
Abstract
Context
The liver is the organ by which the majority of substances are metabolized, including psychotropic drugs. Lithium (Li) used as drug for many neurological disorders such as bipolar disorders.
Objective
This study aims to assess lithium toxicity and to evaluate the hepatic-protective properties of a grape skin seed and extract (GSSE).
Materials and methods
Twenty-four male Wistar rats were exposed for 30 days to either various lithium concentrations, GSSE alone, or lithium supplemented with GSSE. The proteomic analysis revealed alterations of liver protein profiles after lithium treatments that were successfully identified by mass spectrometry.
Results
Lithium treatment induced an oxidative damage by the alteration of antioxidant enzymes activities such as superoxide dismutase, CAT, and Gpx. The regulated proteins are mainly involved in the respiratory electron transport chain, detoxification processes, ribosomal stress pathway, glycolysis, and cytoskeleton. Proteins were differentially expressed in a dose-dependent manner. Interestingly, GSSE reversed the situation and restored the level of liver proteins whose abundance was modified after lithium treatment, arguing for its protective activity.
Conclusion
Our data demonstrated the ability of proteomic analysis to underline the toxicity mechanisms of lithium in animal models. Based on these results, GSSE may be envisaged as a nutritional supplement to weaken the liver toxicity of lithium.
Introduction
Lithium (Li) is a toxic metal found in industrial areas. 1 It disperses in the environment and accumulates in the trophic chain. 2 Even if its toxicity is well documented, lithium remains a drug of choice used as a psychiatric disease therapeutic, particularly in bipolar disorders.3,4 However, Li is at the origin of deleterious damages to different organs including the liver. Numerous investigations report that Li-induced hepatic structural and functional modifications, proved by disorganization in hepatic cords and vacuolization in hepatocytes. Moreover, treatment with a higher dose of Li induces fibrosis and fatty degenerations. 5 Inherently, to its xenobiotic potential, lithium induced the accumulation of hepatic enzymes involved in detoxification mechanism. 6
In such biomedical context, proteomics enables the simultaneous assessment of the abundances of hundreds to thousands of proteins, providing an unprecedented amount of molecular information.7,8 Here, this approach allows for exploring the main metabolic pathways involved and the mechanisms underlying the pathophysiological state of the liver under Li exposure with or without grape skin seed and extract (GSSE) supplementation. Previous works already revealed the potential of proteomics, for instance to assess alcohol toxicity on the liver. 9 Proteomics also aims at studying the post-translational modification of proteins, but such approach is less frequently described in the case of Li toxicity. To achieve these goals, we have applied a proteomics strategy based on nanoliquid chromatography−tandem mass spectrometry (nano-LC−MS/MS). To our knowledge, this is the first proteomic study on the effect of lithium on the liver proteome.
At the basis of its properties, GSSE contains antioxidant compounds and unsaturated fatty acids. This composition gives GSSE numerous biological and health-promoting properties, such as scavenging of reactive oxygen species, 10 but also anti-tumor 11 and anti-inflammatory properties. 12 As a result, GSSE is certified by the FDA as a healthy ingredient. 13
Material and Methods
Animals and Treatments
Forty-eight Male Wistar rats (210–230 g) were purchased from Pasteur Institute, Tunisia. Animals were used in accordance with the Medical Ethics Committee for the Care and Use of Laboratory Animals of the Pasteur Institute of Tunis, Tunisia (approval number: FST/LNFP/Pro 152012). Animals were acclimated on a 12 h light/dark cycle in animal housing with a temperature fixed at 22 ± 2°C. Twenty-four rats (8 rats per group) were exposed daily by intraperitoneal injection to 0, 2 and 100 mg/kg of Li for 30 days. To assess the protective role of GSSE, 3 groups of animals (8 rats per group) received an intraperitoneal injection of GSSE (4 g/kg), a mixture of 2 mg/kg of lithium chloride + GSSE (4 g/kg) or a mixture of 100 mg/kg of lithium chloride + GSSE (4 g/kg), respectively. Doses of Li were chosen in the therapeutic range (2 mg/kg bw) or toxic but non-lethal (100 mg/kg bw) according to Milutinovic 14 (2016). GSSE powder was obtained according to Mezni et al 15 (2017) protocol.
After 30 days of exposure, rats were decapitated and their liver was collected, weighed and frozen in liquid nitrogen. Homogenization was conducted in lysis buffer (urea 7 M, thiourea 2 M, CHAPS 2%, C7BzO .5%, DTT 20 mM, Tris-HCl 50 mM) containing a protease inhibitor cocktail (Roche Diagnostics) using an ultra-thurax for homogenization. The homogenates were then sonicated for 1.5 min (15% amplitude for 3 cycles of 30 s each), these extraction steps being carried out on ice. After debris elimination by centrifugation (10 000 g for 20 min at 4°C), supernatants were assembled and processed with a 2-D Clean-Up Kit (GE Healthcare) for protein precipitation in order to remove interfering compounds and protein levels were determined using the Bradford 16 method.
Protein Electrophoresis and In-Gel Digestion
For each sample, protein electrophoresis was set up according to Obry et al 17 (2014). In brief, proteins extracts were loaded and allowed for migration in an SDS-PAGE stacking gel (7%). After staining with Coomassie, protein band was excised and sequentially immersed in a reductive, and then in an alkylating medium. The protein bands were further submitted to trypsin digestion. Finally, peptide fragments were combined and evaporated.
Analytical Determination
Nano-LC-MS/MS analysis was detailed in a previous paper. 15 Briefly, LTQ Orbitrap coupled with nano-LC equipped with a C18 column was used to separate the peptide mixture and perform MS analysis. The mass spectrometer was operated in a data-dependent mode to automatically switch between Orbitrap MS (from m/z 300 to 2000 with a resolution of 30 000) and LTQ-MS/MS acquisition. For fragmentation, the spectrometer selected the 20 most intense peptide ions. Identifications were performed by implementing a Mascot search against the Rattus norvegicus database.
Antioxidant Enzymes and Histopathology
The liver supernatant was used to determine the antioxidant enzymes activities, such as Superoxide dismutase (SOD) (E.C.1.15.1.1) activity was determined by using modified epinephrine assay (Misra and Fridovich, 18 1972). At alkaline pH, superoxide anion O2− causes the auto-oxidation of epinephrine to adenochrome; while competing with this reaction, SOD decreased the adenochrome formation. One unit of SOD is defined as the amount of extract that inhibits the rate of adenochrome formation by 50%. Enzyme extract was added in 2 mL reaction mixture containing 10 μl bovine catalase (.4 U/μl), 20 μl epinephrine (5 mg/mL), and 62.5 mM sodium carbonate/sodium bicarbonate buffer pH 10.2. Changes in absorbance were recorded at 480 nm.
Catalase (CAT) was assayed by the decomposition of hydrogen peroxide according to the method of Aebi 19 (1974). Decrease in absorbance due to H2O2 degradations was monitored at 240 nm for 1 min and the enzyme activity was expressed as μmol H2O2 consumed/min/mg protein.
The glutathione peroxidase activity (GPx) (E.C. 1. 11. 1. 9) was assessed according to the method of Nakamura. 20 Briefly, 1 mL of reaction mixture containing .2 mL of sample, .2 mL of phosphate buffer .1 M pH 7.4, .2 mL of GSH (4 mM) and .4 mL of H2O2 (5 mM) was incubated at 37°C for 1 min and the reaction stopped by addition of .5 mL TCA (5%, w/v). After centrifugation at 1500 g for 5 min, aliquot (.2 mL) from supernatant was mixed with .5 mL of phosphate buffer .1 M pH 7.4 and .5 mL DTNB (10 mM) and absorbance recorded at 412 nm. GPx activity was expressed as nmol of GSH consumed/min/mg protein.
The liver tissue sections were made with a thickness of .5 μm, colored with hematoxylin-eosin (H and E) and examined using an optical microscope. One representation micrography of liver tissue from each group is shown in Figure 1. Liver histological morphology in rats: (LD) lipid droplets, (VD) vacuolar degeneration.
Statistics and Bioinformatics Analysis
STATISTICA program (version 8.0) was used to develop the statistical analysis. All obtained results were described as mean ± of standard deviation. The observed variation was examined by one-way ANOVA (P < .05). All statistical tests were 2-tailed, and P < .05 was considered significant. (*) indicate significance for lithium groups (2 and 100 mg/kg bw) vs control (0 mg lithium/kg bw), and (§) for lithium+ GSSE vs lithium at each lithium dosage.
The software Progenesis LC-MS (Nonlinear Dynamics) was used for protein quantitation (using multivariate statistics on protein measurements) after data alignment and normalization. NCBI database was used for annotation with gene ontology (GO). Proteins displaying significantly differential abundances between the different groups of animals were analyzed using Pantherdb databases (http://www.pantherdb.org/) to highlight highly represented molecular function and also using Stich (http://stitch.embl.de/cgi/) to plot predicted protein interactions.
Results
Antioxidant Enzymes and Histopathology
Lithium exposure affected the antioxidant enzyme activities, depressing SOD activity (−43.54%; Figure 2A), CAT activity (−82.8%; Figure 2B) and GPx (68.75%; Figure 2C). Effect of GSSE on Li-induced liver superoxide dismutase (A), CAT (B), and GPx (C) activities after 30 days of exposure. Rats were treated with various doses of Li were administered daily (2 and 100 mg/kg bw) with or without GSSE (4 g/kg bw). Results are expressed as a mean ± SEM. (n = 6) P < .05 was considered significant: *for lithium vs control, and §for lithium +GSSE vs lithium.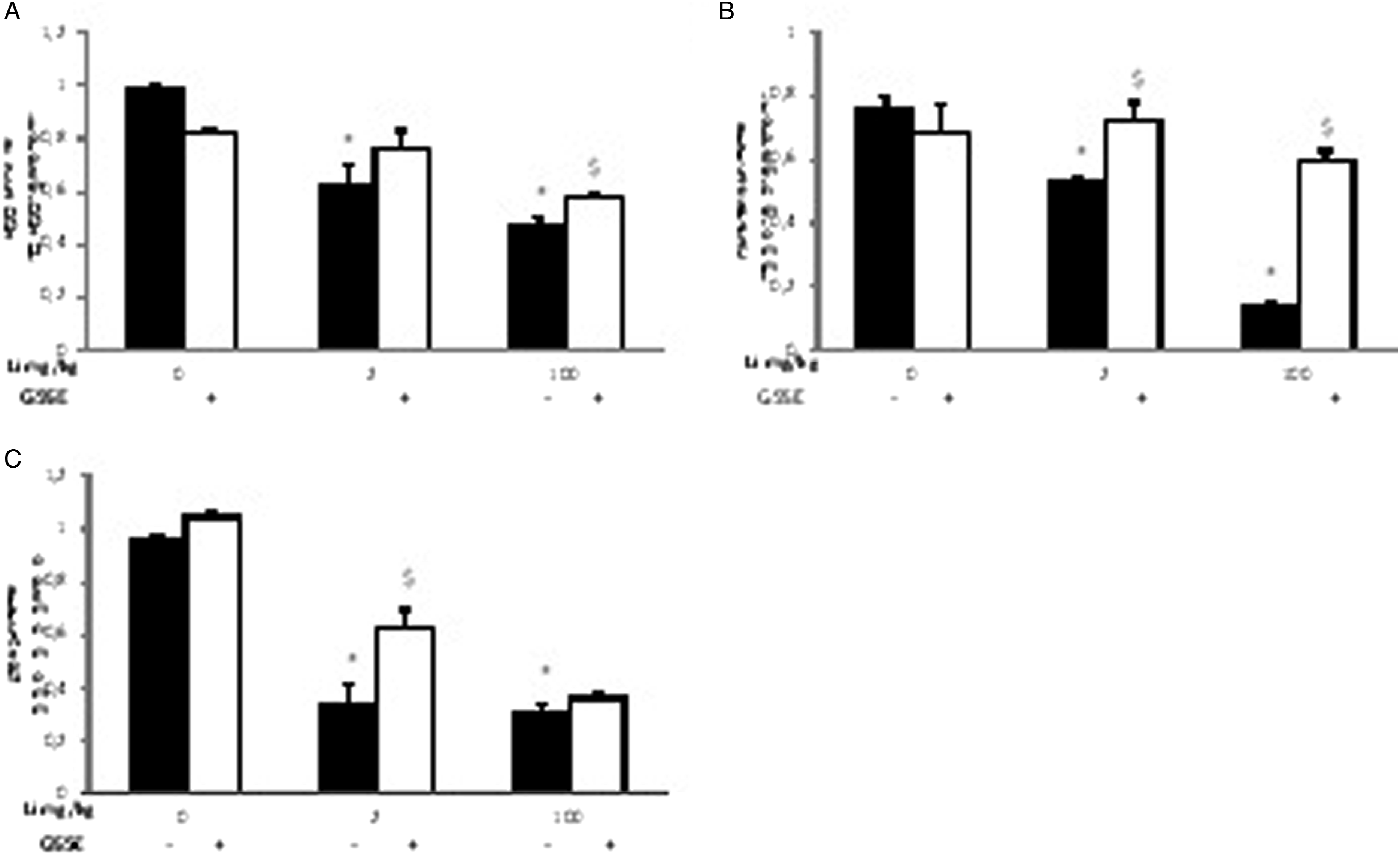
The histological experiments confirmed the protective effect of GSSE against hepatotoxicity induced by lithium (Figure 1). The histological changes were assessed using hematoxylin-eosin (HE)-stained liver tissue sections from each treatment group. The standard control (Figure 1A) and GSSE treated group (Figure 1B) displayed a typical architecture. In contrast, all rats of the Li-treated groups showed significant morphological changes, including lipid droplets (LD) and vacuolar degeneration (VD).
Evolution of Protein Profile
Uptake of lithium induced modifications of protein expression that appeared most of the time in a dose-dependent manner (Figure 3). First, according to their molecular function, 39% of differentially expressed proteins are involved in catalytic activity, 30% are associated with a structural role and 29% are binding proteins (Figure 3A). According to their cellular localization, 42.9% of deregulated proteins belong to the cell part, 14.3% are located in the extracellular region, 7.1% are involved in macromolecular protein complex, and 35.7% of proteins are found in organelles (Figure 3C). Lithium-induced alterations of the liver proteome after 30 days of exposure. Significantly regulated proteins are classified in GO categories; the results are presented as percentage of molecular function (A), biological process (B), cellular component (C), protein class (D), and pathway (E).
More specifically, the most affected protein classes were respectively ribosomal proteins accounting by about 15.6% (60S ribosomal protein L3, L5, L6, L8, L10, and L27 and 40S ribosomal protein S16), and transferases by about 15.6% (isovaleryl-CoA dehydrogenase, creatine kinase, acetyl-CoA C-acyltransferase, betaine-homocysteine methyltransferase, carnitine O-palmitoyltransferase, and transketolase). In addition, the oxidoreductases represent 13.3% (isovaleryl-CoA dehydrogenase, formyltetrahydrofolate dehydrogenase, betaine-homocysteine methyltransferase, transketolase, cytochrome C oxidase, and dehydrogenase/reductase), the cytoskeletal proteins 11.1% (tropomyosin beta chain, vimentin, troponin C, troponin I and myosin), transfer/carrier proteins 8.9% (sideroflexin, myoglobin, serum albumin, and glutathion-S-transferase). Less significantly represented protein classes were also present (Figure 3D), some proteins potentially belonging to multiple protein classes.
List of Identified Proteins that Displayed Differential Abundances Between Exposure to Lithium Alone or Supplemented to Grape Skin Seed and Extract.

Li affects also the calcium-dependent proteins expression (such as troponin C, myosin light chain) or oxidoreductase proteins (isovaleryl-CoA dehydrogenase, formyltetrahydrofolate dehydrogenase, transketolase; betain-homocysteine S-methyltransferase; dehydrogenase/reductase; cytochrome C oxidase), or proteins involved in glycolysis pathway (such as fructose bisphosphate aldolase), or also of nucleic acid-binding proteins (such as 60S ribosomal protein L3, 60S ribosomal protein L5, 60S ribosomal protein L6, 60S ribosomal protein L8, 60S ribosomal protein L10, 60S ribosomal protein L27, 40S ribosomal protein S16).
Interaction Network Analysis
The abundance of proteins involved in drug detoxification process in the liver was also investigated (Figure 4). These proteins displayed an increased abundance under Li exposure (such as GST kappa, GST mu, epoxide hydrolase, cytochrome P450 2A2, cytochrome P450 2C11). Protein-protein interaction network (STRING) of proteins involved in complex of detoxification. Gstm1 (A) and Ephx (B).
Interactions of these proteins with other protein partners were obtained by searching the STRING online database (Figure 4). The presented protein functional association networks are the result of the protein entries: “Epoxide hydrolase” (Ephx) and “Cytochrome P 450 2A2” (Cyp2a2). Interestingly, Ephx binds directly to proteins such as gluthation-S-transferase (Gstm1, Gsto1, Gsta4, Gstt1, Gstt2, and Mgst1) and cytochrome P450 (Cyp1a1, Cyp1a2, and Cyp 2e1). Cytochrome P 450 2A2 binds directly to proteins such as cytochrome P450 (Cyp 4a2, Cyp 4a1, Cyp 4a8, Cyp 2c7, and Cyp 4f1), 3 beta-hydroxysteroid dehydrogenase (hsd3b and hsd3b5), epoxide hydrolase 2 (Ephx 2), gluthation-S-transferase theta 2 (Gstt2), and UDP-glucuronosyltransferase 2B15 (Ugt2b15).
In addition, a more global network was represented (Figure 5), which enables the visualization of essential regulations at multiple levels (protein-protein interactions and protein-chemical interactions). In addition to previously discussed proteins, this also pointed out proteins involved in transition metal ion binding (Gstm1, Alb, Cp, and Cpa2), in lipid oxidation (Cpt2, Ivd, Hadha, and Hadhb), in muscle system processes (Myl1, Tnni2, Tnnc2, Acta1, Tmp2, and Aldolase). Interestingly, one can notice that a very large fraction of regulated proteins has a direct or indirect relationship with ROS, mainly with oxygen and hydroxyl radicals, such as proteins directly involved in oxidative stress mechanism and associated antioxidant properties. Interaction network with STITCH (protein–protein interactions, protein-chemical interactions, and interactions with hydroxyl and oxygen radicals).
Hepatoprotective Effect of Grape Skin Seed and Extract
Annotation of Proteins that Displayed Modified Abundances.

Furthermore, the treatment with Li also down-regulated proteins involved in oxidative stress and detoxification systems (such as GST1, Gstk, and up-regulated Ephx, Cyp2a2, and Cyp2c23); their level was re-established under GSSE exposure. The same also applies to cytoskeletal proteins (tropomyosin chain beta, vimentin, troponin C, troponin I, and myosin light chain), whose level was restored after co-treatment with GSSE. In addition, lithium treatment affects the abundance of many oxidoreductases (such as isovaleryl-CoA dehydrogenase, formyltetrahydrofolate dehydrogenase, transketolase, betain-homocysteine S-methyltransferase, dehydrogenase/reductase, and cytochrome C oxidase) and for this class of proteins, GSSE re-established the abundance of these proteins.
Discussion
Lithium has been classified by the FDA 13 as a mood stabilizer and drug for the treatment of bipolar disorder. 21 However, many side effects represent an obstacle to a more widespread clinical use. 22 Our work highlighted the effects of Li exposure on liver protein profile and the protective effect afforded by grape extract co-administration.
The treatment of healthy rats with various doses of Li altered the expression of proteins involved in the respiratory electron transport chain. More specifically, Li induced an increase of abundance of the terminal oxidase of the mitochondrial respiratory chain, 23 regulating oxidative phosphorylation. 24 In addition, Li treatment also up-regulated ATP synthase D. Almost all functions of mitochondria are either directly or indirectly linked to the oxidative phosphorylation machinery and energy coupling. The central part of this machinery, located in the inner mitochondrial membrane, comprises the four-electron transfer chain complexes (complexes I, II, III, and IV) and ATP synthase (complex V). 25 Lithium induced mitochondrial membrane potential decrease resulting in ROS formation. 26 The goal of oxidative phosphorylation is the generation of a proton gradient within the inner mitochondrial membrane, and at the end to synthesize ATP. Since this process is not completely efficient, electron transfer to O2 may occur at complex I or III, resulting in the generation of reactive oxygen species (ROS). 27
In this regard, many GST protein abundances were decreased after lithium treatment. These proteins constitute the primary regulators of the reduced/oxidized glutathione ratio in cells and are commonly used to measure of cellular redox potential and stress. 28 Lithium reduces the GST, glutathione (GSH) and glutathione peroxidase (GPx) levels. 29 Low levels of GSTs may render animals less protected against oxidative stress, the lower levels being related to either lower production or higher consumption of GST in detoxification processes. 30 Lithium treatment induced oxidative stress by inhibiting all antioxidant enzyme activities such as SOD, CAT, and GPx activities. Previous studies also revealed that the liver is one of the main targets for lithium toxicity. Although, the cellular and molecular mechanisms involved in the lithium induced liver toxicity have not yet been wholly understood. 26 One of the most robust mechanistic hypotheses in lithium-induced hepatotoxicity is oxidative stress. 6 Lithium reduced the glutathione (GSH) levels and also increased lipid peroxidation and the activities of antioxidative enzymes, which included catalase, glutathione S-transferase, and superoxide dismutase. 31 Other research reported that lithium creates oxidative damage on the liver by generating reactive oxygen species (ROS), increasing lipid peroxidation, and reducing antioxidant enzyme activities (SOD, CAT, and GPx). 32 In addition, a significant reduction of CAT activity was noticed in Li-treated rats, which could be explained by the overconsumption of this enzyme involved in the conversion of H2O2 to H2O. The antioxidant enzyme SOD, showed a significant decrease in the liver after lithium treatment, resulting in free radical scavenging mechanism loss. 33 SOD is also known for its role in inhibiting OH production by scavenging O2 and thus leads to a decrease in the initiation of lipoperoxidation (LPO). 34
Interestingly, lithium also up-regulated epoxide hydrolase and decreased the abundance of the P450 cytochromes which constitute the significant enzyme family capable of catalyzing the oxidative biotransformation of most drugs and xenobiotics and are therefore of particular relevance in the context of clinical pharmacology.35-37 By generating reactive epoxides, epoxide hydrolase is an enzyme that metabolizes xenobiotics into water-soluble compounds for excretion. 38 These proteins are described to complement each other in detoxifying a wide array of mutagenic, toxic, and carcinogenic xenobiotics 39 and are called drug metabolizing enzymes (DMEs), involved in the biotransformation of associated substrates.40,41 The EH deficiency or pharmacological inhibition attenuates ER stress and favorably impacts metabolic homeostasis, and reduced fibrosis. 42 In our study, lithium induced an up-regulation of this protein, so the ER stress and inflammation were the result of the alteration of the abundance of EH and P450 cytochromes.
In addition, numerous ribosomal proteins (RP) were affected by lithium treatment, and there was a general trend for a decrease in ribosome protein abundance associated with exposure. In contrast, the level of RPL5, a cytosolic ribosomal protein, increased with lithium exposure. Remarkably, it has been proposed that a defect in a RP causes increased levels of free RPL5 that has been already described in other contexts to activate p53. In addition, RP suppressed the progression of experimental hepatic fibrosis accompanied by a reduction of Akt phosphorylation, and their overexpression reduced the phosphorylation of GSK3b. 43 The deficiency of a ribosomal component triggers a pathological condition, known as ribosomal stress, characterized by activation of p53, leading to apoptosis. 44 Ribosomal proteins are involved in DNA repair, cell cycle arrest, apoptosis and proliferation, and are moderately expressed in various cancers. 45
Another critical point was modifying the expression level of several proteins associated with glycolysis. Lithium treatment has been reported to disrupt glycolysis and gluconeogenesis by altering relevant enzyme activity. 46 Among these proteins, fructose bisphosphate aldolase A was down-regulated and enolase up-regulated after lithium exposure. Fructose bisphosphate aldolase is involved in the fourth step of glycolysis, catalyzing the cleavage of fructose 1,6-bisphosphate into glyceraldehyde 3-phosphate. 47 Enolase is a key glycolytic enzyme that catalyzes the dehydration of 2-phosphoglycerate to phosphoenol pyruvate in the last step of the catabolic glycolytic pathway. 48 Because the glycolytic pathway is the primary energy source for many cells, the effect of lithium on these proteins suggests that it might both disturb energy metabolism. Moreover, lithium induced an increase of the abundance of transketolase, reported to be involved in the reductive CBB cycle (Calvin-Benson-Bassham cycle) and in the non-oxidative part of the pentose phosphate pathway, thus playing a critical role in connecting the pentose phosphate pathway to glycolytic intermediates. 49
Furthermore, lithium treatment also induced a modified abundance of proteins involved in lipid oxidation. Here, it was found that CPT, Slc27, and Ivd levels were decreased upon lithium treatment, leading to an accumulation of intracellular lipids and insulin resistance in insulin-targeting cells. 50 Noteworthy that restricted production of Slc27 lead to liver steatosis. 12 Moreover, fatty acid oxidation has been considered a significant contributor to the development of hepatic steatosis and the progression of non-alcoholic fatty liver disease. 51 In addition, lithium altered abundance of pancreatic lipase (Pnlip), a protein secreted by the acinar pancreas, essential for the hydrolysis and absorption of long-chain triglyceride fatty acids. 52
Another group of proteins regulated by lithium treatment participates in cytoskeleton organization and dynamics. Indeed, in our proteomics study, the levels of the cytoskeletal proteins dramatically decreased in the treated group. These changes might be associated with disruption of liver metabolism and function. Furthermore, the cytoskeleton is an essential structural element of all cell types, playing a pivotal role in the maintenance of the cell architecture, adhesion, migration, differentiation, division, and organelle transport. Accordingly, the alteration of cytoskeletal gene expression is highly likely to be associated with cytoskeleton structural disruption, potentially leading to cell death. 53
From another point of view, the cytoskeletal organization is crucial for normal mitochondrial morphology, motility, and function. In addition, the cytoskeleton is involved in mitochondrial respiration and energy fluxes. 54 The altered abundances of cytoskeletal proteins and mitochondria dysfunction likely result from the altered mitochondrial oxidative phosphorylation (OXPHOS) system and ensuing ROS formation, which plays a crucial role in the disruption of cytoskeleton organization. 55
In facts, Lithium induced also structural changes in liver tissue showed in histological sections by including lipid droplets (LD) and vacuolar degeneration (VD). Our results corroborated previous findings reported by Sharif et al (2011), 56 who indicated that alkali metal causes histopathological and enzymatic changes in rats. The study of the Ben Saad et al 32 (2017) showed that the liver of animals treated with lithium are characterized by an important sinusoidal dilation, congested central veins, vacuolization, and inflammatory cell infiltration
The most relevant result drawn from the current study is the hepatic protection afforded by GSSE against lithium toxicity. GSSE treatment abrogated almost all alterations such as oxidative stress, disruption of the cytoskeleton, mitochondrial dysfunction, and metabolism. Our study paralleled the data of Muzik and coworkers, 57 who used selenium as protective agent against lithium hepatotoxicity. Our data are the first to demonstrate the potent protection of a natural extract as GSSE against lithium hepatotoxicity. In our case, high dosage GSSE reaching 4 g/kg was highly safe when used in various experimental settings as nephroprotection against doxorubicin, 58 neuro-protection against lithium exposure, 15 cardio-protection against arsenic, 59 lung protection when subjected to bleomycin 60 or protection of pancreas submitted to fat treatment. 61
In conclusion, GSSE exerts potent protection against lithium-induced liver toxicity in healthy rats, even at high concentrations. Even in the cases where lithium has the ability to strongly interact with the proteins; GSSE injection may prevent the significant interaction between lithium and the proteome. We can consider that GSSE, through the regulation of damage induced by lithium, can be considered as a potential hepatoprotector, able to minimize the impact of lithium in humans through proteomic stability. Consequently, GSSE may be considered as an adjuvant to therapeutic doses of lithium for avoiding its harmful effects.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the grants from the Ministry of Higher Education of Tunisia.