
Abstract
Neutron irradiation which could trigger severe biological effects, is being applied in nuclear plants, radiotherapy, and aerospace gradually. Low dose hyper-radiosensitivity response of low Linear Energy Transfer (LET) irradiation on the cell survival has become a matter of great interest since its discovery, but a few research have been done on this response induced by neutron irradiation. To investigate this response induced by neutron irradiation, Caenorhabditis elegans (C. elegans) was irradiated by neutron irradiation. The surviving fraction of C. elegans on the 12th day after irradiation was analyzed, which showed a hyper-radiosensitive response at low doses and followed by an increase in survival fraction at slightly higher doses. The finding of this work that neutron irradiation decreased the surviving fraction in a non-dose-dependent manner was different from previous low-LET irradiation studies. To understand the experimental results, a DNA damage-repair model was introduced. By comparing experimental results with theoretical analyses, we suggest that the low dose hyper-radiosensitivity response of neutron irradiation may possible related to different radiation types and DNA damage recognition proteins and immune system of C. elegans.
Introduction
Neutron irradiation tends to be increasing gradually by considering more and more often radiation accidents, medical exposure, natural or workplace radiation sources in modern society. As the high-LET uncharged particles, neutrons transfer energy mainly through the interaction with the nucleus. Multiple experiments revealed that in contrast to photons and electrons, neutron irradiation could trigger severe biological effects such as chromosomal aberration, DNA damage, apoptosis, and necrosis. 1 -3 Furthermore, radiation protection requires investigating the neutron irradiation biological effects, which is relatively less studied than other radiations such as photons. 4 It is essential to study the biological effects of neutron irradiation, to understand the molecular mechanism and characteristics of neutron irradiation, which is crucial for precise risk evaluation in response to neutron irradiation and especially for radiation protection. 5
Currently, there has been sufficient data demonstrated the existence of low dose Hyper-Radio-Sensitivity (HRS) induced by low-LET radiation, which is characterized by a significantly lower cell survival at absorbed radiation doses less than 0.3 Gy and follows by an Increased Radio-Resistance (IRR) response at slightly doses between 0.3-0.6 Gy. 6 Since its identification, multiple experiments suggested that the HRS/IRR response commonly exists in many mammalian normal and malignant cell lines by using the various radiation doses and biological endpoints. In the case of high-LET radiation, such as neutron irradiation, there are not enough consistent experiments to confirm its existence. Marples et al 7 found that V79 cells irradiated by X-ray at doses below 0.6 Gy showed an HRS/IRR response, but it was absent for neutron irradiation and the survival fraction of neutron-irradiated cells decreased exponentially. In contrast, Claude et al 8 had identified an obvious cell killing at low doses of neutron irradiation, and followed by a plateau at higher doses in melanoma cell survival and cytogenetic aberration assays. In line with this work, they also found that an HRS response occurred only at a very low dose rate and was followed by an IRR response induced at a high dose rate in melanoma cells. 9 Xu et al 10 also provided evidence that HRS/IRR response may exist in neutron-irradiated C.elegans.
Early studies of low-LET radiation-induced HRS/IRR response could be evaluated well by fitting data to Joiner’s Induced Repair model. When the dose exceeds a certain threshold dose, the damage-repair system is activated. 6 Meanwhile, studies of heavy-ion beam irradiation on the survival of irradiated plants and microorganisms proposed a unique saddle-type dose-effect curve which is similar to HRS/IRR response 11,12 and subsequently a various model was established, such as the mass deposit effect model and the damage-repair model to explain the saddle-type dose-effect curve. 13 Recently, Madas and Drozsdik 14 had put forward a mathematical model to explain HRS/IRR response by using the principle of minimum mutation load considering that radiation-induced mutagenic DNA lesions and cell divisions are sources of mutations.
Genome dynamics plays a crucial role in population genetics and molecular evolution. 15 Focusing on the dynamics of evolution, there has conducted multiple studies, of which Crow and Kimura 16 have proposed a popular molecular quasispecies model where mutation and selection are assumed to be two parallel processes. The Crow-Kimura model describes an evolutionary process where mutations have resulted from environmental factors rather than self-replication. 17 Recently, Li et al 18 successfully established a damage-repair model of heavy 7 Li ions irradiated plants based on the Crow-Kimura model to explain the saddle-type dose-effect curve, where the dry seeds of irradiated maize plants were considered as independent individuals and the germination process was a dynamic process. Subsequently, Hu et al 19 also proposed a radiation mutagenesis model with the same mathematical form of the Crow-Kimura model, described a dynamic evolution from microscopic radiation damage and repair activation to the final macroscopic biological effects to reveal the mechanism of saddle-type dose-effect curve.
In this work, C.elegans was irradiated by neutron and an HRS/IRR response was obtained. The shape of HRS/IRR dose-response in worms was characterized by introducing radiation dose and immunization effect into the Crow-Kimura model of biological evolution. The experimental results were compared with the theoretical model to validate the model. It was suggested that fast neutron irradiation could induce HRS/IRR response in C. elegans.
Materials and Methods
Worm Strains and Synchronization
The wild-type C.elegans N2 Bristol used in this study was obtained from the Caenorhabditis Genetics Center (St. Paul, MN, USA). Worms were maintained using standard procedures. 20 The worms were placed on the Nematodes Growth Medium (NGM) agar by a sterile platinum worm picker and the medium was seeded with E. coli OP50 strain at 20 oC and 80% relative humidity. The NGM agar medium was prepared as described previously by Stiernagle. 21 To obtain the synchronized young worms, the gravid hermaphrodites were quickly transferred onto NGM agar medium for a 2-hour spawning period and then removed with a worm picker. The acquired embryos were cultured at 20 oC for 14 hours in incubation. The next day, larvae were checked to avoid mixing with other phase worms to obtain the synchronized L1 phase worms.
Neutron Irradiation
The worms were irradiated with 14.1 MeV single-energy neutrons generated by the High-Intensity D-T fusion Neutron Generator (HINEG) which was developed by the Institute of Nuclear Energy Safety Technology of the Chinese Academy of Sciences (Hefei, China) by the Fusion Design and Study team. 22 Before irradiation, all petri dishes (φ = 3.5 cm) were fixed up vertically with the radiation beam on a plastic foam board to ensure homogeneous exposure of worms. The depth of the agar in the dish was measured carefully to ensure that the distance between worms and neutron source was accurate. The distribution of irradiated neutron dose was broad and extended from 0.171 ± 0.011 Gy to 1047 ± 63 Gy. After irradiation, the worms were cultured in an incubator at 20 oC for general experiments immediately after irradiation.
Lifespan Assay
Lifespan of C. elegans was performed starting with worms in synchronized L4 phase at 20 oC without interference and with adequate food. During the reproduction period, worms were transferred into fresh NGM plates every day until the end of the egg-laying period. The viability was evaluated every day with a gentle touch of a worm picker to count the numbers of alive and dead worms. If worms failed to wriggle in response to the gentle nose touch, they were considered to be dead. If worms were wall-crawling, having internal hatching, or missing, they were not considered into the total.
Statistical Analysis
All results were analyzed with the OriginPro 8.0 statistics software and were presented as means ± standard deviations.
Results
Neutron Irradiation Decreased the Worm’s Surviving Fraction in a Nonlinear Manner
Post neutron irradiation, the surviving fraction of C.elegans had no significant changes for the most of the days, but on the 12th day after neutron irradiation, the surviving fraction of C.elegans decreased obviously in a non-dose-dependent manner. As shown in Figure 1A, compared with non-irradiated control groups, the average surviving fraction count of 3.52 Gy decreased to 0.59 ± 0.10 and presented the maximum damage effects. Interestingly, compared with the 3.52 Gy group, the higher doses of 6.85-8.92 Gy did not generate a worsen effect of a shorter surviving fraction and prolonged with a slightly increase to 0.67 ± 0.11 and 0.80 ± 0.02 respectively. As the dose increased greater than 11.3 Gy, the intensity of resistance response reached a maximum and the surviving fraction decreased exponentially in a large-scale view. These results were similar to previous cell survival studies that showed a low dose hyper-radiosensitivity and an increased radio-resistance with increasing dose. 7 And as shown in Figure 1B, Ye et al 23 have reported that the survival fraction of C.elegans on the 12th day decreased in a dose-dependent manner with no significant HRS/IRR response was observed in the experimental results of 60Co γ irradiation.

Effects of irradiation on the survival fraction of the twelfth day of C.elegans. (A) Survival fraction of the twelfth day after neutron irradiation, results are logarithmic. (B) Survival fraction of the 12th day after 60Co γ irradiation.
Model Construction
Based on the model of a damage-repair model of plant mutagenesis induced by ionizing radiation of Li, 24 we assumed that neutron-irradiated worms were independent individuals, the survival process of worms was an individually dynamic process and all of the irradiated worms formed a biological population. The worms were divided into three states: natural state, HRS/IRR state, and fatal state. The natural state worms represent the worms that have not been damaged by neutron irradiation, the HRS/IRR state worms represent the worms that can still survive with repaired-damage and the fatal state worms represent the worms that have been killed with severe damage by neutron irradiation. Based on the studies of Li, we decided to establish a similar damage-repair model to verify the HRS/IRR response. The transition between different states of C.elegans induced by neutron irradiation is shown in Figure 2.

The transition between different states of C.elegans induced by neutron irradiation. f0, f1 , and f2 are the fitness values of each state, indicating the reproductive efficiency of worms under neutron irradiation conditions; x0, x1 , and x2 are the relative concentrations of each state in the biological population exposed to neutron irradiation; m10 and m12 are radiation damage coefficients; r01 and r21 are damage repair coefficients.
The dynamic process of the above states can be expressed as:

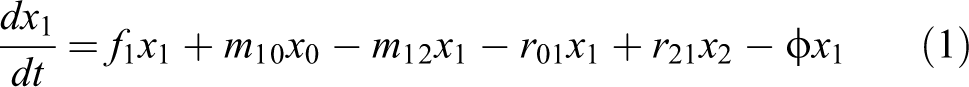

where f0 , f1 , and f2 are the fitness values of each state, indicating the reproductive efficiency of worms under neutron irradiation conditions. x0 , x1 , and x2 are the relative concentrations of each state in the biological population exposed to neutron irradiation. All of them were normalized to x0 +x1 + x2 = 1; m10 and m12 are radiation damage coefficients; r01 and r21 are damage repair coefficients; φ is an input stream to maintain a constant biological group size and this value was numerically equal to the average fitness degree of the irradiated population.
Multiple studies have shown that ionizing radiation could induce various types of DNA damage in cells, occurring at different frequencies, of which DNA Double-Strand Breaks (DSBs) was the most crucial damage. 25 Neutron irradiation can cause various complex microscopic damage at different levels of molecular, cellular, and tissue. After a certain damage accumulation process and a series of amplification effects of biochemical reactions, the microscopic molecular damage will lead to the change of the macroscopic state of C.elegans eventually. Furthermore, it has been found that DSBs may be affiliated with the HRS/IRR response, so we assumed that the yield of DNA damage induced by neutron radiation is the primary microscopic damage and is satisfied by a linear-quadratic function relationship as follows:

where the values of α and β are characteristic of the cells type and conditions of irradiation; µ(D) is the rate of radiation-induced DNA damage; D is the value of neutron irradiation dose.
Given that C.elegans lacks the adaptive immune system and relies on the innate immune system to activate the corresponding signal transduction pathways and produce effector molecules for immune defense. 26 Based on its innate immune and repair ability, the characteristic of microcosmic repair process can be expressed as:
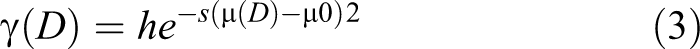
where γ(D) is the rate of the microcosmic repair process; h is the value of repair intensity; s and µ0 are the value of repair range and repair threshold dose respectively.
Equation (1) can be expressed as:



where X(t) = (x0, x1, x2 )T is the relative concentration vector for each state; F= (f0, f1, f2) T is the fitness value matrix for each state; M′ and R′ are the neutron irradiation damage and repair coefficient matrix, respectively. 17 The mathematical representation of the matrix of the Crow-Kimura model is given in Eq. (6). 27 Equations (6) and (5) have the same mathematical form as the Crow-Kimura model, namely the neutron irradiation damage-repair model of C.elegans can be treated as a peculiar form of the Crow-Kimura model.
Steady-State Numerical Analysis
Researches on the effects of ionizing radiation on organisms revealed that the interaction between radiation and organisms is an extremely complex process, including multiple processes such as physical, chemical, biochemical, and biological processes. These complex reactions usually last only a few seconds but are accompanied by biological effects occurred in several days or months. 28 Thus the biological effects reflected as microscopic damages will induce individual state changes eventually. Based on studies of Li, 24 when the time approaches infinity, the macroscopic state of C.elegans would become stable slowly and reached a steady-state. In the conditions of steady state, the character of C.elegans population is dependent on doses rather than on evolution time. Referring to the steady-state solution of the Crow-Kimura model, the relative concentration distributions of the three states were obtained, 29 and given as follows:



During the growth of C.elegans, dying individuals would die eventually, thus the survival fraction of C.elegans after neutron irradiation could be obtained from the lethality rate and given as follows:

Model Fitting of Neutron Irradiation-Induced Effect-Dose in C.elegans
The surviving fraction of the 12th day after neutron irradiation was fitted with the Levenberg-arquardt and global optimization search method. The data points were well fitted (goodness of fit, R2 = 0.99). The theoretical curve of the relationship between survival fraction and neutron dose is shown in Figure 3A. It shows that the experimental survival fraction is well consistent with the theoretical calculation. The convergence condition was reached by calculation and the following parameters were obtained. Namely, the relevant parameters were: α = 0.12, β = 0.009, h = 7.88, s = 0.5 and D0= 14.09 Gy. Where D0 is the threshold dose of the repair process, and beyond this threshold, the effect of repair weakens continuously and organisms will rapidly die due to the sharp increase in damage. According to the above parameters, the relative concentration distribution of tri-states C.elegans with radiation dose was obtained under steady-state conditions theoretically as shown in Figure 3B. And Figure 3B shows that the relative concentration distribution of tri-states and the damage-repair effects varied with the dose of D0. Both models show a consistent trend demonstrating that the experimental survival fraction is consistent with the theoretical calculation. Therefore, the damage-repair model is comparatively reasonable.

Theoretical analysis of survival fraction and dose of neutron irradiation on the twelfth day. (A) The relative concentration distribution of tri-states in steady state induced by neutron irradiation. (B) The comparison of theoretical results with experiment data. The results are logarithmic.
Discussion
In the present study, by aiming at studying how the varying doses of neutron irradiation could impact the lifespan of C.elegans, it has been found that neutron irradiation affects the lifespan of worms negatively. Importantly, by using the lifespan assay, we have discovered a significant damage effect in 3.52 Gy, followed by a radio-resistance effect in 6.85-11.3 Gy on the 12th day after irradiation, beyond the dose of 11.3 Gy, the surviving fraction of C.elegans decreased exponentially in a large-scale view and showed an HRS/IRR response. Based on the studies of Li, 18 we established a DNA damage-repair model to describe the HRS/IRR type dose survival fraction curve caused by neutron irradiation in C.elegans. The theoretical model results were consistent with the experiment results and indicated that the HRS/IRR response was a dynamic process from microscopic damage repair to macroscopic state evolution. By model fitting, the theoretical threshold dose D0 of the repair process was 14.09 Gy other than 11.3 Gy in the actual experiment on account of the discontinuity dose selection. At the neutron irradiation dose below 3.52 Gy, the damaging effect played a dominant role, resulting in the first prominently peak of lower survival. While in the dose range of 6.85-11.3 Gy, the repair effect became essential, leading to slightly higher survival, beyond the dose of 11.3 Gy, the repairability of C.elegans decreased gradually, causing a dramatic increase in damages and deaths. Taken together, our studies indicated the presence of HRS/IRR response caused by neutron irradiation in C.elegans and we speculated that it may be related to the initiation of the immune system of worms and the combination of the DNA damage and repair effect.
Accumulating cell studies have highlighted the HRS/IRR response commonly exists in many mammalian normal and malignant cell lines with various biological endpoints. By using clonogenic survival assay, Short et al 30 proposed that HRS response existed primarily in G2-phase enriched cells, with little or no evidence in G1 and S cell cycle phases in the X-ray irradiated human melanoma cell lines by using clonogenic survival assay. Enns et al 31 found that the human A549 lung carcinoma and T98G glioma cells showed a marked hypersensitivity after the exposure of γ ray irradiation and suggested that p53-dependent apoptosis was an important factor of low-dose hypersensitivity. Increased HRS/IRR response to relatively low doses of low-LET irradiation has been observed in several but not all tumor and human normal cell lines with the different endpoint studies, but there are not enough consistent experiments to confirm its existence of high-LET irradiation.
The HRS/IRR response in C.elegans induced by neutron irradiation in our studies could be related to the initiation of the immune system of worms. In C.elegans, it is found that it can only rely on the innate immune system to activate the corresponding signal transduction pathways and produce effector molecules for immune defense. 26 Murphy et al 32 found that the daf-2/daf-16 pathway overlays with the DBL pathway and the MAPK pathway, and the interaction between the pathways forms a complex signal transduction network by upregulating a variety of genes, which cooperatively determines the lifespan, stress, development and immune defense of worms. Through recruiting complex signalling pathways (DAF-16, SKN-1/Nrf, oleic acid, lipase, etc.), germline ablation can also enhance the effects of immune defence, proteostasis and stress resistance. 33 Also, the HRS/IRR response in C.elegans induced by neutron irradiation in our studies could be related to the different complexities of the DNA damage induced by the different radiation types. Comprehensive researches have been studied that the damage response to DNA DSBs could trigger an intricate signaling network to regulate checkpoints to induce cell cycle arrest and repair of lesions. Hofmann et al 34 found that HUS-1, a nuclear protein, would be relocated to putative sites of DSBs as a conserved checkpoint to regulate DNA damage-induced cell cycle arrest and apoptosis and it also demanded genome stability in irradiated C.elegans. Consistent with this work, Li et al 35 had found that MRT-2/HUS-1/CEP-1/CED-4 proteins had an indispensable role in germ cell apoptosis in proton radiated C.elegans. By the combination of functional genomic mapping approaches in C.elegans, Boulton and his colleagues also suggested MRT-2, HUS-1, and HPR-9 genes and RAD9/RAD1/HUS-1 complex played the part of DNA damage sensor in DNA damage response for proper localization and were functionally conserved in C. elegans. 36,37 Thus we propose that low dose neutron irradiation may cause DNA DSBs, increasing the expression of HUS-1 gene to recognize aberrant structures of DNA and propagate signals to pivotal cellular processes to repair the damages of C. elegans.
In conclusion, we have found the HRS/IRR response in neutron-irradiated C.elegans and established a DNA damage-repair dynamic model to explain this phenomenon. Whereas, it should be noted that our model has some drawbacks that the energy transmission line density, non-target effect, and pretreatment conditions needed to be considered further. At the same time, the dose selection of our experiment was discontinuous and limited. In further work, the mechanism of HRS/IRR response still needs further investigation.
Footnotes
Authors’ Note
Guangyan Feng, Zhao Xu, and Taosheng Li conceived and designed the experiment. Guangyan Feng and Zhao Xu performed the experiments. Guangyan Feng, Lianxin Zhang, and Zhanguo Yang analyzed the data. Yong Zhang and Siwei Zhang maintained the normal operation of the machine. Guangyan Feng wrote the article.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by the National Magnetic Confinement Fusion Science Program of China (2014GB112006) and the Chinese Academy of Science Dean Found (YZJJ2020QN31).