
Abstract
Sarcopenia is often regarded as an early sign of weakness and is the core element of muscle weakness in elderly individuals. Sarcopenia is closely related to the reduction of exercise, and elderly individuals often suffer from decreased muscle mass and function due to a lack of exercise. At present, studies have confirmed that resistance and aerobic exercise are related to muscle mass, strength and fiber type and to the activation and proliferation of muscle stem cells (MuSCs). Increasing evidence shows that microRNAs (miRNAs) play an important role in exercise-related changes in the quantity, composition and function of skeletal muscle. At the cellular level, miRNAs have been shown to regulate the proliferation and differentiation of muscle cells. In addition, miRNAs are related to the composition and transformation of muscle fibers and involved in the transition of MuSCs from the resting state to the activated state. Therefore, exercise may delay sarcopenia in elderly individuals by regulating miRNAs in skeletal muscle. In future miRNA-focused treatment strategies, these studies will provide valuable information for the formulation of exercise methods and will provide useful and targeted exercise programs for elderly individuals with sarcopenia.
Introduction
With the aging of the global population and the improvement of living standards, sarcopenia in elderly individuals has gradually become a popular research topic in the field of geriatric medicine. Data from the Asian Working Group for Sarcopenia in 2016 show that the prevalence of sarcopenia in the elderly Asian population is 4.1%-11.5%. 1 Sarcopenia in elderly individuals is often considered an early manifestation of weakness and can lead to declining physical function, impair the ability to perform daily life activities, and increase adverse outcomes such as falls, disability, hospitalization and so on. 2 The pathogenesis of sarcopenia is complex and mainly includes chronic inflammation, insulin resistance, mitochondrial dysfunction and motor neuron degeneration; age-related hormonal metabolic disorders and genetic factors also contribute to sarcopenia. 3 In addition, sarcopenia in elderly individuals is closely related to decreased exercise. Elderly individuals often suffer from decreased muscle strength and endurance due to a lack of exercise. After the age of 75, approximately 1/3 of men and 1/2 of women do not participate in exercise. Exercise is one of the most effective means to obtain and maintain muscle mass and strength. MicroRNAs (miRNAs) are regarded as the most promising class of small molecule RNAs for targeted diagnosis and treatment in the 21st century. 4 MiRNAs target large numbers of genes, and approximately 60% of human genes are regulated by miRNAs. 4,5 MiRNAs, which regulate cell function by inhibiting or promoting the expression of mRNAs, play a key role in regulating the growth, development and metabolic balance of skeletal muscle. 6 The discovery of the mechanism by which miRNAs regulate cytokine metabolism during exercise and the role of miRNAs in the pathogenesis of sarcopenia has initiated an upsurge in related research in recent years. MiRNA expression profile analysis has shown that miRNAs play a key role in regulating the muscle fiber number, the muscle fiber cross-sectional area, the muscle fiber type and MuSCs during aerobic and resistance exercise.
Sarcopenia and Exercise
Aging muscles retain a certain degree of plasticity. Specific exercise and nutrition interventions can delay the loss of aging muscles and rebuild aging muscle tissue to a certain extent. These methods are also the most important means of treating and preventing muscle loss in clinical and rehabilitation medicine. 7 Although there are no consensus recommendations on physical activity and exercise for elderly individuals with sarcopenia, there are specific guidelines for various types of activity for people over 65 years of age. 8
Resistance exercise involves the active movement of muscles to overcome external resistance, and with the improvement of exercise ability, the demand of the exercise intensity gradually increases. 9 Resistance exercise training is considered an important strategy to prevent muscle atrophy 10 because it stimulates muscle hypertrophy and increases muscle strength. Resistance training increases the production of anabolic-related hormones and reduces the activity of catabolism-related enzymes. These changes increase the efficiency of protein synthesis in muscle cells and shift the balance between muscle protein synthesis and degradation toward synthesis. 11,12 Regular resistance exercise increases the cross-sectional area of muscle fibers, especially fast muscle fibers, by promoting the proliferation of MuSCs. 13,14 Muscle protein synthesis and muscle fiber hypertrophy increase the force production, muscle mass and physical performance. However, resistance exercise training has disadvantages; in particular, it has little effect on mitochondrial protein expression and mitochondrial function, which are considered to be potential causes of age-related sarcopenia. 15
Aerobic exercise refers to physical exercise carried out by the human body under the condition of an adequate supply of oxygen and is characterized by rhythmic and repetitive contraction of large muscle groups. 16 Aerobic exercise causes skeletal muscle mitochondria to produce ATP, which improves aerobic capacity, metabolic regulation and cardiovascular function. Increased ATP production supports mitochondrial biogenesis and kinetic balance, restores the mitochondrial metabolism, reduces catabolic gene expression and increases muscle protein synthesis. 17 Aerobic exercise controls the mRNA expression of the muscle growth inhibitor myostatin. Twelve weeks of aerobic training enhanced the mitochondrial biogenesis and fusion in elderly subjects. 18 In addition, aerobic exercise activates MuSCs, increases muscle fiber capillary blood flow, improves mitochondrial function, and repairs damaged muscle fibers. 19 In general, aerobic exercise improves mitochondria-related problems, promotes efficient energy use, counteracts muscle hypertrophy, and increases muscle strength.
Most studies have focused on the effects of individual exercise methods on sarcopenia. However, aerobic training has little effect on improving muscle mass and strength. Resistance training increases the risk of injury, reduces the participation rate, and is considered by patients to be too boring due to repetitive movements. In addition, due to age-related impairments in mTOR signaling, which is involved in muscle protein synthesis, the effects of resistance exercise are diminished in elderly individuals. 20 It seems that no single exercise can meet all of the requirements of therapeutic exercise for age-related sarcopenia. Therefore, it is recommended that a comprehensive exercise plan combining aerobic and resistance exercise be used to treat sarcopenia in elderly individuals. 21,22
MiRNA and Exercise
MiRNA Structure and Function
More than a decade ago, small noncoding miRNAs were discovered in Caenorhabditis elegans, and homologues were identified in other invertebrates and humans. 23 MiRNAs are small ubiquitous noncoding RNA molecules that contain approximately 22 nucleotides and are highly conserved in eukaryotes. 24,25 MiRNAs guide the RNA-induced silencing complex (RISC) to degrade mRNA or hinder its translation by base pairing with mRNA. Therefore, miRNAs can be used as powerful regulators of gene expression at the posttranscriptional level. 26 Each individual miRNA can bind hundreds of target mRNAs, which have complementary binding sites in their 3′UTRs; thus, miRNAs act as posttranscriptional regulators at the gene expression and protein levels. 27 In addition, some mRNAs can be regulated by a variety of miRNAs; of course, some mRNAs are not regulated by miRNAs.
MiRNAs act as part of a gene regulatory network that controls many biological processes. In recent years, through comprehensive analysis of mammalian transcripts, a large number of miRNAs have been identified, and abnormal miRNA expression has been shown in many disease states. 28 Recent studies have also confirmed that circulating miRNAs are closely related to the occurrence and development of sarcopenia. 29 Both aerobic and resistance exercise promote the active adaptation of skeletal muscle, thereby enhancing muscle mass, strength and endurance, which in turn contribute to the promotion of healthy aging. 30 In animal models and humans, miRNAs have been shown to participate in skeletal muscle movement responses. In-depth research of miRNAs is helpful for the diagnosis, treatment and prognosis of certain diseases.
MiRNAs and Aerobic Exercise
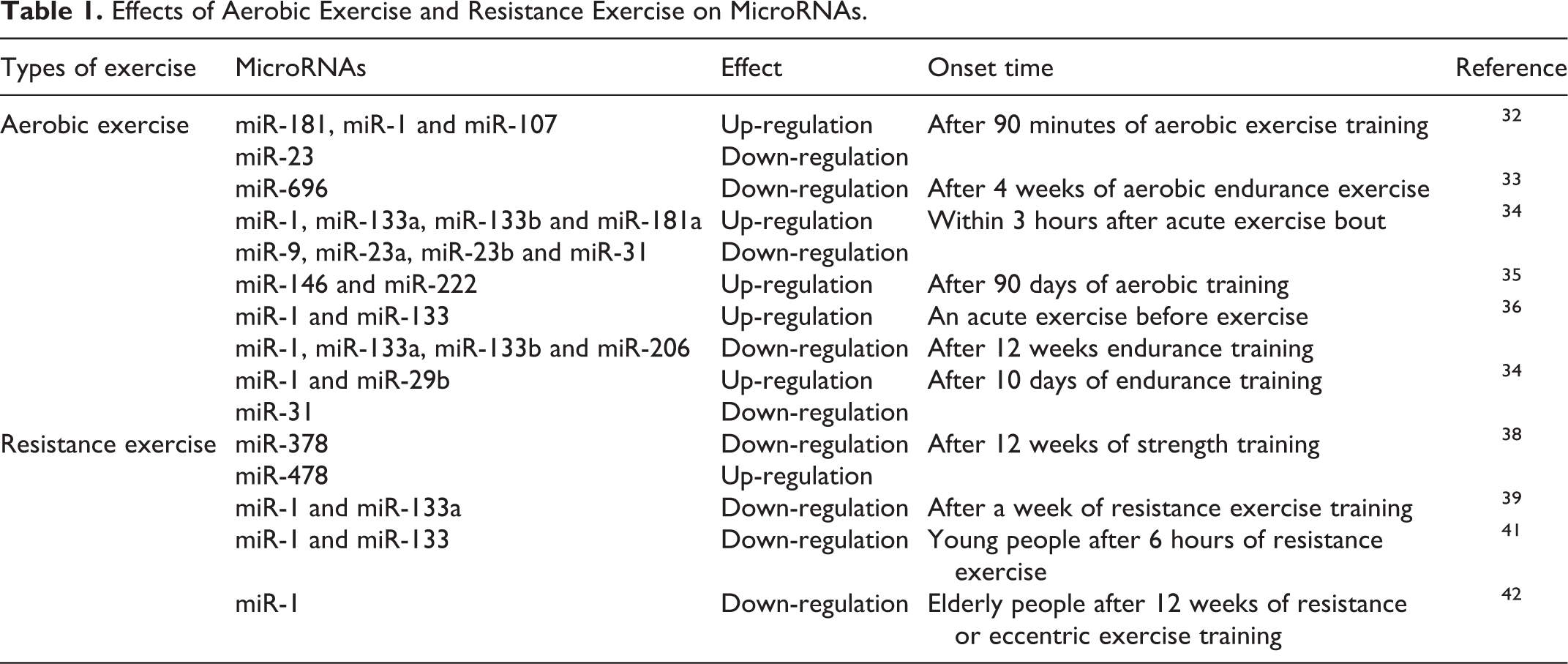
Skeletal muscle is an adaptive organ with extraordinary versatility, and it can respond to mechanical load through cell-autonomous regulatory mechanisms. 31 Aerobic exercise training can improve muscle metabolism, thus enhancing physical strength. In a mouse model, it has been shown that 90 minutes of aerobic exercise upregulates miR-181, miR-1 and miR-107 and downregulates miR-23. 32 A key regulator of mitochondrial biogenesis, peroxisome proliferator-activated receptor γ coactivator-1 (PGC-1), contains sites that are predicted to bind to miR-23a; therefore, the downregulation of miR-23a will reduce the inhibition of PGC-1. Moreover, miR-696 decreases after 4 weeks of aerobic endurance exercise but increases again after 5 weeks of training. 33
One study found that the levels of miR-1, miR-133a, miR-133-b and miR-181a increased within 3 hours after acute exercise. In contrast, the levels of miR-9, miR-23a, miR-23b and miR-31 decreased. Short-term training increased the miR-1 and miR-29b levels, while the miR-31 levels decreased. 34 Baggish et al. 35 reported that after 90 days of aerobic training, the expression of miR-146 and miR-222 increased, but the expression of miR-21 and miR-221 did not change. After aerobic exercise, a positive correlation was observed between the level of miR-146 and the VO2 max. It has been reported that acute exercise before exercise training increases miR-1 and miR-133 expression in skeletal muscle and that acute exercise after exercise training does not have this effect. 36 In addition, after 12 weeks (long-term) endurance training, the expression levels of some muscle-specific miRNAs (myomiRs), such as miR-1, miR-133a, miR-133b and miR-206, were significantly downregulated and returned to the baseline expression level 2 weeks after training. 36 In another study, 10 days of endurance training resulted in an increase in skeletal muscle miR-1 and miR-29b levels and a decrease in miR-31 levels. 34
MiRNAs and Resistance Exercise
The meta-analysis of the effects of moderate and heavy load resistance training on muscle mass and strength in elderly individuals recommends resistance exercise training in middle-aged and elderly people to increase or maintain skeletal muscle mass. 37 However, the muscle mass obtained by individuals varies greatly, and these differences are usually accompanied by the expression of different miRNAs. After 12 weeks of strength training, the expression of miR-378 decreases, and the expression of miR-478 increases. In addition, the change in miR-378 expression is closely related to the lean weight. Further, Davidsen 38 and others found that after resistance exercise, an increase in skeletal muscle was related to selective changes in miRNA expression, and the increased skeletal muscle volume was positively correlated with the expression of miR-378.
In a mouse study, 39 the expression of muscle-specific miR-1 and miR-133a was downregulated by approximately 50% after a week of resistance exercise training. In a study of young healthy men, 40 the subjects were divided into 3 groups: the muscular strength endurance (SE), muscular hypertrophy (MH) and maximum strength (MS) groups. It was found that the levels of 2 miRNAs (miR-208b and miR-532), 6 miRNAs (miR-133a, miR-133b, miR-206, miR-181a, miR-21 and miR-221) and 2 miRNAs (miR-133a and miR-133b) changed significantly in response to the SE, MH and MS protocols, respectively. Other studies found that compared with the baseline, only young people exhibited decreased miR-1 and miR-133 levels after 6 hours of resistance exercise, while the level of the skeletal muscle primary miRNA (pri-miRNA)-206 increased at different time points after exercise in elderly and young men. 41
It has been reported that in elderly men and women, miR-1 was downregulated after 12 weeks of resistance or eccentric exercise training, while the expression of IGF-1, which promotes muscle growth, increased in parallel. 42 Although each type of exercise leads to different molecular and structural adaptations in muscle, changes in miR-1 expression may reflect muscle remodeling because resistance exercise is the only type of exercise that increases the cross-sectional area of the muscle.
The effects of aerobic exercise and resistance exercise on microRNAs are shown in Table 1.
Effects of Aerobic Exercise and Resistance Exercise on MicroRNAs.
MiRNA-Mediated Exercise Delays Sarcopenia
MiRNAs and Muscle Mass
Beginning at 30 years of age, lean mass, which is mainly composed of muscle and visceral organs, begins to gradually decline. Muscle mass continues to decrease throughout the middle age years and accelerates at 60 years of age, and weight gradually decreases due to muscle loss. 43 The loss of muscle mass is manifested by decreases in the muscle fiber number and muscle cross-sectional area. MiR-1, miR-133a, miR-133b, miR-206, miR-208a, miR-208b, miR-486 and miR-499 are the main miRNAs involved in the proliferation, differentiation and apoptosis of skeletal muscle cells, and dysfunctional miRNA expression leads to different physiological and pathological states. 44 MiR-1 regulates MyoD1 protein expression mainly through the mTOR signaling pathway, and miR-1 directly inhibits myostatin (MSTN), which also regulates MyoD1 protein expression. 45 MiR-1 also interacts with some of its targets; for example, miR-1 increases the expression level of the target MEF2, and MEF2, in turn, upregulates the expression of miR-1. 46 Additionally, IGF-1, which is involved in the growth, development and metabolic balance of skeletal muscle, regulates the expression of miR-1 through the IGF-1/AKT/FOXO3/miR-1 axis. 47 MiR-1 and miR-206 regulate muscle oxidation levels and metabolic balance by controlling glucose-6-phosphate dehydrogenase (G6PD). 48 Dai et al 49 found that miR-133a knockout mice showed central nuclear myopathy, mitochondrial dysfunction and muscle fiber morphological changes, and their skeletal muscle cells gradually began to undergo apoptosis. The target gene predicted for miR-208b and miR-499 is myostatin (MSTN), which is mainly involved in the regulation of skeletal muscle quantity. 50,51
It was reported that endurance exercise changes the expression levels of 39 miRNAs. 52 It is predicted that the miRNAs exhibiting increased expression target transcripts involved in inflammatory processes, metabolism and muscle atrophy, and miRNAs affect muscle atrophy through the reduction of mitochondrial biogenesis. 52 IGF-1 is known to be an effective promoter of muscle growth. After 12 weeks of resistance exercise training (RET) or eccentric ergometer training (EET), regulation of miR-1 led to significant increases in muscle fiber cross-sectional area. 42 Recent studies have found that obesity leads to thinning of skeletal muscle capillaries and that this process is regulated by decreased miR-126 levels and increased PI3KR2 expressions. In contrast, exercise training (ET) normalized miR-126 levels and VEGF signal transduction and promoted the proliferation and hypertrophy of muscle fibers. 53
MiRNAs and Muscle Fiber Types
Our understanding of the types of skeletal muscle fibers has evolved over time. Until recent decades, in skeletal muscle, the types of muscle fibers have been defined by 4 isoforms of the adult myosin heavy chain (MyHC). The 4 isoforms of the MyHC gene, namely MyHC-I, MyHC-IIa, MyHC-IIb and MyHC-IIx, are translated into different structural proteins expressed in myofibrils and that subsequently form different types of muscle fibers. 54 Clinically, we often divide muscle fibers into fast and slow muscles. Fast muscle fibers have a larger diameter than slow muscle fibers and contain more contractile proteins. The sarcoplasmic network of fast muscle fibers is also more developed than that of slow muscle fibers; the capillary network around slow muscle fibers is richer than that of fast muscle fibers, and slow muscle fibers contain more muscle hemoglobin, which causes them to usually appear red in color. 55 Compared with fast muscle fibers, slow muscle fibers contain more mitochondria, and their mitochondria are larger in volume. 55 In terms of innervation, slow muscle fibers are innervated by smaller motor neurons with thinner motor nerve fibers and a slower conduction speed, while fast muscle fibers are innervated by larger motor neurons with thicker nerve fibers and a faster conduction speed. 56
Previous studies have clearly demonstrated that muscle fiber is a dynamic structure that can change its phenotype under various conditions. To meet the requirements of the external environment, muscle fiber properties, including size and type, change as a result of age, nutrition, environment and other factors. Changes in the expression of myosin subtypes occurs in an orderly manner according to the “nearest neighbor” rule (i.e., type IIa ↔ type IIx ↔ type IIb). 57 After aerobic exercise, the oxidation capacity of animal skeletal muscle is significantly improved; additionally, aerobic exercise induces the transformation of glycolytic muscle fibers to oxidative muscle fibers (i.e., from type IIb or type IIx to type IIa). 58 Long-term high-intensity intermittent training increases the proportion of type Ⅰ and type Ⅱ muscle fibers in the fast muscle and type Ⅰ muscle fibers in the slow muscle of SD mice. 59
Intriguingly, recent studies have indicated that some miRNAs may be coexpressed with the major histocompatibility complex (MHC). Loss of miR-133a leads to excessive activation of protein kinase B (Akt), which may impair mitochondrial biogenesis and promote type IIb muscle fiber loss. Therefore, miR-133a plays a vital role in muscle function and plasticity. 60 Inactivation of miR-208b and miR-499 causes a significant loss of type I soleus muscle fibers in addition to an increase in sex-determining region Y-box 6 (Sox6) expression and a 60% decrease in MHCII expression. In contrast, overexpression of miR-499 causes all muscle fibers of the soleus muscle to be converted to type I. 61 VanRooij et al 61 showed that miR-208a, miR-208b and miR-499 are intron miRNAs coexpressed with the myosin genes MHC-α, MHC-β and Myh7b. Most importantly, these miRNAs seem to be involved in the regulation of muscle gene expression, fiber size and performance. Recent studies have shown that in C2C12 myotubes and primary cultures, overexpression of miR-151-3p not only increases the proliferation of C2C12 myoblasts but also reduces the expression of slow muscle genes (e.g., MHC-β and muscle troponin I). 62 Overexpression of miR-151 does not seem to affect the myogenic differentiation of C2C12 cells. In contrast, miR-151 inhibits MHC-β and regulates muscle fiber types by directly targeting SERCA2. 62 Peroxisome proliferator-activated receptor (PPAR)β/δ and PPARα, which control the transcription of muscle metabolism genes, also exert an opposing effect on the type I fiber program through miR-499 and miR-208b. 63 Functional studies conducted on mice have shown that stimulation of miR-499 and miR-208b expression by estrogen-related receptor γ (ERRγ) increases the proportion of type I muscle fibers. 64
MiRNAs and MuSCs
Responsible for skeletal muscle regeneration, MuSCs are a type of adult stem cell located between the muscle and basement membrane and are the most important participant in muscle regeneration. 65 In the neonatal period, to accommodate the rapid growth of the body, MuSCs are highly activated and proliferate rapidly. During this period, the number of MuSCs, which account for approximately 30% of the total number of muscle cell nuclei, is at its highest. In adulthood, the number of MuSCs decreases, and the MuSCs usually remain in a resting state. When the muscle is damaged, MuSCs in the resting state are activated, enter the proliferation stage and further differentiate and fuse to form an orderly arrangement of myotubes that fuse to form muscle fibers and, ultimately, to form muscle. 66 With increasing age, the proliferative capacity of MuSCs decreases significantly, which results in impaired muscle regeneration ability in aging individuals. 67 Therefore, improving stem cell function and enhancing skeletal muscle regeneration and postinjury repair are effective ways to treat age-related sarcopenia.
Studies have found that a series of miRNAs participate in the processes of muscle stem cell proliferation and differentiation and promote the activation and proliferation of MuSCs. 6,68,69 MiR-195, miR-489 and miR-497 control the resting state of skeletal MuSCs by inhibiting cell division cycle 25A (CDC25A), cyclin D2 (CCND2) and Dek, respectively. By targeting paired box 7 (PAX7), PAX3, myostatin and serum response factor (SRF), several miRNAs, including miR-1, miR-27, miR-133, miR-206 and miR-486, affect the initiation of myogenic processes. Recently, a specific example of this relationship between a myogenic regulatory gene and a miRNA was confirmed for PAX3, which has a miR-206 binding site in its 3′UTR. 70 PAX3 is transiently expressed during the activation of MuSCs and regulates the entry of muscle satellite cells into myogenesis. Studies have confirmed that both resistance and aerobic exercise activate MuSCs, potentially due to the microstructural damage muscle fibers sustain during exercise, and induce the release of inflammatory substances and growth factors from working muscle and the surrounding connective tissue. 71 Among these studies, many have focused on the ability of resistance exercise to promote the activation and proliferation of MuSCs and increase muscle volume (mainly fast muscle fibers), and the effect of resistance exercise on muscle strength is obvious. 72 Aerobic exercise has little effect on muscle mass and can activate MuSCs. The main benefits of aerobic exercise include increasing mitochondrial function, improving cardiopulmonary function, reducing muscle fiber apoptosis, and maintaining the number of muscle fibers. 19
Summary and Prospects
Progressive age-related changes in skeletal muscle mass and composition are the basis for the decline of muscle function in elderly individuals, which may lead to impaired mobility and decreased quality of life. MiRNAs are important posttranscriptional regulators of gene expression in skeletal muscle and are related to aging. There is increasing evidence that miRNAs play a substantial role in age-related changes in skeletal muscle mass, composition and function. At the cellular level, it has been shown that miRNAs regulate the proliferation and differentiation of muscle cells. In addition, miRNAs are also involved in the transition of MuSCs from a resting state to an activated or senescent state. 6 Evidence from animal and human studies suggests that miRNAs are regulated by muscle atrophy and hypertrophy. In addition, miRNAs are associated with changes in muscle fiber composition, fat infiltration and insulin resistance.
Skeletal muscle is the most important organ related to exercise. Exercise-mediated regulation of gene expression in skeletal muscle is not only critical to the body’s ability to adapt to exercise and perform the activities of daily life but also required to reduce disease risk. Different exercise patterns induce various molecular mechanisms of adaptation, including the activation of specific signaling pathways and related genes, and miRNAs are important factors that affect gene expression. MiRNAs play an essential role in various physiological processes, including the initial stress response of cells; because miRNAs can quickly respond to various stimuli, they are ideal candidate molecules for studying the mechanism of exercise adaptation. At present, some studies have made progress in uncovering the epigenetic mechanisms regulating miRNA levels, such as the mechanism by which exercise regulates miRNAs in skeletal muscle. However, the research on mechanisms of miRNA regulation has mainly consisted of studies on animal models and cultured cells, and their findings need to be validated in human studies. In addition, although it has been confirmed that the miRNAs network is involved in the adaptive mechanism of skeletal muscle after exercise, the specific mechanism regulating the expression of miRNAs is not clear. More scientific research and clinical trials are needed to explore the specific effects of miRNAs-mediated different types of exercise in delaying sarcopenia in the elderly. These studies can ultimately provide extremely useful information for the formulation of exercise methods, and are also of great significance for improving the quality of life of patients and reducing national medical expenditures. Regulating the level of miRNAs in skeletal muscle is expected to become a promising and effective intervention strategy to delay the occurrence and development of sarcopenia in the elderly.
Footnotes
Authors’ Note
Wen-Qing Xie, Chen Men, and Miao He contributed equally to this work. MenChenis now affiliated with Department of Geriatric Cardiology, The First Affiliated Hospital of Nanjing Medical University, Nanjing, Jiangsu, China.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (No. 81874030 and 81871096), the Natural Science Foundation of Hunan Province(2020JJ3060), Innovation-Driven Project of Central South university (No.2020CX045), Wu Jieping Medical Foundation (320.6750.2020-03-14), and CMA Young and Middle-aged Doctors Outstanding Development Program—Osteoporosis Specialized Scientific Research Fund Project (G-X-2019-1107-12).