
Abstract
Background and objective
This study was designed to compare the efficacy of curcumin (CRN) with that of nano-curcumin (N-CRN) in the mitigation of various biochemical indices in hypoxic lung induced by sodium nitrite (SN) in rats.
Methods
Twenty-four adult male albino rats were divided into 4 groups. Group 1: control group received carboxy methyl cellulose; Group 2: hypoxic group injected with single dose of SN (60 mg/kg, s.c.); Group 3: SN-intoxicated rats pre-injected with CRN (100 mg/kg, i.p.); and Group 4: SN-intoxicated rats pre-injected with N-CRN (100 mg/kg, i.p.). Curcumin and N-CRN were administered intraperitoneally 2 hour prior to SN intoxication. Hemoglobin concentration, serum tumor necrosis factor-alpha (TNF-α), and caspase-3 were analyzed. Gene expression of hypoxia inducible factor-1 (HIF-1α), matrix metallo-proteinases (MMP)-2, and tissue inhibitors of metalloproteinases (TIMPs)-2, as well as the protein expression of mitogen-activated protein kinases (MAPKs) and c-Jun NH2-terminal kinase (JNK) were examined in lung tissues.
Results
Hemoglobin level was markedly reduced, and serum TNF-α and caspase-3 were significantly elevated post SN intoxication. The lung MMP-2 and HIF-1α mRNA were overexpressed in the hypoxic group; while TIMP-2 mRNA was downregulated. Sodium nitrite administration increased proteins’ expressions of MAPK and JNK. Pretreatment with CRN or N-CRN markedly mitigated those alterations. These results were supported by histopathological examinations of lung tissue.
Conclusion
Interestingly, N-CRN exhibited a pronounced protective effect via suppression of inflammatory and apoptotic biomarkers and modulation of MAPK/JNK signaling pathway.
Keywords
Introduction
Oxygen is an essential element used by the cells of most organisms to produce adenosine triphosphate required for metabolic activities. Oxygen homeostasis is critically vital to maintain species survival 1 ; accordingly, the body develops multiple mechanistic pathways to balance the intracellular level of oxygen to maintain this homeostasis. When these mechanisms unsuccessfully proceed, a stress condition called hypoxia is initiated. Hypoxia, oxygen insufficiency, likely tends to increase reactive oxygen species (ROS) and alters the levels of antioxidant components, causing pulmonary oxidative damage and establishing inflammatory processes, such as redox state imbalance which may cause cardiovascular and neurodegenerative diseases and even cancer. 2

Oxygen homeostasis is preserved through a family of HIFs which are composed of hypoxia inducible factor-1 (HIF-1α and HIF-1β) subunits. 3 Activation or deregulation of hypoxia-induced transcriptional pathways may contribute to the pathogenesis of various conditions including atherosclerosis, chronic venous diseases, and pulmonary hypertension. 4 Hypoxia via the transcription factor (HIF-1α) induces many genes encoding proteins, which play a part in inflammation. 5 Inflammation represents a protective way to eliminate pathogens and initiate a process of wound healing. For instance, hypoxic cells may develop advanced mechanisms to control the inflammatory response to pathogens. Earlier studies demonstrated that HIF-1α in pulmonary fibroblast and cancer could be activated by c-Jun NH2-terminal kinase (JNK) and mitogen-activated protein kinases (MAPKs).6,7
The extracellular matrix (ECM) development and homeostasis, greatly regulated processes, are coordinated by a family of enzymes called matrix metallo-proteinases (MMPs) which can degrade all structural proteins of the ECM. 8 The activity of MMP family is controlled by tissue inhibitors of metalloproteinases (TIMPs). 8 Accumulating data revealed that the involvement of MAPKs in the regulation of gene expression can be through activating transcription factors and by targeting multiple post-transcriptional mechanisms. 9 Mammalian cells express several groups of MAPKs, including extracellular signal-regulated kinase (ERK)-1/2 and JNK. 10
Curcumin (CRN), a naturally occurring compound, is known to have a variety of pharmacological actions including antioxidant, anti-inflammatory, anticancer, and antimicrobial activities. 11 It is believed that the antioxidant activity of CRN is due to the presence of several functional groups in its structure that giving it the ability to protect bio-membranes against peroxidative damage. 12 Although it is a promising compound, its low water solubility and fast degradation profile may compromise its bioavailability. 11 Great effort has been done to enhance the bio-distribution of the original CRN, and recently its therapeutic efficacy has been improved by the application of the nano-therapeutics field. The development of nano-curcumin (N-CRN) has given a strong platform to gain most of the biological benefits vs the native drug. 13
The current study aims to compare the efficacy of CRN with N-CRN in the downregulation of the expression of inflammatory and apoptotic biomarkers, and modulation of MAPK/JNK signaling pathway in hypoxic lung induced by sodium nitrite (SN) in rats.
Material and Methods
Chemicals
Curcumin and SN and all other chemicals used in this study were purchased from Sigma-Aldrich Chemical Co., St. Louis, MO, USA. Nano-curcumin (particle size = 200 nm) was obtained from Lipolife Company, Drake Lane, Boreham, UK. Sodium nitrite was dissolved in normal saline, while CRN and N-CRN were suspended in 1% carboxy methyl cellulose (CMC) in normal saline.
Animals
Twenty four adult male Wistar albino rats, weighing 200 ±10 g, were obtained from the Animal Resource Unit, College of Pharmacy, KSU, SA. Animals were kept at standardized conditions and allowed to free access to water and chow diet. The animals were left for 1 week for acclimation. The protocol of this experiment was approved by the Scientific Research Ethics Committee, King Saud University (KSU-SE-19-60).
Experimental Design
The animals were randomly divided into 4 groups, 6 rats each. Group 1: served as control group and received CMC; Group 2: hypoxic group injected with a single subcutaneous dose of SN (60 mg/kg) 14 ; Group 3: SN-intoxicated rats and pre-injected with CRN (100 mg/kg, i.p.) 11 ; and Group 4: SN-intoxicated rats and pre-injected with N-CRN (100 mg/kg, i.p.). 11 Curcumin and N-CRN were administered 2 hour prior to SN intoxication.
One hour after SN injection, all rats were sacrificed under anesthesia. Blood samples were collected for serum separation, and lung was excised and washed in cold phosphate-buffered saline (PBS). Pieces from the lung were fixed in 10% neutral buffered formalin for histopathological study, and other samples were homogenized in cold PBS (10% w/v), while others were kept frozen at −80°C for gene and protein expression analysis.
Determination of Hemoglobin Level
Colorimetric technique was used to determine hemoglobin (Hb) levels according to the method of Kjeldsberg. 15
Determination of Serum TNF-α and Caspase-3 Using ELISA
Serum tumor necrosis factor-alpha (TNF-α) and caspase-3 were determined using ELISA kits (R & D systems MN, USA).
RT-PCR for MMP-2, TIMP-2, and HIF-1α Detection
Sequences of the Primers Used for RT-PCR.

Abbreviations: TIMP, tissue inhibitors of metalloproteinase, HIF-1, hypoxia inducible factor-1, RT-PCR, reverse transcription polymerase chain reacti.
Western Blot Analysis for MAPK and JNK Detection
The lung samples were homogenized in radioimmunoprecipitation assay buffer with proteinase inhibitors and centrifuged at 10 000 rpm, following this the pure supernatant was collected. Bradford protein assay kit was used to determine protein concentration, and 40 μg proteins were subjected to 10% SDS-PAGE and then transferred to nitrocellulose membranes. The membrane was blocked for 1 hour at room temperature and incubated with antibodies for MAPK (cat. no. ab199149), JNK (cat. no. ab112501), and β-actin (cat. no. ab8227) overnight at 4°C. The membranes were incubated with the secondary antibodies and developed using the enhanced chemiluminescence kit (BIO-RAD, USA). The obtained bands were quantified using ImageJ software, and the results were normalized as percent of control. 17
Examination of DNA Fragmentation in Lung Tissue
DNA fragmentation was examined by agarose gel electrophoresis. In addition, it was quantified in the lung tissues from control and treated rats as described by the method of Hickey’s group. 18 This technique is based on lysing the tissue followed by centrifugation to produce intact chromatin (pellet) and fragmented DNA (supernatant). After protein precipitation, the samples were treated with diphenylamine, and the absorbance of the developed color was measured at 600 nm.
Histopathological Examination
Deparaffinized sections of 4 μm of lung tissues were stained with hematoxylin and eosin and examined under light microscope.
Statistical Analysis
Statistical analysis was performed using one-way analysis of variance (ANOVA) by GraphPad Prism program followed by post hoc test. Data were expressed as means ± SEM, and the level of significance was set at P ≤ .05.
Results
Retaining the Hb Levels to Normal by CRN and N-CRN
Hemoglobin Levels in Control, Hypoxic, and Different Pretreated Groups.

Data are expressed as means ± SEM (n = 6). * = P ≤ .05 compared to hypoxic group, and + = P ≤ .05 compared to control group.
Abbreviations: SN, sodium nitrite; CRN, curcumin; N-CRN, nano-curcumin.
Attenuation of the Inflammatory and Apoptotic Markers by CRN and N-CRN
Serum TNF-α and caspase-3 proteins were significantly elevated post SN intoxication, while treatment with CRN and N-CRN produced marked reduction in their levels (Figure 1). Interestingly, N-CRN significantly downregulated TNF-α and caspase-3 compared to CRN. Effect of CRN and N-CRN on serum levels of TNF-α and caspase-3 following SN administration. Data are expressed as means ± SEM (n = 6). ***P-value <.001. Abbreviations: SN, sodium nitrite; CRN, curcumin; N-CRN, nano-curcumin; TNF-α, tumor necrosis factor-alpha.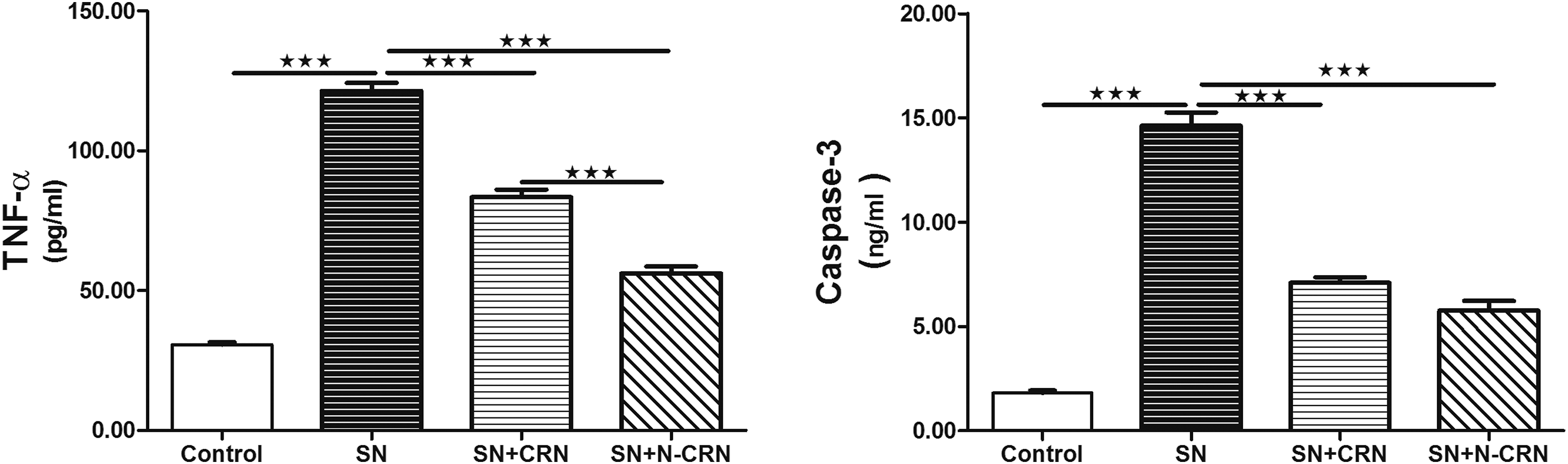
Modulation of the Expression of MMP, TIMP, and HIF-1α mRNA by CRN and N-CRN
The expressions of lung MMP-2 and HIF-1α mRNA were increased in the hypoxic group compared with the control group, while TIMP-2 was significantly downregulated (Figure 2). Pretreatment with CRN or N-CRN markedly improved these alterations compared with SN-intoxicated group. The improvement in the expression of lung MMP-2, TIMP-2, and HIF-1α mRNA produced by N-CRN was obvious compared to CRN. Effect of CRN and N-CRN on the expression of lung HIF-1 α, MMP-2 and TIMP-2 mRNA following SN administration. Data are expressed as means ± SEM (n = 6). *P-value ≤ .05, **P-value ≤ .01, ***P-value ≤ .001. Abbreviations: SN, sodium nitrite; CRN, curcumin; N-CRN, nano-curcumin; MMP-2, matrix metallo-proteinase-2; TIMP-2, tissue inhibitor of metalloproteinase-2, HIF-1, hypoxia inducible factor-1.
Downregulation of the Expression of MAPK and JNK Proteins by CRN and N-CRN
The proteins’ expressions of MAPK and JNK were significantly increased in the hypoxic group compared with the normoxic group. Pre-administration of CRN as well as N-CRN markedly modulated these deviations compared to the hypoxic group (Figure 3). Clearly, N-CRN produced a pronounced modulation of the expression of MAPK and JNK proteins vs CRN. Effect of CRN and N-CRN on the protein expression of lung MAPK and JNK following SN administration. Data are expressed as means ± SEM (n = 6). ***P-value ≤ .001. Abbreviations: SN, sodium nitrite; CRN, curcumin; N-CRN, nano-curcumin; JNK, c-Jun NH2-terminal kinase; MAPK, mitogen-activated protein kinase.
Alleviation of the Histopathological Changes Induced by SN Overdose by CRN and N-CRN
Figure 4A showed normal control of lung section of the normal control group which characterized by normal appearance of lung tissues. The lungs of rats exposed to SN toxicity exhibited degeneration and necrosis in addition to binucleated cellular infiltration (Figure 4B). Rats that received CRN or N-CRN showed reduction in the pathological changes in lungs’ architectures (Figure 4C and D). Histological examination (H&E, x400) of lung’s sections from the normal control group, hypoxic group, and treated groups. (A) Normal control section shows normal alveoli with thin inter-alveolar septum (arrow), and type I and type II pneumocytes were also seen (star); (B) lung section from rats exposed to SN shows alveoli with thick inter-alveolar septum (arrow), the septum is infiltrated by large number of lymphocytes (star); and (C) and (D) lung sections from rats received CRN or N-CRN showed improvement of the pathological changes in lung architecture. Abbreviations: SN, sodium nitrite; CRN, curcumin; N-CRN, nano-curcumin; H&E, hematoxylin and eosin.
Reduction of the DNA Fragmentation Induced by SN Overdose by CRN and N-CRN
Lung tissue from the hypoxic group showed an exaggeration in DNA fragmentation which was reduced in CRN and N-CRN pretreated groups with the superiority of N-CRN (Figure 5). (A) Hypoxia caused a significant increase in the DNA fragmentation, while the use of antioxidants decreased the DNA degradation and (B) quantitative analysis of DNA fragmentation levels in control, hypoxic, and all treated groups. Data are expressed as means ± SEM (n = 6). ***P-value ≤ .001.
Discussion
Sodium nitrite is an inorganic salt which can be used in the food industry; it is commonly used as a fish and meat preservative. Therefore, the diet is generally the most common source of human exposure. In blood, SN is highly reactive with Hb, thus affecting hematopoiesis. The toxicology of SN has given a major concern due to its ability to induce methemoglobinemia—a condition in which there is a reduction in hemoglobin’s ability to transport oxygen. As Hb level is a sensitive indicator of the organism’s physiological state, the current study is conducted firstly to investigate the effect of an acute SN treatment on Hb level in rats. In this study, treatment with SN caused a significant decrease in Hb level compared with the control group. Pretreatment of hypoxic rats with either CRN or N-CRN markedly modulated the deviation in Hb level.
In a mouse model of hypoxia/reoxygenation which has clinically triggered a typical lung injury, Chen and colleagues revealed increasing of alveolar wall thickening, infiltration by neutrophils, alveolar hemorrhage, and increased levels of proinflammatory factors. 19 Hypoxia can induce the production of ROS through the mitochondrial electronic transport chain, 3 Nicotinamide Adenine Dinucleotide Phosphate Hydrogen oxidase, 20 and xanthine oxidase. 21 Under normal conditions, ROS are produced at low levels and act as intracellular messengers to maintain cell functions. 22 Excessive production of ROS in lung can initiate inflammatory responses by activating redox-sensitive transcription factors, including activator protein-1, HIF-1α. 23 Furthermore, it has been shown that the mouse model of hypoxia-induced pulmonary inflammation which is characterized by the upregulation of proinflammatory factors such as TNF-α. 23 Likewise, human cardiomyocyte cell lines exposed to hypoxia have shown an increased production of ROS and apoptosis. 24 The results from these studies are parallel with the results obtained from current study as serum level of proinflammatory marker (TNF-α) and apoptotic marker (caspase-3) were significantly elevated post SN intoxication, while pretreatment with CRN and N-CRN produced marked reduction in their levels.
Herein, hypoxia caused significant increase in the gene expression of MMP and HIF-1α and marked decrease in the expression of TIMP. These results were equivalent to the results of a study done by Osinsky and coworkers where they revealed that the increasing of tumor hypoxia level was clearly related to tumor growth and positively associated with the activity of MMP in lung and negatively related to TIMP expression. 25 Matrix metallo-proteinases play a crucial role in the ECM remodeling and homeostasis; abnormal MMP-1 expression is detected in serious diseases and always linked with poor patient prognosis.26-28 Additionally, it has been shown that the expression of MMP-1 can be induced by exogenous sources of ROS, such as cigarette smoke and UV irradiation.29-31 Previously, it was revealed that the increasing of the intracellular redox level of tumor cells can drive MMP-1 transcription via redox-mediated JNK and MAPK signaling pathways. 32
Mitogen-activated protein kinases/JNK cascades play a vital role in the transduction of extracellular signals to cellular responses. 10 The MAPK pathways are involved in a series of protein cascades which play an essential role in the regulation of cell proliferation. 10 c-Jun NH2-terminal kinase pathway is implicated in both apoptosis and survival signaling, and it has been reported that UV-induced apoptosis in fibroblasts requires JNK to induce releasing of cytochrome c from the mitochondria, but the mechanism is unclear. 33 Additionally, JNK is activated by multiple stimuli, including TNF-α and ROS. The phosphorylated-JNK can be recruited outside mitochondrial membrane by TNF-α, leading to generation of ROS and sustained activation of JNK. Upon stimulation of TNF-α, caspase-8 can be initiated and directly converted to caspase-3, leading to cell death.
Alternatively, caspase-8 can cleave Bcl-2 interacting protein which triggers mitochondrial release of cytochrome c and formation of apoptosome, leading to activation of caspase-3. 34 Furthermore, Xi and colleagues have shown that MAPK/JNK signaling was markedly expressed in hypoxic/reoxygenated rat myocardial cell lines. 35 The results of the current study were parallel to the previous studies as the hypoxic rats exhibited significant increase in the expressions of MAPK and JNK. These observations were comparable with what was observed in a mouse model of hypoxia/reoxygenation which was triggered a typical lung injury. 23
In the current work, it was revealed that the protective effect of N-CRN on injured lungs of rats administered to SN could be through its antioxidant and anti-inflammatory properties. The antioxidant property of CRN can be seen in its role in decreasing of the peroxidation of membrane lipids and oxidative damage of genes and proteins which are thought to be associated with a wide range of chronic pathological complications such as cancer, cardiovascular, and neurodegenerative diseases. 36 Curcumin is believed to play an essential role against these pathological diseases due to it scavenging activity against ROS. Likewise, our study revealed the greater anti-inflammatory and anti-apoptotic effects of N-CRN than CRN. Curcumin has a powerful anti-inflammatory activity which is equivalent to non-steroidal anti-inflammatory drugs such as indomethacin. 36 The anti-inflammatory action of CRN is thought to be mediated through inhibition of the inducible nitric oxide synthase (iNOS), TNF, and the transcription factors like nuclear factor-κB (NF-κB). 36 Additionally, the observations obtained from histopathological analysis of the N-CRN group provided substantial evidence in the reduction of lung degeneration, necrosis, and cellular infiltration in hypoxic lung induced by SN intoxication. Recently, our research group showed that CRN attenuated the apoptotic marker (BCL-2-associated X (Bax)) and DNA damage induced by lead acetate. 37 Additionally, our previous study revealed that the oxidative stress and inflammation have been mitigated by liposomal CRN. 38
To sum up, the current findings may have important implications on the development of a new therapeutic strategy aimed at using N-CRN as a supplement drug for protection against lung injury. Furthermore, this study gives an insight into the reasons that make N-CRN a more therapeutically advanced supplement than its native drug.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant from the Research Center of the Female Scientific and Medical Colleges, Deanship of Scientific Research, King Saud University, Saudi Arabia..