cell infiltration, alveolar septal thickening, and significant alveolar space reduction, indicating severe pneumonia. Group III (MP + TSN-treated) showed improved alveolar integrity, reduced inflammatory infiltration, and attenuated edema compared to Group II. Effect of TSN on the Lung Histopathology of the Experimental Mice. Representative H&E-Stained Lung Sections From Experimental Mice (Original Magnification 200×). Group I (Control): Normal Lung Histology With Clear Alveolar Spaces and Minimal Inflammation. Group II (MP-Infected): Severe Inflammatory Cell Infiltration, Alveolar Collapse, and Interstitial Edema. Group III (MP + TSN-Treated): Partial Restoration of Alveolar Architecture and Reduction in Inflammatory Infiltration
To provide objective support for histological differences, a semiquantitative lung injury scoring system was used. Lung tissue was evaluated for four parameters: Alveolar wall thickening, Hemorrhage, Inflammatory cell infiltration, and Alveolar structure disruption. Each parameter was scored on a scale of 0-4 (0 = normal, 4 = severe), and the total lung injury score was calculated as the sum. The mean ± SEM scores were: Group I: 1.3 ± 0.2, Group II: 9.1 ± 0.5, Group III: 4.2 ± 0.4, Statistical analysis showed that TSN treatment significantly reduced lung injury scores compared to the MP group (P < .01), indicating histological protection.
Results of Molecular Docking Analyses
The Table Detailed the List of Targets, PDB ID, Binding Affinity (kcal/mol), RMSD (Å), and its Interacted Residues With the Respective Ligand Toosendanin and its Type of Bonds

Note:pa- pi alkyl; pc- pi cation; ch- carbon-hydrogen bond; pps - pi-pi stacked; ps – pi-pi sigma.
IL-1β
The TSN interacted with the residues of IL-1β including TYR24, LYS74, and GLN81, via the hydrogen bonds and with other residues, such as LEU80, TYR24, and PRO23, via a carbon-hydrogen bond. Additionally, the residue THR79 interacted with IL-1β using a pi-sigma bond. Also, the hydrophobic residues such as GLU25, LEU82, GLU83, PHE133, and LEU134 surrounded the IL-1β with a binding affinity of −6.6 kcal/mol and the estimated RMSD 2.364 Å (Figure 8A–B & Table 1). The Docking Pose of the Selected Targets IL1 (A, B), IL6 (C, D), TGF-β1 (E, F), and NF-kB (G, H) With TSN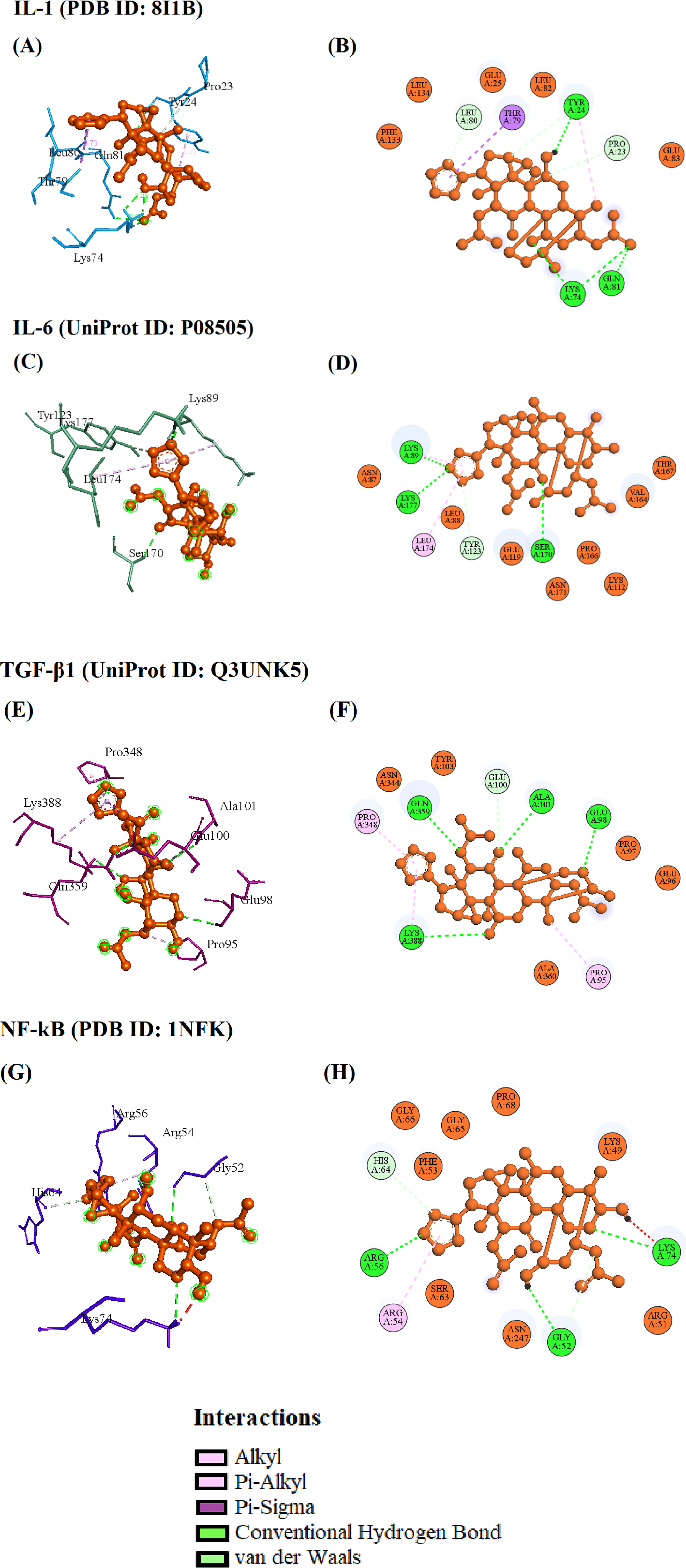
IL-6
Il-6 residues such as LYS89, LYS177, and SER170 interacted with the TSN via the hydrogen bond, and the TYR123 and LEU174 interacted via the carbon-hydrogen and pi-alkyl bonds. Also, residues such as ASN87, LEU88, LYS112, GLU119, ASN171, VAL164, PRO166, and THR167 surrounded the IL-6 and TSN docked complex with the binding affinity of −6.5 kcal/mol and the RMSD of 2.671 Å (Figure 8C, D & Table 1).
TGF-β1
The TSN formed four hydrogen bonds with GLU98, ALA101, GLN359, and LYS388 residues of TGF-β1. The other residue, GLU100, interacted via a carbon-hydrogen bond. Also, the residues PRO95 and PRO348 interacted with the TSN via pi-alkyl bonds. Likely, the residues such as GLU96, PRO97, TYR103, ASN344, and ALA360 surrounding the complex of TGF-β1-TSN with the binding affinity of −7.3 kcal/mol and the RMSD of 2.411 Å (Figure 8E, F & Table 1).
NF-kB
The TSN formed three hydrogen bonds with the residues such as GLY52, ARG56, and LYS74. Also, the carbon-hydrogen and pi-alky bond formed with the residues GLY52, HIS64, and ARG54. Meanwhile, the hydrophobic residues such as LYS49, ARG51, PHE53, SER63, GLY65, GLY66, PRO68, and ASN247 surrounded by the NF-kB-TSN complex with the binding affinity of −6.7 kcal/mol and the RMSD of 2.074 Å (Figure 8G, H & Table 1).
Molecular docking studies revealed that TSN exhibited strong binding affinities with all four inflammatory targets, with the highest affinity observed for TGF-β1 (−7.3 kcal/mol). Binding affinities ranged from −6.5 to −7.3 kcal/mol, indicating favorable interactions. RMSD values (2.074-2.671 Å) suggest reliable docking poses. • TSN–IL-1β (PDB ID: 8I1B): Binding affinity of −6.6 kcal/mol; interactions observed with key residues TYR24, LYS74, and GLN81, located within the receptor’s active site region. • TSN–IL-6 (UniProt: P08505): Affinity of −6.5 kcal/mol; hydrogen bonding with LYS89, SER170, and LYS177, suggesting interaction near the receptor-binding interface. • TSN–TGF-β1 (UniProt: Q3UNK5): Strongest binding at −7.3 kcal/mol; interaction with GLU98, ALA101, GLN359, consistent with previously defined ligand-binding pocket residues. • TSN–NF-κB (PDB ID: 1NFK): Affinity of −6.7 kcal/mol, with binding at residues GLY52, ARG56, LYS74, within the DNA-binding domain region, suggesting potential interference with transcriptional activity. Collectively, these results suggest that TSN predominantly interacts at functionally relevant active or binding interface regions, rather than remote allosteric sites.
Discussion
MP is currently acknowledged as the tiniest prokaryotic microbe that can survive on its own without a host cell. It is also the primary cause of pediatric CAP. On a global scale, outbreaks of MPP occur every 3-7 years, and during these epidemic years, they are responsible for about 40% of cases of pediatric CAP. 24 Previously, it was thought that MPP typically occurred in children aged five and older with minor symptoms. Recent clinical observations have shown that there is a growing tendency for MPP to occur at a younger age. As a result, this poses a considerable risk to the children’s health. Regrettably, the management of pneumonia in current clinics is hindered by drug resistance and the ineffectiveness of present therapy options. Additionally, there is a lack of understanding of the molecular pathways that induce lung damage following infection, further complicating the situation. 25 Hence, a thorough investigation has been carried out to clarify the mechanisms underlying tissue damage induced by pneumonia and to provide an innovative therapy strategy. Eventually, the findings of the present work highlighted that TSN effectively ameliorated the pneumonia conditions in the MP-induced mice.
The escalation of immune reactions and heightened release of immune markers, along with the occurrence of immune cells, might lead to damage to the respiratory system. 26 Furthermore, the precise pathological development of pneumonia caused by MP infection is still not fully understood. The earlier report indicated the presence of various immune cells, such as macrophages and neutrophils, in the pneumonia lesions. 27 The release of proinflammatory cytokines during MP infection is associated with several types of immune cells, specifically macrophages. 28 Neutrophils are vital immune cells that have essential roles in innate immunity, which is responsible for promoting inflammatory responses. Neutrophil infiltration is commonly regarded as a significant occurrence in MPP. Earlier studies have indicated that neutrophil levels were considerably increased in both the blood and BALF of patients with corticosteroid-resistant refractory pneumonia. 29 The excessive concentration of neutrophils can result in a hyperinflammatory state caused by the release of pro-inflammatory regulators. 30 The present results proved that the TSN treatment successfully reduced the cell counts in the BALF of MPP mice, which may be due to the TSN-mediated reduction in the inflammatory cells.
The immune response encompasses both humoral and cellular immunity. Humoral immunity is the process by which B cells are activated and bind to antigens, including microbes, viruses, and other foreign substances. This leads to the development and function of specific antibodies. 31 In previous studies, the changes in serum-specific antibodies during MP infection were well reported. They found that the majority of patients produce MP-specific antibodies of the types IgG, IgM, IgA, and IgE. The current results highlighted that the TSN treatment remarkably reduced MP-mediated Ig-M production in the MPP mice. These findings proved that TSN successfully reduced the inflammatory response in the mice with MPP.
Cytokines, which play a role in immune response, function as connectors within the immune system to facilitate the synchronization of various reactions in order to protect the body from infections. 32 The pathogen’s membrane lipoprotein can trigger local cytokine synthesis in tissues with a high concentration of cytokine-producing cells, leading to inflammatory tissue damage.33,34 Mounting evidence has verified that pneumonia caused by MP is strongly linked to an increased synthesis of proinflammatory cytokines. 35 These chemicals are not just byproducts but rather pivotal agents that trigger a sequence of intricate immunological reactions. 36 The activation of inflammatory cytokines can trigger an innate immune reaction in the pulmonary tissues. It was well known that the various cytokine levels were increased in the BALF of MPP patients. 37 Serum analysis revealed alterations in cytokine levels, specifically IL-1, IL-6, and IL-18. 38 IL-6 can cause immature B cells to develop into cells that produce antibodies, stimulate the activation of T-cells, and play an essential role in the immunological response of the body. In mycoplasmal pneumonia, these inflammatory mediators are essential in breaking down the alveolar matrix. 39 The cytokine release, such as IL-1, IL-6, and IL-8, during MP infection is connected with the worsening of pneumonia symptoms. Fluctuations in cytokines may serve as one of the disease-causing mechanisms of MP infection. 40 In this current study, the findings indicate that the TSN treatment successfully diminished these cytokines in the mice with MPP. These results highlighted that TSN has strong anti-inflammatory activity.
NF-κB is a crucial transcription factor that exists in an inactive state in all cells and can be activated in response to different stimuli. Therefore, it is considered a primary responder to various internal and external stimuli, such as free radicals, cytokines, and infections. In addition, NF-κB plays a vital role in regulating the immunological response to infections. 41 Infection with MP activates NF-κB signal transduction in macrophages. This results in the release of immunomodulatory chemicals and other cytokines. These substances can enhance the immune reaction and participate in the progression of pneumonia. The pro-inflammatory regulators play a critical role in the inflammation triggered by MP. Prior studies have shown that the cytokine levels in the lung tissues were significantly increased in MP-infected animals. 42 As a result, samples of drugs and phytocompounds that hinder the activation of NF-κB could potentially serve as anti-inflammatory agents in treating MPP. 43 Similarly, the present findings demonstrated that the NF-κB levels in the lung tissues of MPP mice were successfully reduced by the TSN treatment, which proves the anti-inflammatory properties of the TSN.
Pneumonia is significantly influenced by oxidative stress in its pathogenesis. Studies have demonstrated that individuals diagnosed with pneumonia exhibit heightened oxidative stress levels. The presence of MP infection results in elevated oxidative stress levels in the lungs. This increase is a significant factor in the host’s innate immune reaction to foreign pathogens. 44 The disruption of the oxidative equilibrium caused by excessive secretion of free radicals and/or suppression of antioxidants can amplify the intensification of inflammation and cellular demise. 45 Typically, host cells include antioxidant defense systems that enable them to maintain an equilibrium between oxygen and free radicals in their metabolism. 46 Cellular damage can occur due to impaired mitochondrial activity and a reduction in levels of antioxidants such as SOD and GSH, which happen in pathological situations as a result of oxidative stress induced by an excess of oxygen-free radicals. Oxidative stress is commonly associated with an elevation in the production of MDA and ROS. The primary cause of cell injury is the disturbance of many signal transduction pathways caused by an elevation in oxidative stress. 47 Prior knowledge indicated that individuals diagnosed with CAP exhibited elevated levels of oxidative stress compared to individuals without the condition. 48 Similarly, the present results also demonstrated that the mice with MPP had elevated MDA levels, whereas diminished SOD and GSH levels were noted. Remarkably, the administration of TSN successfully elevated the GSH and SOD levels while concurrently reducing the levels of MDA, thus providing evidence for the antioxidant capabilities of TSN.
The molecular docking analysis confirmed a significant interaction between the defined targets and the TSN. The TSN interacted more with the TGF-β1 than other targets based on the binding energy (−7.3 kcal/mol) and the number of hydrogen bonds (#4) involved. In the previous study, Deng et al stated that targeting TGF-β1 signaling has therapeutic potential in diseases with excessive inflammation or immune dysregulation. 49 It is likely because the induced TGF-β signal pathway activation in the lung stimulates chronic, aggressive fibrosis in adult rats.
Limitations of the Study
This study provides promising evidence for the protective effects of Toosendanin (TSN) against Mycoplasma pneumoniae-induced pneumonia in mice through suppression of NF-κB-mediated inflammatory responses. However, several limitations should be acknowledged. First, a formal sample size calculation was not conducted; instead, animal group sizes were based on precedents from similar studies. Although the sample size (n = 6 per group) was sufficient to detect significant differences, a power analysis would enhance the robustness and reproducibility of the findings. Second, while semiquantitative histopathological scoring was incorporated, further objective quantification using immunohistochemistry (e.g., for NF-κB p65 nuclear localization or inflammatory markers) would provide stronger mechanistic support. Third, the dose-response relationship of TSN was not explored. Only a single dose (20 mg/kg) was tested, which limits the ability to determine the optimal therapeutic window or potential toxicity thresholds. Fourth, in silico docking predictions were not experimentally validated using molecular assays such as protein binding or activity inhibition studies. Therefore, the computational findings should be interpreted as supportive but preliminary. Fifth, the study focused primarily on acute inflammation. Long-term effects of TSN on lung remodeling, fibrosis, or chronic infection resolution were not assessed.
Finally, while the study confirmed the anti-inflammatory potential of TSN in vivo, mechanistic studies using knockout mice or NF-κB-specific inhibitors could further validate the direct involvement of this signaling pathway. Future research should address these limitations to strengthen the translational relevance of TSN as a therapeutic candidate for MPP. Concurrently, it has also been noted that people with interstitial lung fibrosis express more TGF-β1. In this condition, TSN can be a potential compound to inhibit the over-activation of TGF-β1 under certain diseased conditions.
Conclusion
The present results confirm the beneficial benefits of TSN in protecting mice from MPP. TSN administration led to a decrease in lung weight, lower inflammatory cytokines and MDA levels, increased levels of antioxidants, and decreased expression of NF-κB in mice with MPP. The data demonstrate the therapeutic efficacy of TSN in treating MPP. Thus, TSN has the potential to be an effective therapeutic drug for treating MPP. In the future, the TSN can be targeted to treat as a combination therapy after validating with extensive In vitro and In vivo experiments.
Supplemental Material
Suppplemental Material - Toosendanin Attenuates Mycoplasma pneumonia Induced Pneumonia (MPP) in Mice via Inhibiting NF-κB-Mediated Inflammatory Response: In vivo and silico Studies
Supplemental Material for Toosendanin Attenuates Mycoplasma pneumonia Induced Pneumonia (MPP) in Mice via Inhibiting NF-κB-Mediated Inflammatory Response: In vivo and silico Studies by Yin Zhou, Xinyou Su, Cuicui Wang, Abdullah A. Alarfaj, Abdurahman Hajinur Hirad in Dose-Response
Footnotes
Acknowledgment
The authors would like to express their sincere gratitude to the institutions and laboratories that supported this research. We are especially thankful to King Saud University for providing the facilities and resources necessary for the in silico studies. We also acknowledge the valuable contributions of the staff at Ankang People’s Hospital, the Central Hospital affiliated with Shandong First Medical University, and the Affiliated Hospital of Yan’an University in supporting the in vivo experiments.
Ethics Approval
The study was approved by the Ethics Committee of Central Hospital Affiliated to Shandong First Medical University, Shandong Province, China (WQEE-2023-153).
Authors Contributions
Yin Zhou: investigation, formal analysis; Xinyou Su: data curation, supervision, Cuicui Wang: software, project administration; Abdullah A. Alarfaj: data curation, supervision; Abdurahman Hajinur Hirad: software, project administration; Xiaogang Li: Conceptualization, methodology, and writing – review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors extend their appreciation to the Ongoing Research Funding Program (ORF-2025-677), King Saud University, Riyadh, Saudi Arabia.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data presented in this study are available on request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.