
Abstract
Previous studies evaluated the adenosine receptor antagonists alone to determine their effects on oxidative stress, but little is known about adenosine’s protective efficacy when oxidative injury occurs in vivo. Adenosine is a crucial signaling molecule recognized by four distinct G-protein-coupled receptors (GPCRs) (i.e., A1R, A2AR, A2BR, and A3R) and protects cells against pathological conditions. The present study was performed to evaluate the role of antagonist modulation in the setting of paraquat toxicity with adenosine pretreatment. First, PC12 cells were exposed to paraquat (850 μM) and adenosine (30 μM) to develop an in vitro model for the antagonist effect assay. Second, we found that the A1R antagonist DPCPX enhanced the viability of paraquat-induced PC12 cells that underwent adenosine pretreatment. Moreover, the A2AR antagonist ZM241385 decreased the viability of paraquat-induced PC12 cells that underwent adenosine pretreatment. Our findings indicate that adenosine protection requires a dual blockade of A1R and activation of A2AR to work at its full potential, and the A2B and A3 adenosine receptor antagonists increased paraquat-induced oxidative damage. This represents a novel pharmacological strategy based on A1/A2A interactions and can assist in clarifying the role played by AR antagonists in the treatment of neurodegenerative diseases.
Introduction
Oxidative stress is one factor that plays a potential role in the pathogenesis of neurodegenerative disorders, including Alzheimer’s disease (AD), Parkinson’s disease (PD), and other neurodegenerative diseases.1,2 The herbicide paraquat (PQ, N,N′-dimethyl-4,4′-bipyridinium dichloride) undergoes a nicotinamide adenine dinucleotide phosphate (NADPH)-dependent reduction in vivo, yielding a stable paraquat radical that reacts with oxygen to generate a superoxide anion, which is a reactive oxygen species (ROS) that causes immediate damage by direct contact, ingestion, or inhalation. 3 Therefore, paraquat is a satisfactory chemical substance for generating an oxidative stress model using cells or animals in the laboratory.4,5
Adenosine (ADO) is an omnipresent and rapidly metabolized purine nucleoside with numerous physiological actions in the cardiovascular, nervous, renal, pulmonary, and immune systems. 6 It is well documented that ADO is a metabolic byproduct of adenosine triphosphate (ATP) breakdown from precursor adenosine monophosphate (AMP), 7 and it is upregulated during hypoxic conditions resulting from an imbalance between tissue oxygen supply and demand.8,9 Physiological concentrations of extracellular ADO have been reported to be in the low nanomolar range, but under pathological conditions, they can be as high as 100 mM.10,11 A large amount of evidence from previous studies has suggested that the effects of ADO-associated drugs can be used for the treatment of pathological conditions, such as oxidant injury in neural cells, 12 anxiety disorder, schizophrenia, epilepsy, and drug addiction. 13
It has become apparent that the physiological and pathological effects of ADO occur because it is a crucial signaling molecule mediated by four distinct G-protein-coupled receptors (GPCRs), namely A1R, A2AR, A2BR, and A3R. 14 Four adenosine receptors (ARs) are endogenously activated by extracellular ADO and mediate its protective function in response to stress stimuli, tissue damage, or inflammation.
There is growing evidence that the ARs may be a promising therapeutic target under many conditions. For example, treatment with AR antagonists, such as the selective A1R antagonist DPCPX, suppressed tumor progression in hepatocellular carcinoma, 15 and attenuated the injury of neuroblastoma cells induced by aluminum chloride. 16 The A2AR-specific antagonist ZM241385 abrogated the effect of inosine in checkpoint blockade immunotherapy. 17 The A2BR-specific antagonist MRS1754 suppressed renal cell carcinoma (RCC) cell proliferation and migration. 18 Experiments showed that the other A3R antagonist VUF5574 was responsible for neuroprotective and anti-inflammatory effects and was suitable for potential treatment of glaucoma and asthma. 19
Most previous studies have examined the effects of ADO, whereas the role of AR antagonists has been evaluated alone, and there is a lack of data on what transpires in real-world environments where adenosine exerts its protective effects in response to injury. In this study, we applied PC12 (rat adrenal pheochromocytoma) cells to imitate neuronal cells and test whether antagonists might adjust the PC12 cell injury induced by paraquat in the presence or absence of adenosine pretreatment.
Materials and Methods
Culture of PC12 Cells
PC12 cells (purchased from the Cell Bank of the Chinese Academy of Sciences) were grown in Gibco’s Dulbecco’s modified Eagle’s medium (DMEM) with high glucose and supplemented with 15% heat-inactivated horse serum (Sangon Biotech, Shanghai, China), 2.5% heat-inactivated fetal bovine serum (FBS, Boxun Biotech, Shanghai, China), and 1% penicillin/streptomycin (Sangon Biotech, Shanghai, China) at 37°C in an incubator (Thermo Forma 311, MA, USA) with a humidified atmosphere of 95% air and 5% CO2. The medium was changed every other day. Cells were collected by digestion with TrypLE™ Express (Thermo Fisher Scientific, MA, USA) in their logarithmic growth phase. Prior to the following experiments, PC12 cells were centrifuged at 250 g for 5 min to remove the liquid medium and endogenous adenosine.
Chemicals and Reagents
Paraquat solution
Paraquat was supplied by Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). A total of .2559 g paraquat was dissolved in 10 mL sterilized distilled water in a tube to produce a stock solution with a theoretical final drug concentration of .1 M. The solution was then transferred into Eppendorf tubes and stored at −20°C for the following paraquat poisoning assay.
Adenosine Solution
Adenosine (purity ≥99%) was supplied by Sigma Aldrich (Merck KGaA, Darmstadt, Germany). A total of .8017 g adenosine was dissolved in 10 mL sterilized distilled water in a tube to produce a stock solution with a theoretical final drug concentration of .3 M. The solution was then stored at −20°C in Eppendorf tubes for the following adenosine protection assay.
AR Antagonist Solution
Four different ligands for specific ARs were tested in this study, including a selective adenosine A1 receptor (A1R)-specific antagonist [8-cyclopentyl-1,3-dipropyl xanthine] DPCPX (Sigma, USA, named DPCPX A1R below), a selective adenosine A2A receptor (A2AR)-specific antagonist [4-(2-[7-amino-2-(2-furyl)[1,2,4]-triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)-phenol] ZM241385 (Sigma, USA, named ZM A2AR below), a selective adenosine A2B receptor (A2BR)-specific antagonist [N-(4-cyanophenyl)-2-[4-(2,3,6,7-tetrahydro-2,6-dioxo-1,3-dipropyl-1H-purin-8-yl)- phenoxy]acetamide] MRS1754 (Sigma, USA, named MRS A2BR below), and a selective adenosine A3 receptor (A3R)-specific antagonist [N-(2-methoxyphenyl)-N′ -[2-(3-pyridinyl)-4-quinazolinyl]-urea] VUF5574 (Sigma, USA, named VUF A3R below). All reagents were dissolved in 25% dimethylsulfoxide (DMSO) solution. All other chemicals and solvents were from standard commercial sources and were of analytical or pharmaceutical grade.
Development of the Paraquat-Adenosine-PC12 Cell Model
Paraquat poisoning assay
Dosing solutions were prepared by serially diluting the stock solution with sterile phosphate-buffered saline (PBS, pH 7.4) to obtain the following different concentrations: 200 μM, 400 μM, 600 μM, 800 μM, and 1000 μM; sterile PBS buffer (pH 7.4) only was considered as the blank. The viabilities of PC12 cells treated with paraquat for 24 h were assessed by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) colorimetric assay.
Adenosine Protection Assay
An 850-μM paraquat treatment dose based on the LD50 of the above paraquat-poisoning assay was selected to determine the concentration that offered the strongest protection for the adenosine-pretreated cells. Prior to paraquat poisoning, the adenosine solution was diluted with sterile PBS (pH 7.4) to obtain the concentrations of 3 μM, 15 μM, 30 μM, 300 μM, and 3000 μM, which were used to pretreat PC12 cells for 24 h. Sole treatment with sterile PBS buffer (pH 7.4) was considered to be 0 μM. Viabilities of PC12 cells were also assessed by MTT colorimetric assay.
AR Antagonist Treatment and Grouping
To assess the chemical effect of the adenosine antagonists, PC12 cells were individually treated with four AR antagonists: DPCPX, ZM241385, MRS1754, and VUF5574. The DPCPX A1R group, ZM A2AR group, MRS A2BR group, and VUF A3R group were pretreated with DPCPX (10 μM), ZM241385 (1 μM), MRS1754 (10 μM), and VUF5574 (10 μM) for 24 h, respectively, and were cultured at 37°C in an incubator in 5% CO2. The concentration of each antagonist along with their molarity was based on previous studies.20,21 The ADO (adenosine-treated) group was pretreated with 30 μM adenosine (the protective concentration obtained in the above adenosine protection assay) for 24 h and was cultured in an incubator in the same manner as that which was used for the antagonist groups. The PQ group was treated with 850 μM paraquat for 24 h, and the same volume of PBS was used as that for the blank control group.
To assess the modulation effect of the four AR antagonists on the corresponding receptors including A1R, A2AR, A2BR, and A3R of PC12 cells, the viabilities of the DPCPX A1R group, ZM A2AR group, MRS A2BR group, and VUF A3R group cells in the presence or absence of adenosine pretreatment were measured by exposure to paraquat. The ADO, DPCPX A1R , ZM A2AR , MRS A2BR , and VUF A3R group cells were pretreated with 30 μM adenosine for 24 h. Subsequently, the ADO, DPCPX A1R , ZM A2AR , MRS A2BR , VUF A3R , and PQ group cells were co-cultured with 850 μM paraquat for another 24 h. After a total of 48 h, the viabilities of PC12 cells were assessed by MTT colorimetric assay.
Staining of Apoptotic Cells and Cell Viability Assay
Cells exposed to paraquat typically exhibit nuclear condensation or reduced nuclear size. The characteristic features of apoptotic nuclei among adenosine, paraquat, and adenosine+paraquat treatments were assessed using Hoechst 33 342 fluorescent dye. Briefly, PC12 cells were fixed with 4% paraformaldehyde and then stained with 5 mg/mL of Hoechst 33 342 for 10 min. Hoechst 33 342 was excited using 351 nm UV laser light, and its emission was captured with a bandpass filter set at 450 ± 20 nm. Nuclear morphology was visualized under a fluorescence microscope (Leica, Allendale, NJ, USA).
An MTT cell proliferation and cytotoxicity assay kit (BBI, Crumlin, UK) was used to assess the PC12 cell viability according to the recommended protocols.
22
Briefly, 5 × 103 cells/well density of PC12 cells were incubated in a 96-well plate overnight, and the assay trials were carried out according to the above-described methods the following day. To obtain robust calibration data, at least six replicates of each treatment were set up. Medium-only as zero-adjustment was used for standardized data, and PBS was added to the plate wells surrounding the treatment wells to maintain moisture. All the culture medium was aspirated from each well after treatment for 24 h, and MTT solution dissolved in fresh medium was added according to the manufacturer’s protocol. After the solution was incubated at 37°C for 4 h, the supernatant was carefully removed. Subsequently, 100 μL of DMSO was added to the wells for precipitation, and the plates underwent shaking for 10 min. The liquid in the 96-well microplates was analyzed under the absorbance of 570 nm using a multifunctional microplate reader (SpectraMax® M3, Molecular Devices, CA, USA). The original absorbance value was subtracted from the absorbance value of the zero-adjusted well to obtain the final result, and the cell viability is expressed as a percentage of the control group. The equation below was used to calculate the PC12 cell survival

Statistical Analysis
A dose-mortality modeling technique, yielding the parameters for dose effects of the paraquat-poisoned PC12 cells, was performed as previously described. 23 The procedures, including the test for goodness of fit and estimation of virulence indices (LD50) using the parameters, were conducted using the maximum likelihood method. Comparison and analysis of the protective effects of various concentrations of adenosine pretreatment on paraquat-poisoned PC12 cells, comparison and analysis of the protective effects of adenosine pretreatment with four antagonists, comparison and analysis of the modulation effect of the antagonists on A1R, A2AR, A2BR, and A3R, and the survival of PC12 cells were analyzed by one-way analysis of variance (ANOVA) and post hoc multiple comparisons using Tukey’s test.
Multinomial logistic regression (MLR) analysis was performed to determine the underlying effects on ARs. MLR predictive analysis was conducted for the adenosine antagonists. All data are presented as the mean ± standard deviation. P < .05 was considered to indicate a statistically significant difference. All analyses were conducted using the latest version of Data Processing System (DPS) software. 24
Results
Paraquat Poisoning Concentration Evaluation
To evaluate the survival responses to elevated oxidative stress, we first administered various doses of paraquat to PC12 cells for 24 h. In Figure 1, the injury of PC12 cells by paraquat is illustrated by an S-shaped curve over increasing concentration, which indicates that the higher the concentration, the more severe the injury sustained by cells. Treated with 200, 400, 600, 800, or 1000 μM of paraquat, the mortality of cells after 24 h was 23%, 23.6%, 30.3%, 41.6%, and 79%, respectively. The curve equation Y = 1/[−.07 + 1.75 × EXP(−X)] was well fitted with the relationship between paraquat concentration and cell mortality (P < .0001, R2 = .89). In addition, the mortality of PC12 cells treated with different paraquat doses in the poisoning test was corrected by the natural mortality rate of the control, and a probit analysis was performed. The LD50 (95% CI) of paraquat treatment on PC12 cells was calculated to be 813 (612–1079) μM. Therefore, 850 μM was selected as the optimal poison dose of paraquat for subsequent experiments. Paraquat-poisoning effect on PC12 cells. After 24 h of incubation with paraquat (PQ) at increasing doses (200, 400, 800, and 1000 μM) demonstrated as Log-transformation, cell viability was ascertained by MTT colorimetric assay. The fitted S-shaped curve over increasing concentration was illustrated as Y = 1/[−0.07 + 1.75 × EXP(−X)] (R2 = 0.89).
Protective Effect of Adenosine Against Paraquat-Induced Cytotoxicity
The MTT test of cell viability showed that adenosine attenuated the cytotoxic effect of paraquat in a dose-dependent manner up to 30 μM (Figure 2). Treatment of PC12 cells with 3-30 μM adenosine led to a significant increase in cell viability to 122.74% ± 10.68% (P < .01). However, when the concentration exceeded 300 μM, the protective effect of adenosine pretreatment on cells began to decrease. When the concentration reached 3000 μM, there was no longer any protective effect by adenosine pretreatment on cells, which resulted in aggravated cell injury. Therefore, 30 μM was selected as the optimal adenosine protective concentration for subsequent PC12 cell experiments. Effect of adenosine on paraquat-induced apoptosis as measured by MTT. The number of PC12 cells pretreated with increasing concentrations of adenosine was counted as viable cells. ** p < 0.01 for = comparison between the treated group and the control group. “–” and “+” indicate “in the absence of” and “in the presence of,” respectively.
In addition, we performed Hoechst 33 342 staining to observe the nuclear morphological changes associated with apoptosis. In Figure 3, compared with the non-treated group (negative control, NC), PC12 cells treated with ADO as a positive control exhibited no characteristic features of apoptotic nuclei. Inversely, PC12 cells treated with 850 μM PQ alone exhibited typical characteristics of apoptosis, such as highly condensed nuclei and decreased numbers of cells, when observed under a fluorescence microscope. However, 30 μM adenosine prevented these manifestations during exposure to paraquat, resulting in greater numbers of more round and homogeneously stained nuclei, similar to those observed in the control cells. Effect of adenosine on paraquat-induced nuclear condensation. PC12 cells were treated with paraquat (PQ, 850 μM) for 24 h in the presence or absence of adenosine (ADO, 30 μM), PBS as a negative control (NC), and adenosine as a positive control. The cells were observed by fluorescence microscopy after the nuclei were stained with Hoechst 33342. Arrows indicate nuclear condensation.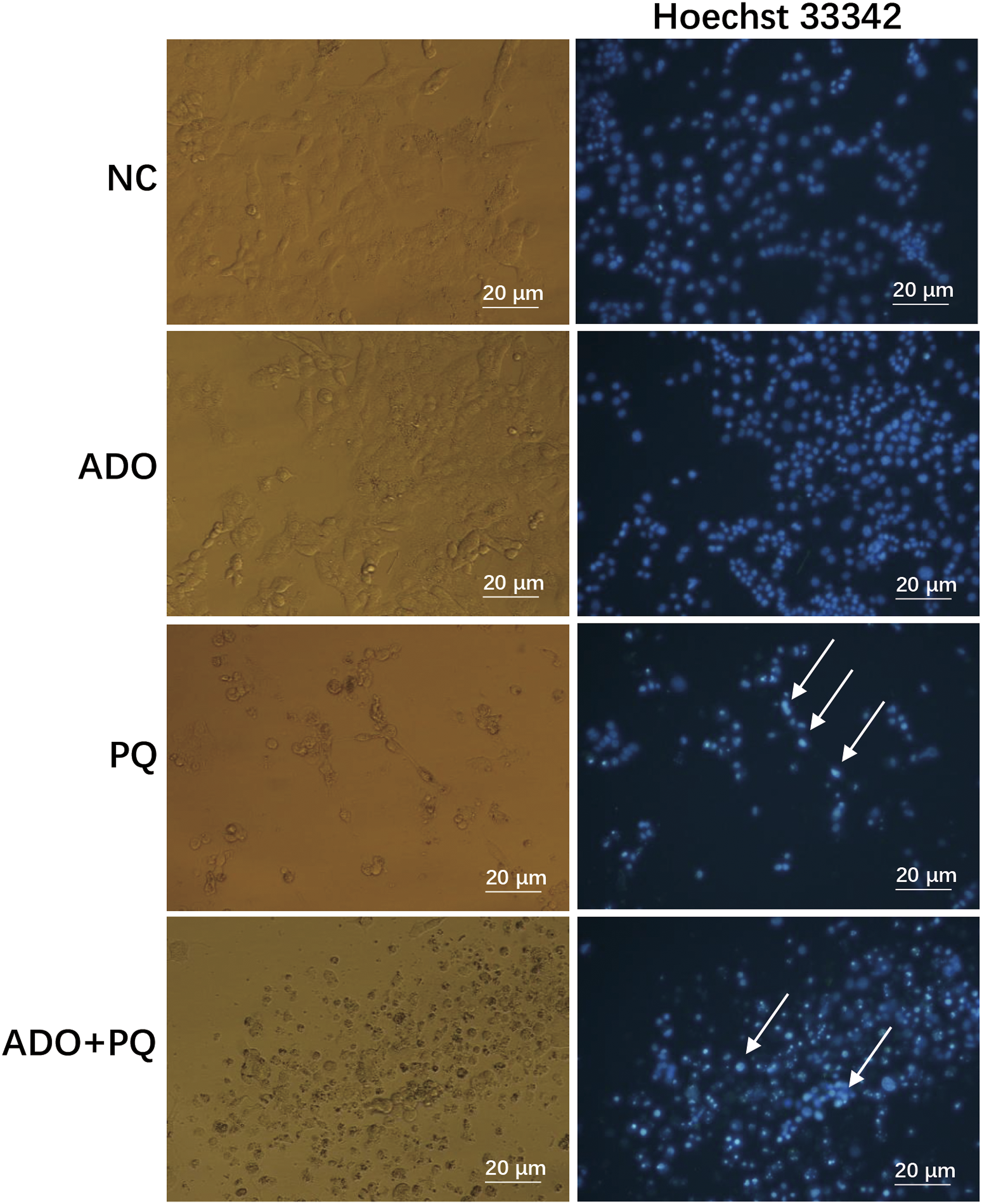
Adenosine Receptor Antagonist Treatment
The protective effects of four adenosine antagonists (DPCPX, ZM241385, MRS1754, and VUF5574) are demonstrated in Figure 4A. A significant difference was observed when comparing the ADO group and the four antagonist groups to the PQ group (P < .01). This indicates that the four antagonists conferred protective effects on paraquat-induced PC12 cells. However, there were significant differences between the MRS
A2BR
and VUF
A3R
groups and the ADO group (P < .05), which may reflect different modulatory effects of the corresponding ARs (A1R, A2AR, A2BR, and A3R) in the presence of paraquat. Protective effect of antagonists on ARs as measured by MTT. The protective effects of the four AR antagonists, DPCPX, ZM241385, MRS1754, and VUF5574, are demonstrated in panel A. Panel B shows that there are significant differences between the DPCPXA1R and ZMA2AR groups in the presence or absence of adenosine. There were no significant differences between the MRSA2BR and VUFA3R groups. The numbers of viable cells were normalized by the control (PQ group pretreated with and without adenosine). Each value represents the mean ± SD relative to the control from six independent experiments. * and ** indicate p < 0.05 and p < 0.01, respectively, compared with the control group. “¬–” and “+” in the figure indicate “in the absence of” and “in the presence of,” respectively.
Using the conditions of the established adenosine-paraquat-PC12 cell model, the modulatory effect of the four antagonists is shown in Figure 4B. All data were normalized by the controls (PQ group pretreated with and without adenosine). There was a significant increase in cell viability in the DPCPX A1R + ADO group as compared to the DPCPX group in the presence of paraquat (P < .05). The cell viabilities in the DPCPX A1R + ADO group and DPCPX A1R group were 96.82% ± 12.73% and 124.67% ± 22.44%, respectively. Additionally, there was a significant decrease in cell viability in the ZM A2AR + ADO group as compared to the ZM A2AR group in the presence of paraquat (P < .01). The normalized cell viabilities of the ZM A2AR + ADO group and ZM A2AR group were 102.34% ± 6.28% and 90.74%± 5.61%, respectively. There was no significant difference between the other two antagonist groups and the normalized control.
Multinomial logistic regression analysis of AR antagonists under the adenosine-paraquat-PC12 cell model
The categorical variable assignment was as follows: DPCPX A1R -treated = 1, else = 0; ZM A2AR -treated = 1, else = 0; MRS A2BR -treated = 1, else = 0; VUF A3R -treated = 1, else = 0. Antagonists were chosen as the dependent variables: DPCPX A1R as X1, ZM A2AR as X2, MRS A2BR as X3, and VUF A3R as X4. Paraquat treatment-protected in the absence of adenosine (Pf) and paraquat treatment in the presence of adenosine [(P + A)f] were used as independent variables. The results showed that the logistic regression model was fitted very well, because the three P values of likelihood ratio chi-square, Pearson chi-square, and Freeman–Tukey chi-square were .99.
A logistic model for further predictive analysis was obtained according to the parameter estimates of the MLR model for paraquat treatment in the presence of adenosine [(P + A)f] vs paraquat treatment-protected in the absence of adenosine (Pf) that was as follows: (P + A)f/Pf = EXP(.08 + .17X1 − 0.2X2 − 0.1X3 − .13X4). The parameter estimates for the model were both significant (P < .05). The estimated value of the DPCPX A1R parameter was .17, while the estimated value of the control (adenosine) parameter was .08. This suggests that the adenosine antagonist DPCPX A1R enforces its protective effect in the presence of adenosine. Alternatively, A1R might play an important role in weakening the protection of adenosine. However, the estimated values of the other three antagonists were all negative, which suggested that they have a repressing effect on the protection of adenosine. Alternatively, A2AR, A2BR, and A3R might play an important role in strengthening the protection of adenosine.
Comparing the presence of adenosine with the absence of adenosine, the results of partial correlation analysis for the predictive frequency of antagonists are shown in Figure 5. There are different predictive frequencies for various antagonists. In the presence of adenosine, DPCPX
A1R
, whose frequency is 56.29%, has a higher protection predictive frequency than the other three antagonists, and even the adenosine control. This suggests that the protection of adenosine would be enhanced through inhibition by the A1 adenosine receptor antagonist DPCPX. ZM
A2AR
has the lowest predictive frequency among ARs, whose frequency is 47%. This suggests that the protection of adenosine would be suppressed through inhibition by the A2A adenosine receptor antagonist ZM241385. As for MRS
A2BR
and VUF
A3R
, their predictive frequencies are 49.6% and 48.72%, respectively, both of which are lower than that of the adenosine control, which is 51.98%. Partial correlation analysis for the predictive frequency of antagonists in the presence or absence of adenosine after paraquat exposure. (P + A)f denotes paraquat treatment in the presence of adenosine, and Pf denotes paraquat treatment-protected in the absence of adenosine. DPCPXA1R, ZMA2AR, MRSA2BR, VUFA3R, and the control represent the DPCPX, ZM241385, MRS1754, and VUF5574 treatments and adenosine treatment, respectively.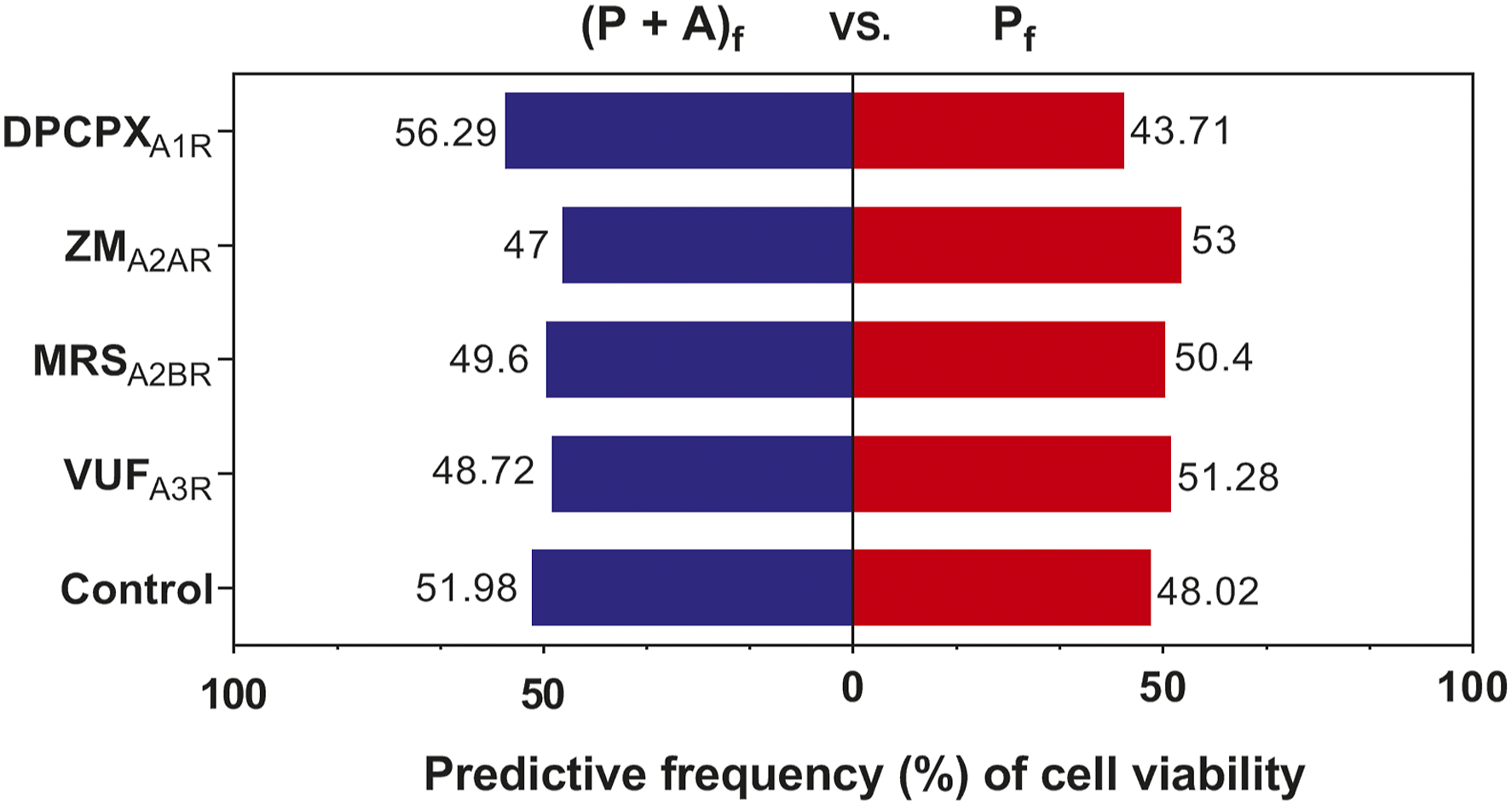
Discussion
There are many conflicting effects for AR antagonists during various in vivo and in vitro models correlated with oxidative stress. The main reasons for such controversies are that the increasing concentrations of adenosine that occur along with oxidative stress have been ignored. Additionally, because the physiological half-life of adenosine is a few seconds, it is difficult to measure the concentrations of circulating adenosine in vivo. 25 Furthermore, there are different dissociation equilibrium constants for saturation binding, affinity, or K D values with the antagonists during various conditions for the four ARs.
It is therefore of importance to characterize the mechanisms of protection that could be useful for future therapies. With this in mind, the present study was designed to evaluate the role of antagonist protection in the setting of paraquat toxicity with adenosine pretreatment. To accomplish these goals, we conducted two distinct sets of experiments using PC12 cells exposed to paraquat (850 μM) and PC12 cells pretreated with adenosine (30 μM) as an in vitro model. In addition, we also provided evidence that antagonists for A1R prevented paraquat damage in adenosine-pretreated PC12 cells, while the results for antagonists for A2AR, A2BR, and A3R were the opposite.
First, the optimal concentration of paraquat was determined using a dose-mortality model with PC12 cells. The PC12 cell line is derived from rat adrenal medulla pheochromocytoma cells, which exhibit typical neuronal characteristics and are often used in neurotoxic damage research and the development of drugs for neurodegenerative diseases. 26 Paraquat is a potent oxidizing herbicide that induces cytotoxicity or cell death in vitro and is a promising candidate for the study of neurodegenerative diseases.27,28
PC12 cells have been reported to have different LD50 values for paraquat.29,30 In the present study, 850 μM was chosen as the treatment dose based on the LD50 for the following adenosine-paraquat model. The adenosine protection results showed that exogenous adenosine protected PC12 cells damaged by oxidative stress. Similar studies documented that adenosine released in large quantities triggered a cytoprotective response in heart injury, such as ischemia or hypoxia, 31 or in the brain affected by neurodegenerative diseases. 32 The strongest protective effect for paraquat-induced PC12 cells was obtained with 30 μM adenosine.
The protective effect of adenosine on PC12 cells began to decline when concentrations were higher than 30 μM. The lowest cell viability was attained at the highest concentration (3000 μM). It is well known that adenosine can undergo extracellular to intracellular transport via equilibrative nucleoside transporters (ENTs) on the cell membrane, as well as by binding to ARs. 33 Adenosine entering the cell is converted to adenosine monophosphate (AMP) by adenosine kinase, and this process initiates the production of AMP-activated protein kinase (AMPK), which regulates ATP levels for energy homeostasis 34 but also leads to different types of apoptosis.35,36 Therefore, a reasonable explanation for such a discrepancy is likely related to high concentrations of exogenous adenosine that decreased the survival rate of PC12 cells via the apoptosis caused by the ENT-AMPK pathway.
In the second part of this work, we aimed to explore the antagonists’ modulatory effect on PC12 cells by exposure to either paraquat alone or in combination with adenosine pretreatment. The concentration of each antagonist along with their molarity was selected based on previous studies. A similar protective effect was observed upon administration of adenosine A1 receptor antagonist (DPCPX) with adenosine to cells exposed to paraquat alone (Figure 4A). However, there was a significant increase in cell viability in the DPCPX + ADO group as compared to that in the DPCPX group (Figure 4B). This suggested that the protection of adenosine would be enhanced through inhibiting the A1 adenosine receptor by DPCPX. A1R might be a negative receptor for adenosine protection, which is consistent with previous studies that reported that DPCPX reversed the injury of PC12 cells induced by adenosine acting on A1R. 37 Additionally, it was reported that the activation of A1R resulted in an observable anti-proliferative and pro-apoptotic effect on cancer stem cells, which could lower the recurrence of glioblastoma multiforme. 38
A similar protective effect was observed for the A2AR antagonist ZM241385 with adenosine when PC12 cells were exposed to paraquat alone (Figure 4A). However, there was a significant decrease in cell viability in the ZM A2AR + ADO group as compared to that of the ZM A2AR group (Figure 4B), which suggested that ZM241385 dramatically inhibited the binding between adenosine and A2AR. Alternatively, A2AR may play an essential role in protection. In previous studies, it was found that stimulation of A2AR prevented apoptosis in PC12 cells induced by hypoxia 39 or in PC12 cells undergoing serum deprivation.40,41 It has been reported that the A1R and A2BR subtypes play a pivotal role in the survival and/or differentiation of glioblastoma cancer stem cells (CSCs). 38 However, our results showed that A2BR and A3R conferred no significant effects on paraquat-induced PC12 cells in the presence or absence of adenosine pretreatment.
Specifically, we analyzed the predictive frequency of the four antagonists on the protection of PC12 cells. A partial correlation analysis showed that the highest protection predictive frequency was calculated for the DPCPX treatment group, while the lowest was for the ZM241385 treatment group. This was consistent with our results, as demonstrated in Figure 4; that is, A1R was disadvantageous and A2AR was beneficial for protection of paraquat-exposed PC12 cells by adenosine.
The reason why different protective effects on PC12 cells were observed for the four antagonists might be due to their ARs’ affinity for adenosine. A2AR and A1R are high-affinity ARs, while A2BR and A3R are low-affinity ARs that require higher concentrations of adenosine for activation.42,43 Additionally, adenosine might be involved in cellular energy transfer and plays important roles in various signal transduction pathways as a component for the formation of signaling molecules such as cyclic adenosine monophosphate (cAMP). 6
When mitogen-activated protein kinases (MAPKs) are activated, they can phosphorylate their specific cascade proteins and therefore control many cellular activities, including cell proliferation, differentiation, and cell death.44,45 Therefore, the underlying signaling pathways that are involved in the ERK, p38, and JNK MAPK pathways should be determined so that the protective effects of adenosine in response to paraquat-induced PC12 cell injury can be elucidated. Furthermore, signal transduction in the coupling of ARs to Gi and Gs proteins might be important when interpreting previous results, for example, a previous report revealed that the A1R-A2AR heterotetramer may constitute a functional unit for signal transduction in HEK-293T cells. 46
Conclusion
These results demonstrate that adenosine protection through the activation of the intracellular antioxidant mechanism requires dual blockade of A1R and activation of A2AR to work at its full potential. They also showed that A2B and A3 AR antagonists increase paraquat-induced oxidative damage. Furthermore, our data provide a novel pharmacological strategy based on A1/A2A interactions and can assist in clarifying the role played by AR antagonists in the treatment of neurodegenerative diseases.
Footnotes
Acknowledgments
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The authors would like to thank the Natural Science Foundation of China (31461143030), National Key Research and Development Plan (2018YFA0108403) and the Zhejiang Natural Science Foundation (Y18C140002, LGN18C200026, 21SBYB08) for supporting this project.