
Abstract
Dose rate is one of the most varied experimental parameters in radiation biology research. In this study, effects of dose rates on the radiation responses of 2 different types of human epithelium-derived cells, immortalized keratinocytes (HaCaT), and colorectal cancer cells (HCT116 p53+/+ and HCT116 p53−/−) were systematically studied. Cells were γ-irradiated at one of the 4 dose rates (24.6, 109, 564, and 1168 mGy/min) to a total dose of 0.5 to 2 Gy. Clonogenic survival and mitochondrial membrane potential (MMP) were measured to assess the levels of reproductive cell death and damage to mitochondrial physiology, respectively. It was found that clonogenic survival was similar at all 4 tested dose rates in the 3 cell lines. The loss of MMP occurred at all tested dose rates in all 3 cell lines except for one case where the MMP increased in HCT116 p53+/+cells after exposure to 0.5 Gy at 24.6 mGy/min. In HCT116 cells, the loss of MMP was the most severe at high dose/dose rate combination exposure and when p53 was expressed. In contrast, no effect in dose rate was observed with HaCaT cells as the reduction level of MMP was similar at the tested dose rates.
Introduction
It is broadly agreed that cellular or organismal responses to ionizing radiation (IR) are influenced not only by the total absorbed dose but also by the dose rate. The consensus position is that the higher the dose rate, the more detrimental the effects. 1 -3 However, fascinatingly, recent research by Montay-Gruel et al (2017) showed that very high dose rates of greater than 100 Gy/s appear to reduce brain toxicity and impaired spatial memory that otherwise usually occur following whole brain irradiation at a lower dose rate (0.1 Gy/s). 4 In the context of dose in radiobiology, IR can induce hormetic mechanisms at low doses 5 ; however, little is known regarding the influence of low dose rate in hormesis. While studies continue to report and focus on the total dose, the inclusion or consideration of dose rate is less ubiquitous. In fact, dose rate remains one of the most varied experimental parameters in radiation biology research. The availability and the original activity of a given radiation source, the applied distance between the source and the irradiated target, and the different radioactive decay rate of the used radionuclides are among the factors making dose rate such a challenge to control for experimentations. In addition, in studies where both very low and very high doses are tested (eg, to study the radio-adaptive response), the very low doses are usually achieved at a low dose rate and the very high doses at a high dose rate. Thus, this makes it difficult to know if the reported effects are due to only dose or both dose and dose rate.
As well as damaging deoxyribonucleic acids in the cell nucleus, IR can also impose direct damage to the mitochondria without direct nuclear exposure. 6 Following IR, endogenous oxidative stress increases, the mitochondria depolarize, the mitochondrial membrane transition pores are formed, cytochrome c is released, and cell death programs are activated. 7 Therefore, understanding the influence of IR on the mitochondrial health is important. In fact, in oxygenated tissues, mitochondrial stress dictates the radiosensitivity. 8 Although the radiation effects on the mitochondrial biology to date is mostly restricted to the principle of total absorbed dose, the influence of dose rate in this regard is rarely examined.
The tumor suppressor p53 is one of the most important proteins in orchestrating cellular responses to radiation insults. Upon irradiation, p53 is activated via post-translational modification by different groups of enzymes including kinases and histone acetyltransferases. 9 Activated p53 has both transcription-dependent and -independent functions. As a transcriptional factor, activated p53 transactivates different target genes, many of which are important in cell cycle regulation, DNA repair, and cell death programs. 10 Activated p53 also regulates the mitochondrial membrane potential (MMP) and initiates the cell death programs independent of its transcriptional activity. 7,11 In cells that lack functional p53, loss of the MMP and other cellular consequences take place via p53-independent mechanisms. 12 Cells that have mutated p53 or are deficient in functional p53 tend to have lower MMP than do cells with wild-type p53. 13,14
In this study, we sought to study how dose rate affects the clonogenic cell survival (as an end point for the damage leading to reproductive cell death) and the MMP (as a measure for the damage level to the mitochondrial physiology) in different types of human epithelium-derived cells: spontaneously immortalized keratinocytes and colorectal carcinoma cells. The influence of functional p53 on the dose rate effects was studied in the colorectal carcinoma cells.
Materials and Methods
Cell Lines
Three immortalized human epithelium-derived cell lines were used in this study. They were HaCaT, HCT116 p53+/+, and HCT116 p53−/−. HaCaT is a keratinocyte cell line derived from a normal non-tumorigenic skin biopsy from a patient with a documented melanoma. 15 HCT116 p53+/+ (p53 wild-type or p53 wt) is a colorectal carcinoma cell line with a wild-type p53 gene. 16 HCT116 p53−/− (p53 null) is derived from the HCT 116 p53+/+ cell line and has both of its p53 alleles inactivated via the homologous recombination technology. 17 The HaCaT and HCT116 p53 variant cell lines were kindly provided by Dr Petra Boukamp (German Cancer Research Center, Heidelberg, Germany) and Dr Robert Bristow (Princess Margaret Hospital & University of Toronto, Toronto, Ontario).
All 3 cell lines were maintained in the common growth media made up of the basal Roswell Park Memorial Institute (RPMI) medium (Gibco, Thermo-Fisher Scientific), 10% fetal bovine serum (FBS, PAA), 100 U/mL penicillin, 100 μg/mL streptomycin, 25 mM HEPES, and 2.05 mM
Irradiation
The γ radiation source was 37 TBq (or 1 kCi) 137Cs located in the Taylor Radiobiology research facility at McMaster University (Hamilton, Ontario). The γ source was calibrated by health physicists at McMaster University. Thermoluminescent dosimeters were used to measure radiation doses. Irradiated flasks were placed at a certain source-to-surface distance (SSD) to acquire specific doses at specific dose rates as previously demonstrated. 20 In this work, cells were irradiated at 2 doses (0.5 and 2 Gy) and 4 dose rates were used (24.6, 109, 564, and 1168 mGy/min). The lowest dose rate was achieved by placing the flasks 90 cm away from the source and using one shielding lead filter of 0.31 cm thickness. The other dose rates were achieved with no lead filters at the SSDs of 12, 18, and 45 cm. Irradiation was performed at room temperature, and the cell culture flasks were immediately returned to the 37°C incubator following irradiation.
The lowest dose rate of 24.6 mGy/min was in a range that would be considered low in radiation oncology and cellular radiobiology experiments. 21 -24 However, 24.6 mGy/min is relatively higher than both the environmental contamination/occupational exposure levels associated with radioactive fallout accidents 25 -28 and the UNSCEAR’s recommended low dose rate of <0.1 mGy/min (UNSCEAR 2012). 29 The other 3 dose rates used here would generally be considered high. 21,23,24,30
Measuring the Level of Reproductive Cell Death by Clonogenic Assays
Following the Puck and Marcus method, 31 400 cells were seeded into triplicate 25-cm2 flasks containing 5 mL of fresh growth media. After incubation for 6 hours at 37°C, cells were irradiated with either 0.5 or 2 Gy of γ rays at one of the 4 dose rates (24.6, 109, 564, and 1168 mGy/min). Sham- and γ-irradiated flasks were returned to the 37°C incubator immediately after irradiation. After 9 days, macroscopic surviving colonies (having at least 50 cells) were stained with the carbol fushc in solution (Ricca Chemical Company, Arlington, Texas) diluted at 1:4 (v/v) in water. The numbers of survivor colonies were counted, and the clonogenic survival fraction was determined as previously demonstrated. 18 -20 Three to 4 independent experiments were performed.
Quantitative Measurement of the Relative MMP by the Fluorescent Probe JC-1
The relative MMP (rMMP) was measured using a highly sensitive potentiometric and ratiometric fluorescent dye JC-1 (5,5′,6,6′-tetrachloro-1,1′3,3′-tetraethylbenzamidazol-carboncyanine). JC-1 is mitochondria-bound upon crossing the plasma membrane. Under normal conditions, the mitochondrial potential gradient converts JC-1 monomers, which fluoresce green, to the J-aggregates, which fluoresce orange. When the mitochondria membrane depolarizes and the mitochondrial potential decreases or collapses due to, for example, oxidative stress or apoptosis, J-aggregates disintegrate back into the JC-1 monomers, resulting in a lower ratio of orange/green fluorescence.
As previously demonstrated, 18,20 we used the quantitative approach for cultured cells using the multi-well microplates, an automated cytofluorometric plate reader, and the MitoPT JC-1 Mitochondrial Permeability Transition Kit (ImmunoChemistry Technologies, Bloomington, Minnesota). Briefly, 1.5 million sham or irradiated cells were incubated with 500 μL of JC-1 working solution (prepared as recommended by the manufacturer) in the 37°C water bath for 20 minutes. JC-1 dye was added after irradiation. Cell pellets were rinsed with DPBS once and then resuspended in 1 mL of fresh DPBS. In a CellStar 96-well black tissue culture polystyrene microplate with μClear round chimney bottom (Greiner Bio-One), 100 μL containing 1.5 × 105 cells was added into 6-well replicates. Carbonyl cyanide m-chlorophenylhydrazone (CCCP) was used as a positive control. Fluorescence was measured with the 485 ± 9 nm excitation and dual emission wavelengths of 535 ± 20 nm for green fluorescence and 590 ± 20 nm for orange fluorescence using the Tecan Infinite M200 Pro microplate reader with integrated command via the i-control software and data acquisition via Microsoft Excel software at the BioInterfaces Institute (McMaster University, Hamilton, Ontario). Numerical data were recorded in relative fluorescence units (RFUs). Ratios of orange RFUs to green RFUs were calculated, representing relative levels of MMP. All MMP ratios were normalized to those of the control cells to yield the rMMP to the control. The mitochondria were deemed to be depolarized when the rMMP ratios were lower than 1.0.
The rMMP was measured in cells following exposure to γ rays or treatment with the mitochondrial depolarization-inducing protonophore CCCP. For γ radiation exposure, 5 × 105cells were seeded per 25-cm2 flask for 24 hours and irradiated with 0.5 Gy or 2 Gy at one of 2 dose rates (24.6 and 1168 mGy/min).Sham-irradiated control flasks received no radiation. Both irradiated and sham-irradiated flasks were immediately returned to the 37°C incubator following irradiation and kept at 37°C for 2 hours prior to MMP analysis with JC-1. Two hours post irradiation was previously found effective to detect dose-dependent mitochondrial depolarization in HCT116 and HaCaT cells. 18 For CCCP treatment, 1.5 × 106 cells were evenly divided into 4 15-mL conical tubes. After the 267 g centrifugation for 4 minutes, cell pellets were resuspended in 1 mL of the complete growth medium containing the vehicle control (0.1% DMSO) or 5 μM CCCP. The tubes were incubated at 37°C with 5% CO2 for 60 minutes, followed by MMP measurement by JC-1. For both radiation and CCCP exposure, 3 independent experiments were performed.
Statistical Analyses
A 1-way analysis of variance test with the Tukey multiple comparisons post-test with a 95% confidence interval and a Student t test following a distribution normality check based on the Mann-Whitney test were 2 statistical tests used wherever appropriate. The statistical analyses were performed with the GraphPad Prism 6 software, and the differences were deemed statistically significant when P < .05.
Results
Effects of Dose Rates on the Clonogenic Survival and rMMP in HaCaT Cells
Two doses, 0.5 and 2 Gy, and 4 dose rates, 24.6, 109, 564, and 1168 mGy/min, were tested. In HaCaT cells, 2 Gy reduced clonogenic survival more than did 0.5 Gy (Figure 1A). Clonogenic survival was similar in all tested dose rates (Figure 1A). Since there was no significant difference in clonogenic survival due to different dose rates, only the lowest (24.6 mGy/min) and the highest (1168 mGy/min) dose rates were considered for the MMP studies. The rMMP was not statistically significantly different in both tested doses and dose rates (Figure 1B). The result shows that at 0.5 to 2 Gy dose rates ranging between 24.6 and 1168 mGy/min led to similar reductions in the clonogenic survival and the MMP in immortalized human keratinocytes.

Effects of dose rate on the clonogenic survival and relative mitochondrial membrane potential (rMMP) in the human keratinocyte HaCaT cell line. Gamma irradiation was performed at 24.6, 109, 564, or 1168 mGy/min. Cells received a total dose of 0.5 or 2 Gy. (A) Clonogenic survival fractions were presented as mean ± SEM from 3 independent experiments. (B) The rMMP was measured with the fluorescent probe JC-1 and data were normalized to the nonirradiated control and presented as mean ± SEM from 3 independent experiments. The 1-way ANOVA test along with the Tukey multiple comparisons test with a 95% confidence interval was used for statistical analysis. P < .05 is deemed significant. Groups with the same letter are not statistically different from one another whereas those with different letters are statistically different. ANOVA indicates analysis of variance.
Effects of Dose Rates on the Clonogenic Survival and rMMP in HCT116 p53 wt and Null Cells
In both p53 wt and null cell lines, 2 Gy reduced clonogenic survival more than 0.5 Gy (Figure 2). At each dose and in each cell line, clonogenic survival was not statistically significantly different at all tested dose rates (Figure 2). This result shows that at 0.5 to 2 Gy dose rates ranging between 24.6 and 1168 mGy/min led to similar reductions in the clonogenic survival in human colorectal cancer cells. The influence of p53 was further examined by comparing the clonogenic survival between HCT116 p53 wild-type and null cells for each dose and dose rate combination using Student t tests. It was found that there was no statistically significant difference between the wild type and null cells (P > .05).
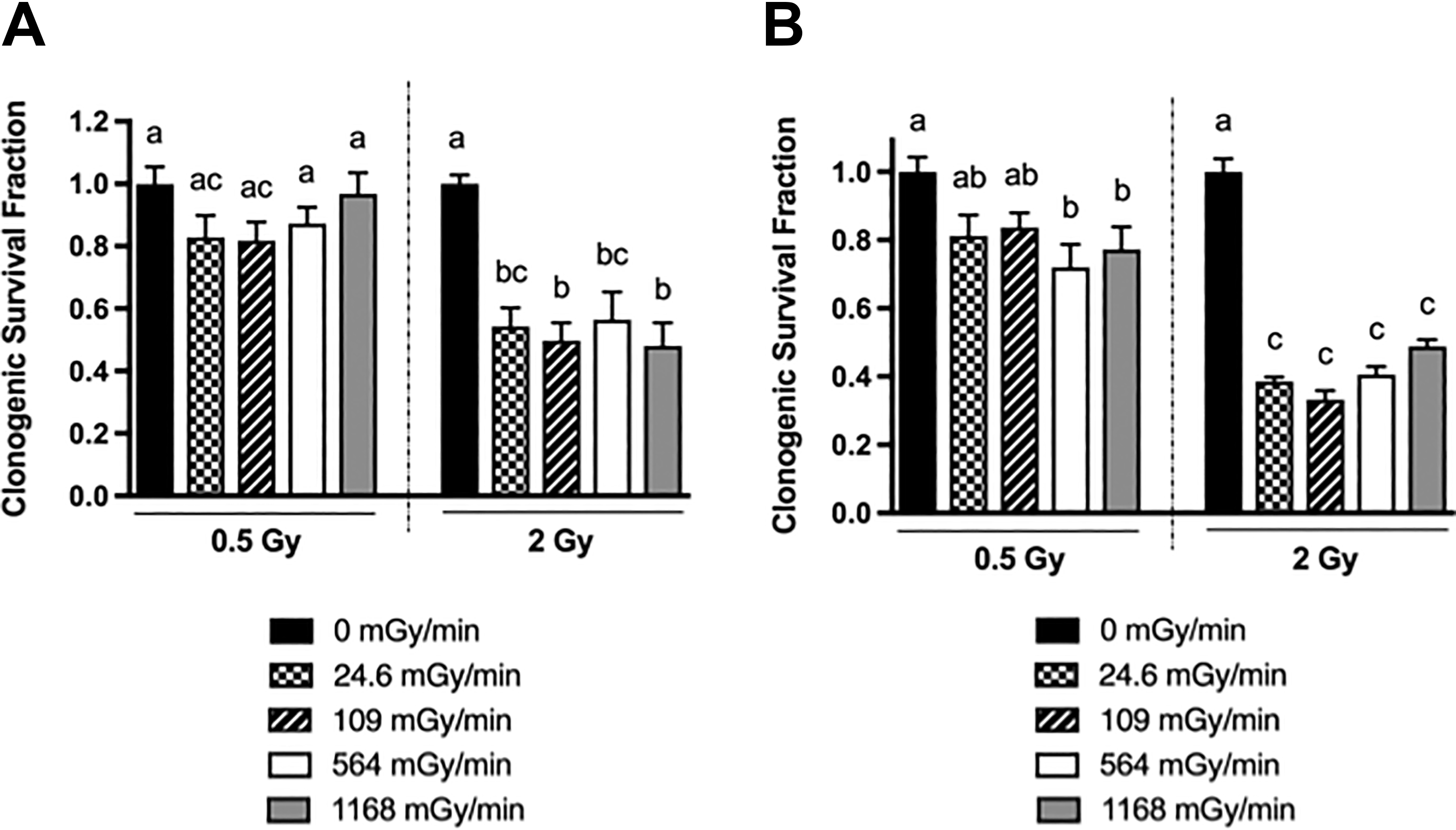
Effects of dose rate on the clonogenic survival of the human colorectal cancer HCT116 p53+/+ (A) and p53−/− (B) cell lines. Gamma irradiation was performed at 24.6, 109, 564, or 1168 mGy/min. Cells received a total dose of 0.5 or 2 Gy. Clonogenic survival fractions were presented as mean ± SEM from 3-4 independent experiments. The 1-way ANOVA test along with the Tukey multiple comparisons test with a 95% confidence interval was used for statistical analysis. P < .05 is deemed significant. Groups with the same letter are not statistically different from one another whereas those with different letters are statistically different. ANOVA indicates analysis of variance.
Two doses (low at 0.5 Gy and high at 2 Gy) and 2 dose rates (the lowest at 24.6 mGy/min and the highest at 1168 mGy/min) gave 4 radiation conditions: low dose/low dose rate (L2), low dose/high-dose rate (LD/HDR), high dose/low-dose rate (HD/LDR), and high dose/high-dose rate (H2). In the HCT116 p53 wt cells, the rMMP increased in the L2 condition but decreased in LD/HDR and both high-dose conditions (Figure 3A). At both 0.5 and 2 Gy doses, the higher dose rate decreased the MMP more significantly than did the lower dose rate (Figure 3A). Interestingly, the rMMP level in HD/LDR was similar to that in LD/HDR but was significantly higher than that in H2 (Figure 3A). Thus, in HCT116 p53 wt cells, the MMP was dependent on both dose and dose rate.

Effects of dose rate on the rMMP in the human colorectal cancer HCT116 p53+/+ (A) and p53−/− (B) cell lines. Gamma irradiation was performed at 24.6 or 1168 mGy/min. Cells received a total dose of 0.5 or 2 Gy. The rMMP was measured with the fluorescent probe JC-1 and data were normalized to the non-irradiated control and presented as mean ± SEM from 3 independent experiments. The 1-way ANOVA test along with the Tukey multiple comparisons test with a 95% confidence interval was used for statistical analysis. P < .05 is deemed significant. Groups with the same letter are not statistically different from one another whereas those with different letters are statistically different. ANOVA indicates analysis of variance.
In HCT116 p53 null cells, the rMMP decreased in all of 4 radiation conditions (Figure 3B). At 0.5 Gy, the rMMP was similar at both dose rates (Figure 3B). However, at 2 Gy, the higher dose rate depolarized the mitochondria more than did the lower dose rate (Figure 3B), which was the same trend as in the HCT116 p53 wild-type cells (Figure 3A). Interestingly, the rMMP in the HD/LDR was similar to those in both low-dose conditions. Thus, in HCT116 p53 null cells, the dual combination of high dose and high dose rate played a bigger impact on the degree of mitochondrial depolarization. Nevertheless, the loss of MMP in p53-deficient HCT116 cells further showed that p53 is not a sole determinant regulating the mitochondrial depolarization leading to cell death.
The influence of p53 on the MMP was studied by comparing the responses between HCT116 p53 wt and null cells in each radiation condition. In L2, the rMMP in the p53 wild-type cells was significantly higher than p53 null cells (P = .0001, Student t test). In LD/HDR, the rMMP was similar in both cell populations (P = .8426, Student t test). However, in both high-dose conditions, the rMMP in the p53 wild-type cells was significantly lower than their p53 null counterparts (P < .001, Student t test). Our analysis shows that when p53 is absent, the loss of MMP is more serious at low dose/dose rate but alleviated at high dose.
Effect of CCCP on the rMMP in HCT116 p53 wt and null Cells
To understand further whether or not influence of p53 on the mitochondrial depolarization state might be an exclusive mechanism in response to radiation, CCCP, which is a mitochondrial depolarizer, was used and the degree of mitochondrial depolarization was assessed. The rMMP was significantly higher in the p53 wt cells than in the p53 null cells (Figure 4). Therefore, in contrast to the radiation stimulus (Figure 4), mitochondria in cells without p53 were more prone to the CCCP’s mitochondria-depolarizing action than those in cells with functional p53. Collectively, our data showed that the p53 response to loss of MMP is not programmed via a general mechanism and is tailored to a specific stressor, such as radiation in this case.
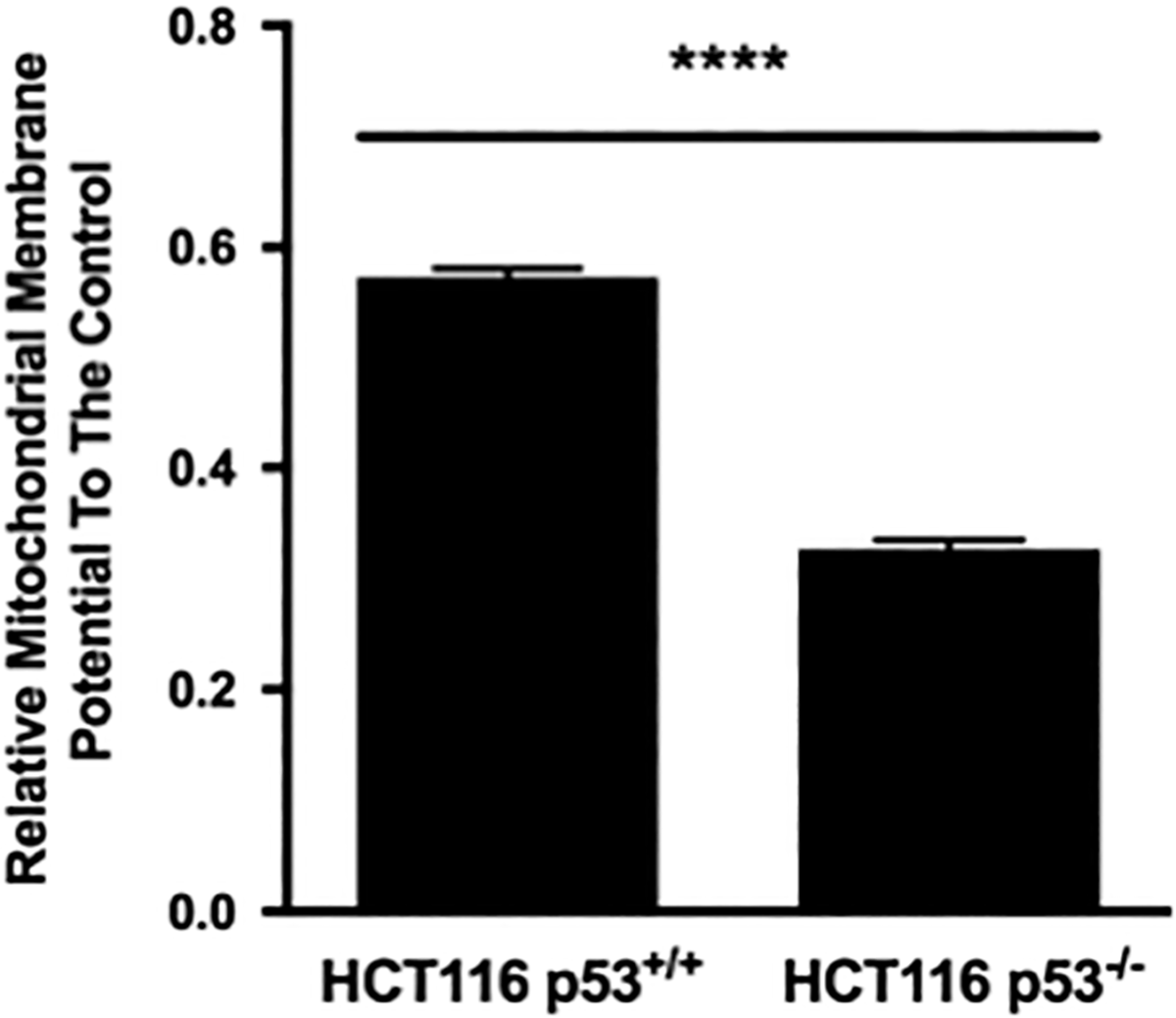
Effects of the mitochondria-depolarizing protonophore carbonyl cyanide m-chlorophenylhydrazone (CCCP) on the rMMP of the HCT116 p53+/+ and p53−/− cell lines. Cells were treated with 5 μM CCCP for 60 minutes. The vehicle control cells received 0.1% DMSO. The rMMP was assessed by the JC-1 probe. Data were normalized to the vehicle control and presented as mean ± SEM from 3 independent experiments. A Student t test based on the Mann-Whitney test was performed for statistical analysis. ****P < .0001.
Discussion
In this study, for the dose rate range of 24.6 to 1168 mGy/min, we found that there was no major dose rate effect in the clonogenic survival of HaCaT and HCT116 cells following γ radiation. In the view of the reproductive cell death, the dose rate effect appears to be cell type dependent. Similar observations were previously made with human neuroblastoma cells receiving γ rays at a dose rate range of 10 to 900 mGy/min; however, a lower dose rate of 2.5 mGy/min was found to have a less deleterious effect. 22 For human normal dermal fibroblasts and melanoma cells following X-ray irradiation, 1120 mGy/min was much more damaging than 8.8 mGy/min. 23 Enn et al (2004) found that there was no major dose rate effect (1.8 vs 220 mGy/min) in human lung adenocarcinoma cells A549. However, the same group also found that the dose rate of 1.8 mGy/min was less damaging to human glioblastoma cells T98G than 220 mGy/min at only 1 dose point (1 Gy). 30
Mitochondrial depolarization is one of the central mitochondrial events as a result of IR. 7 The most serious consequence of mitochondrial depolarization is the downstream activation of signaling cascades leading to cell death. Loss of MMP could occur at both low and high doses 18,20,32 and at a dose rate of as low as 0.66 μG/d (following chronic alpha particle irradiation). 18 Recently, upon γ radiation, both HaCaT and HCT116 p53 wild-type cells were shown to experience the loss of MMP at a low dose of 0.04 Gy at a dose rate of 148 mGy/min. 18 Thus, the current results with high dose rate were consistent with the previous work.
In this study, we found that the loss of MMP occurred at all tested dose rates except for one case where the MMP increased in HCT116 p53 wild-type cells after exposure to 0.5 to Gy γ rays at a dose rate 24.6 mGy/min. Elevation of the MMP following exposure to a potentially lethal stressor including radiation has been seen previously and appears to be restricted to a small set of cell phenotypes. 20,33 -36 The elevation of MMP is usually coupled with increased oxidative stress, which occurs upon irradiation, 33,37 develops transiently, and always precedes the mitochondrial depolarization event. 34,35 This would be a logical explanation for the observed elevated MMP in HCT116 p53 wild-type cells after exposure to 0.5-Gy γ rays at a dose rate 24.6 mGy/min and it is logical to anticipate that the mitochondrial depolarization would be the next step to follow. Interestingly, p53 is intimately involved in the early development of MMP elevation. 33 In HCT116 p53 null cells that do not express p53, for the same exposure regimen (0.5-Gy, 24.6 mGy/min), the MMP elevation was not observed and only the loss of MMP was observed.
The role of p53 appears to be more significant in its contribution to the loss of MMP at high dose as the lack of p53 relieves this tension (MMP is higher). This observation makes sense as radiation-induced activated p53 can influence the loss of MMP in 2 ways: the first being transactivating p53 target genes that are involved in mitochondrial signaling such as those of pro-apoptotic nature 6 and the second being directly translocating into the mitochondria and initiating the loss of MMP prior to p53’s transactivation of genes in the nucleus. 7,11 As a result, p53 would add an additional wave of biochemical signals amplifying the effect. Our observation is in agreement with Charlot et al (2004), who showed that cells with wild-type or mutated p53 experienced a larger loss of MMP after apoptosis induction than did p53-null cells.
Overall, this study is one of a few systematically looking at the dose-rate radiobiology in human epithelium-derived cells. Dose rates ranging from 24.6 to 1168 mGy/min had little effects on the clonogenic survival reduction in HaCaT and HCT116 cells. With respect to the MMP, different dose rates induced different degrees of mitochondrial polarization in HCT116 cells but not HaCaT cells. The dose rate effect on the MMP is more pronounced when wild-typep53 is expressed. Why there was no dose-rate effect on the MMP in HaCaT cells requires future investigations. One explanation could be that HaCaT cells are more resilient in coping with oxidative stress than are HCT116 cells. It is well known that IR elevates the level of reactive oxygen species (ROS) that depolarize mitochondria and damage cellular functions. HaCaT cells have a much higher basal level of endogenous intracellular reactive oxygen species than do HCT116 cells, 38 which could enable HaCaT cells to be more adaptive and confer a higher level of resistance toward the ROS imbalance. In fact, in a low-dose ultraviolet radiation challenge, ROS levels in HaCaT cells were statistically indifferent at different dose rates. 39 As the HCT116 and HaCaT cell lines are common cell models in radiation biology and the dose rates we used were within the dose-rate range used by many researchers, our dose rate findings would be of interest to many radiation researchers in the field.
Footnotes
Acknowledgments
The authors thank Scott McMaster and the Health Physics department at McMaster University for calibrating the Taylor gamma irradiation source. The authors are grateful to Dr Robert Bristow (Princess Margaret Hospital & University of Toronto, Toronto, ON) for the gifts of the 2 HCT116 cell lines and to Dr Petra Boukamp (German Cancer Research Center, Heidelberg, Germany) for the gift of the HaCaT cell line.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article:This research was supported by the Canada Research Chairs program to CEM (grant number 950-221284) and the Natural Sciences and Engineering Research Council (NSERC) of Canada in the form of an Industrial Research Chair grant to CBS (grant number RGPIN293153-12). This research was also supported by am NSERC Collaborative Research and Development grant to CEM and CBS (grant number CRDPJ484381-15) in an industrial partnership with CANDU Owners Group and Bruce Power.