
Abstract
Most studies of the biological effects of ionizing radiation have been done on a single acute dose, while clinically and environmentally exposures occur under chronic/repetitive conditions. It is important to study effects of different patterns of ionizing radiation. In this study, a rat model was used to compare the effects of repetitive and acute exposure. Groups: (I) control, (II, III) were exposed to fractionated doses (1.5 GyX4) and (2 GyX4), respectively/24h interval, and (IV, V) were exposed to 6 Gy and 8 Gy of whole-body gamma irradiation, respectively. The gene expression of MAPT and tau phosphorylation increased in all irradiated groups but the gene expression of PKN not affected. TGFβ% increased at dose of 2 GyX4 only. In addition, the cell cycle was arrested in S phase. Micronucleus (MN) increased and cell proliferation decreased. In conclusion, the dose and pattern of ionizing radiation do not affect the MAPT and PKN gene expression, but TGF-β, p-tau, MN assay and cell proliferation are significantly affected. The dose of 2 GyX4 showed distinctive effect. Repetitive exposure may increase TGF-β%, which causes radio-resistance and, G2/M delay. Thus, the cell cycle could be regulated in a different manner according to the dose and pattern of irradiation.
Keywords
Introduction
Everybody is regularly exposed to different amounts of various sorts of ionizing radiation, such as cosmic rays, which cannot be avoided. 1 The major sources of these exposures are medical imaging and radiotherapy. Even though ionizing radiation has many benefits in medicine, specifically in the means of prevention, diagnostics, and therapy of various diseases, it may also lead to serious health problems. 2 Radiation accidents can happen despite strict regulations and safety standards. Unexpected radiation exposure (1-10 Gy) may occur, resulting in severe radiation effects. 3 So it is important to study and understand the biological effects of different doses and patterns of ionizing radiation. Cells with a high proliferation rate are generally more sensitive to radiation. As a result, bone marrow is the most sensitive tissue to ionizing radiation because its cell turnover is among the fastest. 4
The cell cycle is an important and complicated process. Normally, it is divided into four phases: the G1 phase, in which the cell gets ready for DNA synthesis; the S phase, at which replication of DNA occurs; the G2 phase, where the cell starts preparation for mitosis; and the M phase, at which cell division takes place. Each phase has a specific length. Several controlling proteins regulate switches between phases and stop progression at checkpoints if errors are detected. Radiation is known to disturb the cell cycle in normal cells, causing affected cells to stop at checkpoints. 5
From these proteins that may have a role in cell cycle progression, tau protein, which is one of a protein group denoted as Microtubule-Associated Proteins (MAPs). It is involved in nucleic acid protection according to its cellular localization, either in the cell body, cytoplasmic membrane, or nucleus. 6 Tau protein encourages tubulin association into microtubules (the stabilizers of the cell), participating in different cellular processes such as cell division. 7 The expression of tau protein is encoded by the MAPT gene (Microtubule-Associated Protein–Tau). 8 It was revealed that ionizing radiation can promote abnormal tau phosphorylation. 9 The phosphorylation of tau has an important role in the tubulin joining of tau, as it is controlled by its phosphorylation condition. Phosphorylation of tau is regulated ordinarily by the harmonized effects of some kinases and phosphatases on tau molecules, such as protein kinase-N. A normal degree of phosphorylation is essential for the optimal functioning of tau. Besides, the biological activity of tau is lost by hyper-phosphorylation, which is associated with many neurological diseases, such as Alzheimer’s disease. 10 In addition, tau protein accumulation induces abnormal cell cycle behavior, acclimations to sustain survival, cellular alteration, and metabolic dysfunction. 11
In addition to its role in tau phosphorylation, protein kinase-N (PKN), a serine/threonine protein kinase, 12 plays a significant role in the regulation of cell division as it delays mitosis. 13 Moreover, many studies indicate that PKN is involved in a diversity of cellular processes due to its interaction with a large number of proteins. These processes include cytoskeletal regulation, cell adhesion, apoptosis, regulation of meiotic maturation and signaling to the cell nucleus, and tumorigenesis. 14
Another factor that may affect cell cycle progression and is affected by ionizing irradiation is transforming growth factor-β (TGF-β) which regulates a wide range of biological processes, including cell growth, differentiation, apoptosis, motility, immune regulation, cell invasion, and cell cycle regulation by blocking progress through the G1 phase. 15 On the other hand, the effect of TGF-β is dependent on the physiological environment. TGF-β is recognized as a modulator of late irradiation effects, and its post-irradiation response is required for DNA repair and cell cycle progression. The different TGF-β signaling can lead to either increased radio-sensitivity or increased radio-resistant. 16 TGF-β is activated by radiation-induced reactive oxygen species. 17
As an indicator of the DNA damage, which affects the cell cycle progression, the micronucleus assay and proliferation rate were used. When it comes to the cytotoxic effects of ionizing radiation, the most sensitive organ is bone marrow. In bone marrow cells, ionizing radiation causes a variety of DNA damage that may not be repaired. This damage could result in cell death or genomic instability, 4 and consequently cell cycle delay. This study aims to understand the link between the above-mentioned factors together, their effect on the cell cycle, and the effect of different ionizing radiation patterns on it.
Materials and Methods
Irradiation Source
The irradiation was done using a 137Cs unit manufactured by Atomic Energy of Canada Ltd and installed at the National Center for Radiation Research and Technology (NCRRT, Nasr City, Cairo, Egypt). Animals were held in well-ventilated cages and their movements were restricted. The animals were exposed to whole-body irradiation at a rate of .4 Gy/minute.
Animal Protocol
Healthy male rats (weighing around 180-220 g) were obtained from the animal house of the National Center of Radiation Research and Technology (NCRRT) and housed under standard laboratory conditions. The rats were provided a free standard pellet diet and water. All the study’s protocols and the animal care and handling were in accordance with the guidelines set by the Research Ethics Committee (REC-NCRRT). Rats were divided into five groups, with seven animals in each one. Group (I) control was not irradiated and groups (II) and (III) were exposed to 6 Gy and 8 Gy of whole-body gamma irradiation as a fractionated doses (1.5 GyX4) and (2 GyX4) with 24 hr interval, respectively. Groups (IV) and (V) were exposed to 6 Gy and 8 Gy of whole-body gamma irradiation as acute doses, respectively. Rats were sacrificed 24 hr after last irradiation dose under anesthesia, and the bone marrow was collected from the femur. At the end of the experimental period, 24 hr after irradiation, there was no perceived lethality. Whereas the applied dose of γ-irradiation of 8 Gy was reported as a lethal dose 11 days post-irradiation, 18 6 Gy was chosen as a sub-lethal dose, and these doses were fractionated into 2 Gy and 1.5 Gy, respectively, as mild doses 19 ; thus, the selected irradiation doses (lethal, sub-lethal, and mild repetitive) were suitable for initial screening and evaluation of the effect of radiation in the present study. The whole procedure and the animal handling were done in accordance with the guidelines regularities approved by the Research Ethics Committee (REC-NCRRT) (1A/21).
Quantitative analysis of MAPT and PKN mRNA expression by real-time PCR:
Total RNA Extraction
Total RNA was extracted from tissue homogenate of bone marrow using the SV Total RNA Isolation System (Promega, Madison, WI, USA) according to the manufacturer’s instructions. The RNA concentrations and purity were measured with an ultraviolet spectrophotometer.
Complementary DNA (cDNA) Synthesis
The cDNA was synthesized from 1 μg RNA using the SuperScript III First-Strand Synthesis System as described in the manufacturer’s protocol (#K1621, Fermentas, Waltham, MA, USA). In brief, 1 μg of total RNA was mixed with 50 μM oligo (dT) 20, 50 ng/μL random primers, and 10 mM dNTP mix in a total volume of 10 μL. The mixture was incubated for 5 minutes at 56°C before being placed in ice for 3 minutes. The reverse transcriptase master mix containing 2 μL of 10× RT buffer, 4 μL of 25 mM MgCl2, 2 μL of .1 M DTT, and 1 μL of SuperScript® III RT (200 U/μL) was added to the mixture and incubated for 10 minutes at 25°C, followed by 50 minutes at 50°C.
Real-Time Quantitative PCR (RT-PCR)
The primer sequence of the studied genes.

Western Blot detection of p-tau protein (V3 Western WorkflowTM Complete System, Bio-Rad® Hercules, CA, USA)
P-tau protein was extracted from bone marrow homogenates using ice-cold radio-immuno-precipitation assay (RIPA) buffer supplemented with phosphatase and protease inhibitors (50 mmol/L sodium vanadate, 0.5 mM phenylmethylsulphonyl fluoride, 2 mg/mL aprotinin, and 0.5 mg/mL leupeptin), then centrifuged at 12,000 r/min for 20 minutes. The phosphor tau protein concentration for each sample was determined according to Bradford 20 (1976). Equal amounts of protein (20-30 μg of total protein) were separated by SDS/polyacrylamide gel electrophoresis (10% acrylamide gel) using a Bio-Rad Mini-Protein II system. The protein was transferred to polyvinylidene difluoride membranes (Pierce, Rockford, IL, USA) with a Bio-Rad Trans-Blot system. After transfer, the membranes were washed with PBS and blocked for 1 h at room temperature with 5% (w/v) skimmed milk powder in PBS. The manufacturer’s instructions were followed for the primary antibody reactions. Following blocking, the blots were developed using antibodies for p-tau and beta actin supplied by Thermoscientific (Rockford, Illinois, USA) and incubated overnight at pH 7.6 at 4°C with gentle shaking. After washing, peroxidase-labeled secondary antibodies were added, and the membranes were incubated at 37°C for 1 h. Band intensity was analyzed by the ChemiDocTM imaging system with Image LabTM software version 5.1 (Bio-Rad Laboratories Inc., Hercules, CA, USA). The obtained results were presented as relative expression after normalization for β-actin protein expression.
Flow Cytometry Assays
TGF-β Expression Cell Surface Markers
One hundred microliters of cell suspension (1 × 106 cell/mL) was prepared by the isolation of mononuclear cells from the processing of bone marrow with Tris-EDTA buffer. The cells were washed with PBS/BSA (bovine serum albumin) with 2 mL and then centrifuged at 2000 r/min for 5 minutes. The supernatant was discarded, and the pellet was resuspended in 100 μL of PBS. Seven microliters of a TGF-β marker [BD, Cat. No. 563143] were thoroughly mixed, and the tube was incubated at room temperature in the dark for 30 minutes. Cells were washed twice with 2 mL of PBS/BSA and centrifuged at 2000 r/min for 5 minutes, with the supernatant discarded. Finally, cells were resuspended in 200 μL of 4% paraformaldehyde in PBS for fixation until acquired by using the BD Flow cytometry Accurie C6 Plus version (Becton Dickinson). Using BD Accuri C6 Plus software, the DNA content of 10 000 cells was analyzed by FACS Caliber flow cytometry to calculate the percentages of TGF-β cell surface markers. Files can be exported in FCS 3.1 format and imported into the flow cytometry analysis program FCS Express™ software. 21 Samples were run in triplicate, and each experiment was repeated three times.
Cell Cycle
One hundred microliters of cell suspension (1 × 106 cell/mL) was prepared from the processing of bone marrow cells with Tris-EDTA buffer. One hundred microliters of cell suspension were added to PI buffer (propidium iodide with RNase) and incubated for at least 1 hour in the dark at +4°C. After incubation, the cells were acquired using the BD Flow cytometry Accurie C6Plus version (Becton Dickinson, San Diego, CA). The DNA content of 10,000 cells was analyzed by FACS Caliber flow cytometry to calculate the percentages of cells occupying the different phases of the cell cycle by using the BD Accuri C6 Plus software files, which can import data into flow cytometry software for seamless software and FCS Express™. 21 Samples were run in triplicate, and each experiment was repeated three times.
Micronucleus Assay
Rat’s bone marrow samples were collected from femur according to Schmid 1976, 22 triplicate slides were prepared for each rat for the micronucleus evaluation. The slides were stained with 5% (v/v) Giemsa stain diluted in phosphate buffer (Na2HPO4 .06 M and KH2PO4 .06 M, pH 6.8). For each animal, 1000 polychromatic erythrocytes (PCEs) were scored under oil immersion using a LeitzWetzlar—Orthomat binocular optical microscope with a magnification of ×1000 to determine the number of micronucleated polychromatic erythrocytes (MNPCEs) and also for normochromatic erythrocytes NCEs. The ratio of PCEs to total erythrocytes (PCEs+NCEs) was evaluated to determine the cytotoxic effects of irradiation in the bone marrow.
Statistical Analysis
The obtained data was expressed as a mean ±SE. Statistical analysis was performed by one-way analysis of variance (ANOVA) followed by LSD multiple comparison tests to make multiple comparisons between all irradiated groups with control and with each other. In addition, tow-way (ANOVA) followed by LSD multiple comparison tests to evaluate the significance of effect of irradiation dose and fractionation on studied parameters. P values <.05 were considered significant. 23
Results
MAPT Gene mRNA Expression in the Bone Marrow
As shown in Figure 1, irradiation resulted in a significant increase in MAPT gene expression in bone marrow as compared with the control group, but there is no significant difference between all irradiated groups. However, two-way ANOVA analysis revealed that the irradiation dose and pattern had no significant effect on this expression in-between irradiated groups. MAPT gene expression in rat bone marrow was reported as relative mean of housekeeping gene (beta actin); (a): significant difference when compared to the control group, there in non-significant difference between all irradiated groups (P < .05).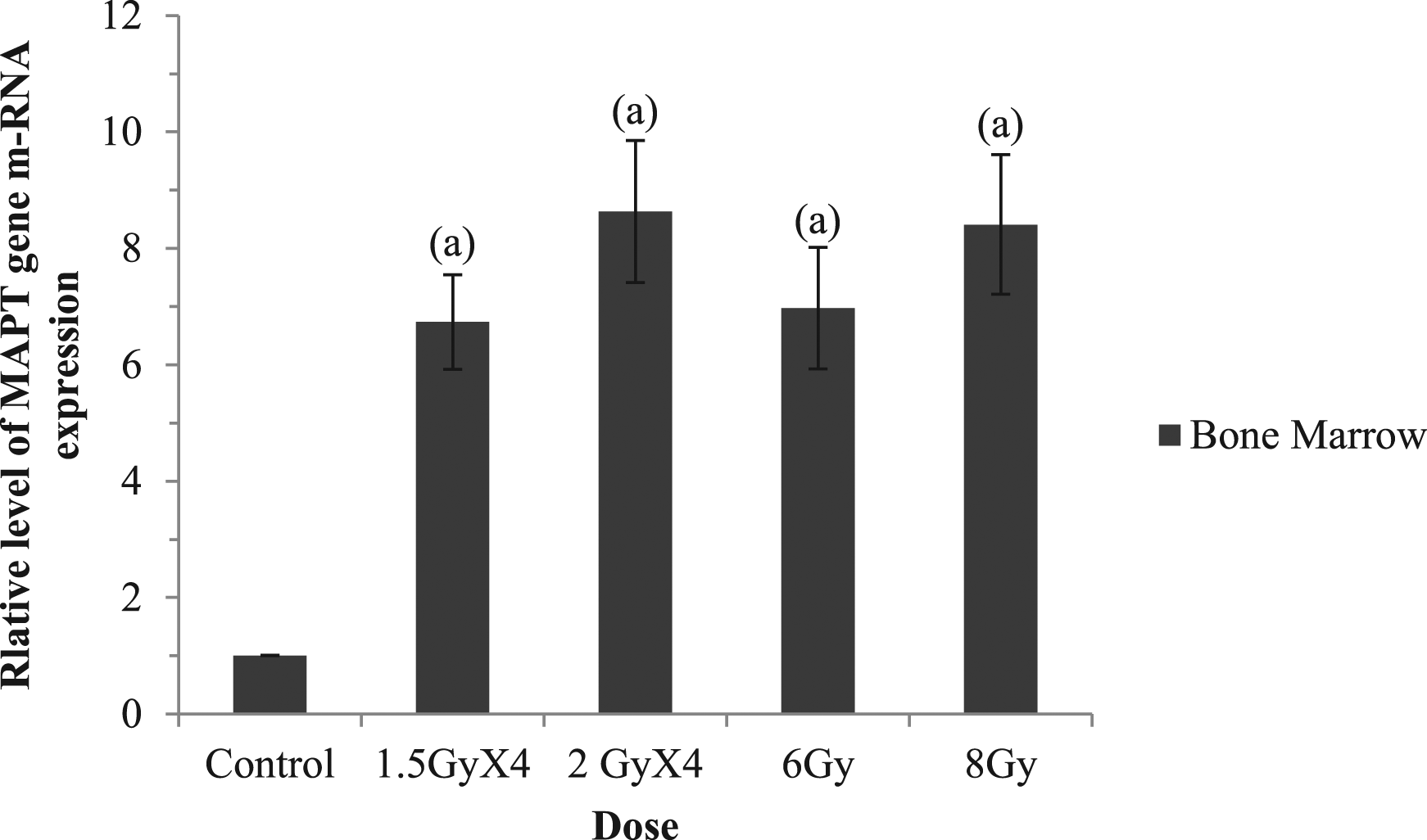
PKN gene mRNA expression in the bone marrow
As illustrated in Figure 2, the relative expression of the PKN gene in the bone marrow shows a non-statistically significant difference between the irradiated groups except for the 8 Gy group which is significantly different from the 6 Gy group. When compared to the control group, irradiation resulted in a non-significant decrease in PKN gene expression in bone marrow except for the 8 Gy group. This reduction is significant when compared with the 6 Gy group. The two-way ANOVA analysis shows that the dose and the pattern of irradiation had no significant effect on the PKN gene expression in the bone marrow cells. PKN expression in rat bone marrow expressed as the relative mean of the housekeeping gene (beta actin); (a): significant difference when compared to the control group; (b): significant difference when compared to the 1.5 GyX4 group; (c): significant difference when compared to the 2 GyX4 group; (d): significant difference when compared to the 6 Gy group, (P < .05).
Western blot analysis of p-tau protein in the bone marrow
As shown in Figures 3 and 4, in the fractionated dose groups (1.5 GyX4 and 2 GyX4), the elevation of p-tau relative density in bone marrow cells was inversely related to the dose. The p-tau relative density was also reported as a reverse relationship with the dose in the acute dose groups (6 Gy and 8 Gy), but this increment was significantly greater than that of the fractionated dose. The two-way ANOVA analysis confirmed the significant effect of pattern and dose of irradiation. Western blot analysis shows the effect of different patterns of irradiation on the p-tau in bone marrow cells. β-actin was used as an internal control. P-tau relative density in rat bone marrow expressed as the relative mean of the housekeeping gene (beta actin); (a): significant difference when compared to the control group, (b): significant difference when compared to the 1.5 GyX4 group, (c): significant difference when compared to the 2 GyX4 group, (d): significant difference when compared to the 6 Gy group, (P < .05).

Flow Cytometric Analysis
TGF-β Cell Surface Markers in the Bone Marrow
As shown in Figures 5 and 6, (a) fractionated dose of radiation (1.5 GyX4) resulted in a significant decrease in TGF-β%, whereas the dose (2 GyX4) resulted in a significant increase in TGF-β% when compared to the control. On the contrary, in the acute dose groups (6 Gy and 8 Gy), the percent of TGF-β decreased in inverse proportion to the radiation dose, becoming significantly lower than in the control. That indicates the significant effect of the pattern and the dose of irradiation, which is declared by the two-way ANOVA analysis. Flow cytometric analysis of TGF-β expressed by the bone marrow cells in rats. (a) charts of control group samples, (b) charts of irradiated group (1.5 GyX4), (c) charts of irradiated group (2 GyX4), (d) charts of irradiated group (6 Gy), E: charts of irradiated group (8 Gy). Where M1 denotes the cell population not stained with the TGF-β antibody, while M2 denotes the cell population stained with the TGF-β antibody. TGF-β (%) in the bone marrow cells of rats. (a) significant difference when compared to the control group, (b) significant difference when compared to the 1.5 GyX4 group, (c) significant difference when compared to the 2 GyX4 group, d significant difference when compared to the 6 Gy group, (P < .05).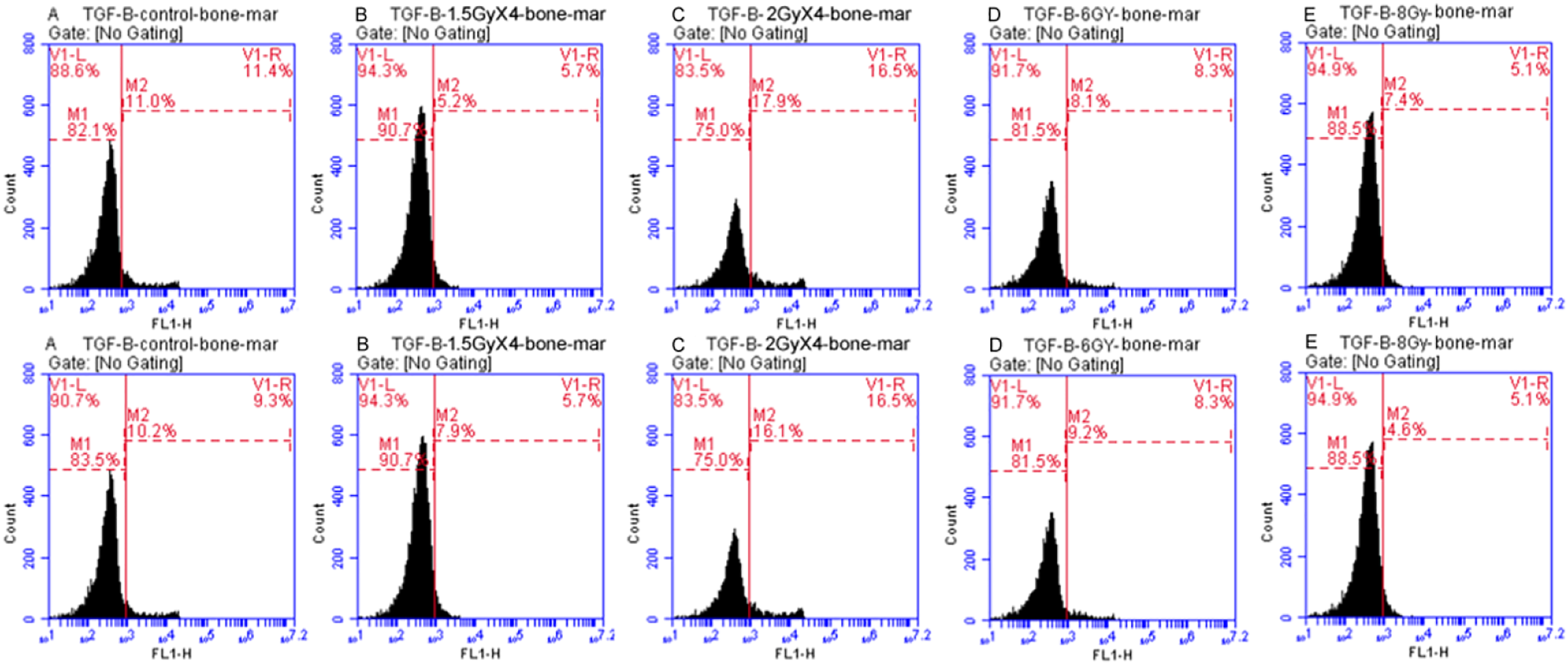

Cell Cycle (Bone marrow)
As shown in Figures 7 and 8, there was a significant decrease in the G0/G1 cells percentage in the group 2 GyX4 when compared to the control group. Results demonstrated that the irradiation 2 GyX4 caused G2/M cell cycle arrest. Moreover, irradiation induces S-phase arrest in the different treated groups; it was more significant in the 2 GyX4 and 8 Gy groups. Analysis by two-way ANOVA showed a significant effect of the pattern and the dose of irradiation on all phases of the cell cycle progression except for the subG1 phase, which is not significantly affected by the dose of irradiation. Cell cycle flow cytometric analysis of rat bone marrow cells of control group and irradiated groups (1.5 GyX4, 2 GyX4, 6 Gy, and 8 Gy) samples. Where the cell cycle phases are (M3: sub G1, M7: G0/G1, M13: S, M14: G2/M). Cell cycle distribution percentage in rat bone marrow of control group and irradiated groups exposed to 1.5 GyX4, 2 GyX4, 6 Gy, and 8 Gy γ-irradiation. Cell cycle phases are expressed as a percentage for each group.

Micronucleus Results
Frequency of micronucleated PCEs, NCEs and ratio of PCE/(PCE+NCE) in bone marrow of rats exposed to various doses of γ-rays; a: significant difference when compared to the control group, b: significant difference when compared to the 1.5 GyX4 group, c: significant difference when compared to the 2 GyX4 group, d: significant difference when compared to the 6 Gy group, (P < .05).

summarizes the significant effect of dose and pattern of γ-radiation on some parameters in rats’ bone marrow exposed to 1.5 GyX4, 2 GyX4, 6 Gy, and 8 Gy as confirmed by two-way ANOVA analysis. (√= significantly affected, x= not significantly affected).

Discussion
Ionizing radiation can cause cell cycle arrest, cell death, or cell mutation. Ionizing radiation’s effects are attributed to its potency as a DNA-injuring agent, which can trigger the flow of cellular reactions and change gene expression profiles. The generation of definite flows of genes is a fundamental feature in the reaction of cells to radiation. These endpoints are definitely crucial in the response to ionizing radiation. 24 The current study focuses on the effects of various patterns and doses of ionizing irradiation on MAPT and PKN gene expression, p-tau relative density, TGF-β% and cell cycle progression, induction of micronuclei, and the proliferation ratio in the bone marrow cells and their role in cell cycle progression. Many studies have been done on MAPT and PKN gene expression, and p-tau concentration in the brain but not in the bone marrow. The results showed that ionizing irradiation at all used patterns increased MAPT gene expression in bone marrow cells with a non-significant effect of dose and pattern. This increase is consistent with the findings of Buratovic et al, 2014, 25 who found that ionizing irradiation increased tau protein levels in male mice. The findings of Cimini et al, 2022, 26 support a role for tau in chromosome stability via its interactions with both microtubules and chromatin. Moreover, Bougé and Parmentier, 2016, 27 have found that an excess expression of human Tau protein induces mitotic arrest, which leads to aneuploidy and cell death. And this is in agreement with the present results of the cell cycle and MN assay.
The study of Herod et al, 2022, 28 pointed out that PKN is cleaved during apoptosis. PKN controls cell migration and gene expression through its kinase activity but does not affect cell proliferation. 29 On the contrary, Misaki et al, 2001, 13 and Al-Sha’er et al, 2022, 30 suggest that PKN plays a significant role in the control of mitotic timing and cell cycle progression. But the present study findings demonstrate a non-significant decrease in the relative gene expression of PKN with the different used patterns and doses of ionizing irradiation except for the 8 Gy dose. These results are in agreement with those of Kovalevska et al, 2022, 29 which pointed out that PKN does not affect cell proliferation. On the other hand, PKN plays a role in regulating tau phosphorylation. 31
The present study results show a significant effect of dose and pattern of ionizing irradiation on the elevation of p-tau in bone marrow cells. Li et al, 2014, 32 reported that ionizing radiation causes changes in tau phosphorylation in cultured cells, which may increase the risk of the development of Alzheimer’s disease. Mitotic tau phosphorylation has been suggested to resemble the hyperphosphorylated tau state seen in Alzheimer’s disease. 33 Tau mis-localization caused by aberrant phosphorylation results in a variety of pathogenic consequences. 34 This alteration in tau, for example, could cause microtubule disruption. 35 Tau’s affinity for microtubules is thought to be reduced when it is phosphorylated. 33
A number of studies have found that chromosomal instability is not only linked to cancer, but may also be a catalyst for it. There are four known direct mechanisms of chromosomal instability: (1) chromosome cohesion abnormalities, (2) spindle assembly checkpoint defects, (3) super-numeral centrosomes, and (4) kinetochore-microtubule dynamics defects. 36 In addition, though tau localization at the mitotic spindle has been observed, nothing is known about its physiological function. Aneuploidy is increased in tau knockout mice, according to research. 33
TGF-β is a multifunctional cytokine that affects cell proliferation, cell death, and immune system responses. It is one of the most prevalent cytokines generated after cells are exposed to ionizing radiation. TGF-β, unlike several other cytokines, has a direct relationship with radiation dose. 37 This is disagreeing with the present study results, which declared that all studied patterns and doses revealed a significant decrease of TGF-β% in bone marrow as compared with the control group except for the group irradiated with 2 GyX4 which showed significant increase in this percent as compared with the control group and all other treated groups. It is known that TGF-β is activated in cells as a result of DNA damage, which is the first genetic consequence of ionizing radiation. Additionally, radiation-induced cell death is important in the release of TGF-β. Radiation can cause cell death by a variety of methods, including apoptosis, mitotic catastrophe, necrosis, senescence, and autophagy. The removal of dying cells that have gone through each of these death stages has various repercussions. Apoptosis clearance by macrophages results in the release of anti-inflammatory cytokines such as TGF-β. 38 According to the mechanisms out-lined, the equilibrium between apoptosis and necrosis plays an important role in TGF-β release. Radiation exposure is another major factor that influences the occurrence of apoptosis and necrosis. Low radiation exposures might increase the apoptosis to necrosis ratio. But this is clear at radiation doses less than 1 Gy. The varying outcomes of different doses and patterns of irradiation may be linked to specific forms of cell death. 39 These findings explain the different TGF-β results in the present study. Moreover, the elevation of TGF-β% in the group irradiated with 2 GyX4 may be attributed to the rupture of MNs after repetitive exposure to the ionizing radiation and the induction of inflammatory response as stated by Fenech et al, 2020. 40 Radiation is a potent inducer of MN for cells exposed in all cell cycle phases. Cell cycle progression and MN formation are required for the synthesis of cytosolic DNA by radiation. The effect of repeated exposure to radiation on MN DNA has not been adequately explored, and it appears to be vital to describe the effect of fractionation in this new setting. 41
It has been known that irradiation of eukaryotic cells slowdown the cell cycle progression. Cell cycle progression is arrested due to DNA damage induced by irradiation, as the cell activates DNA repair mechanisms. 5 If DNA is successfully repaired, that means further progression of the cycle with a delay in the G1, S, and G2 phases of the cell cycle. On the contrary, if the DNA is unsuccessfully repaired, the cell may die and exit from the cycle or continue with genome instability. 42 When compared to the control group, ionizing irradiation increases G2/M, especially at the dose of 2 GyX4, increases S phase at all doses and patterns of radiation, decreases G0/G1, especially at the dose of 2 GyX4, and decreases subG1 at all doses. TGF-β induced a delay in G2/M progression. 43 These findings demonstrate that the increase in G2/M in the 2 GyX4 group was caused by an increase in TGF-β in the same group. When normal cells are exposed to radiation, the G1/S transition is disrupted, stopping further development into the S phase and allowing additional time to repair DNA damage before DNA replication. The arrest is usually temporary, but it might become permanent after exposure to high doses of ionizing radiation. 44 The delay in progression through S phase of the cell cycle is due to slowing of the DNA synthesis rate and DNA damage. The dose response of this effect is biphasic (radio-sensitive/radio-resistant). 45 When double-stranded DNA breaks occur, cells undergo a G2/M arrest, which prevents them from entering the M phase, allowing for coordinated repair of the damage. The recovery process can be prolonged in severe cases of radiation, and irreversible DNA damage can sometimes lead to mitotic catastrophes, which result in cell death. 44
On the other hand, the frequency of bone marrow micronucleated PCEs and NCEs showed increase and ratio of PCE/(PCE+NCE) showed decrease in all irradiated groups in a dose dependent manner. Reduced ratio of PCE/(PCE+NCE) may be attributed to the cell death. This ratio is a measure of the rate of proliferation. A decrease in the post-irradiation ratio shows induced bone marrow cytotoxicity or erythropoiesis suppression, which is an indication of the known early effects of radiation on the cell cycle.
4
According to Fenech et al, 2011,
46
hypo-methylation of centromeric and paracentromeric regions (satellite) is the fundamental mechanism of MN formation resulting from chromosome mal-segregation. Tubulin de-polymerization prevents the mitotic spindle from pulling chromatids/chromosomes apart, preventing them from segregating. It is essential to note that MNs containing entire chromatids/chromosomes form as a result of spindle failure.
47
In accordance with the previously mentioned results of MAPT gene and p-tau in the present study and their relations with the microtubules and the mechanism of MN formation, the obtained results of the MN assay are clear. Figure 9 summarizes the suggested effect of different patterns and doses of γ-radiation on the studied parameters and their consequent effects on the cell cycle from the obtained results, pointing out the role of the genetic control on the cell cycle progression. Schematic diagram of the suggested effect of different patterns and doses of γ-radiation on the studied parameters and their consequent effects on the cell cycle, pointing out the role of the genetic control on the cell cycle progression. (Created in BioRender.com).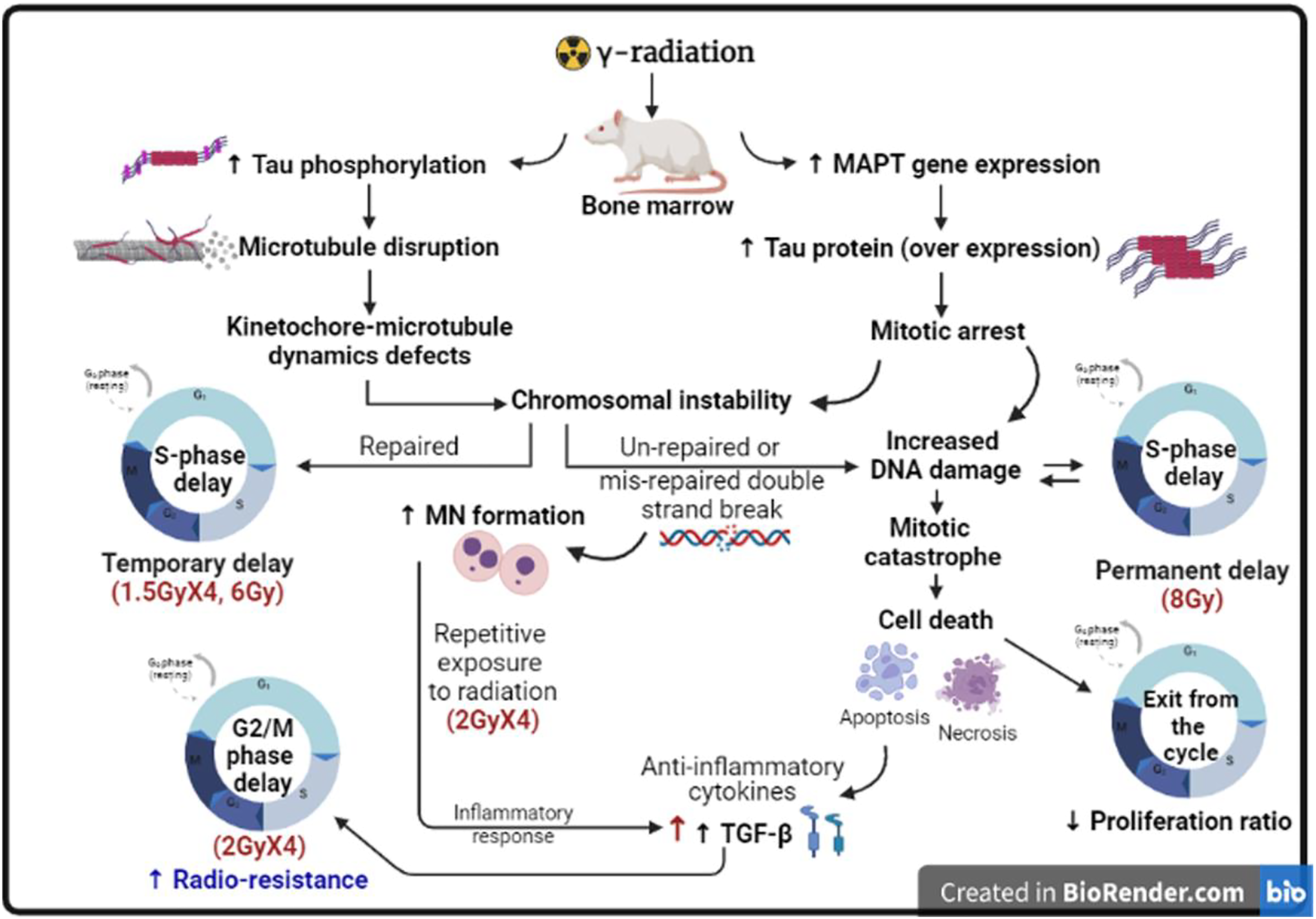
This study did have some limitations. The first issue was the lack of previous studies on the effect of ionizing radiation on these factors (MAPT, PKN gene expression, p-tau, and TGF-β) in bone marrow. Another limitation was the sample volume (the more factors to be studied, the more samples that were needed). The MN assay yielded the most relevant results for humans because cytogenetic analysis has been considered a comprehensive and broad measurement responsive to different γ-irradiation doses in both humans and animals, but TGF-β could be affected by the body’s inflammatory status. Finally, there are definitely still numerous information gaps that need to be filled, and future mechanism research is needed to clarify the mechanism by which DNA damage (micronucleus), tau phosphorylation, TGF-β and cell cycle are linked.
Conclusion
From this study, it could be concluded that the irradiation dose and patterns had a significant effect on increasing tau phosphorylation, TGF-β% in different fashions, all phases of the cell cycle except for the subG1 phase, MN frequency, and cell proliferation reduction but not on the elevation of MAPT gene expression and PKN gene expression. The fractionated dose 2 GyX2 has a distinct effect on TGF-β elevation and G2/M phase delay, which may be attributed to repeated exposure of formed MNs, which when ruptured induce inflammatory reactions and lead to production of the TGF-β as an anti-inflammatory cytokine. Increased DNA damage causes a biphasic delay in S-phase advancement, showing the role of a genetic regulatory mechanism in determining the extent of S-phase delay.
Footnotes
Acknowledgments
We would like to express our special gratitude to our colleagues and members of NCRRT, EAEA for providing the necessary irradiation facilities.
Author Note
The authors alone are responsible for the content and writing of the paper.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.