
Abstract
The unsatisfactory clinical efficacy of dendritic cell (DC)-based cancer vaccines prepared by conventional methods is partly due to their insufficient capacity for migration. Our previous study showed that exposure to low-dose radiation (LDR) at a dose of 0.2 Gy promoted DC migration in vitro. The present study further investigates whether exposure to LDR at a dose of 0.2 Gy during the DC vaccine preparation could increase the antitumor effect of DC vaccines derived from mouse bone marrow. Our results showed that the migratory capacities of DCs were significantly increased after exposure to LDR. Furthermore, exposure to LDR resulted in an increased ability of DCs to induce T-cell proliferation, and the cytotoxic effect of cytotoxic T lymphocytes (CTLs) primed by the DCs exposed to LDR was significantly enhanced. An in vivo study using a mouse transplanted tumor model showed that subcutaneous injections of a DC vaccine exposed to LDR led to an increased mouse survival rate, infiltration of CTLs into tumor tissue, and apoptosis of tumor cells, which were accompanied by significant upregulation of serum interferon γ and interleukin 12. These results indicate that exposing DCs to LDR during the DC vaccine preparation is an effective approach to enhance its antitumor effect.
Introduction
Dendritic cells (DCs) are the most important antigen-presenting cells due to their “professional” and extremely efficient antigen-presenting function. The DCs that populate nonlymphoid tissue capture the antigens derived from malignant, infected, and allogeneic cells and subsequently process the captured antigens into the form of peptide–major histocompatibility complex molecule complexes. After migration into secondary lymphoid organs, such as draining lymph nodes (LNs), the antigen-loaded DCs will be present to naive CD8+ T cells, giving rise to cytotoxic T lymphocytes (CTLs), as well as to naive CD4+ T cells, contributing to the differentiation of B cells into antibody-secreting cells. 1 Effector CTLs play a central role in host defense by killing malignant or infected target cells through various mechanisms. 2 However, unlike infected cells, malignant cells generally induce a weak and even ineffective immune response against cancers due to the poor activation of DCs resulting from the poor immunogenicity of tumor cells 3 or the inhibitory effect of cytokines that are secreted by cancer cells in the tumor microenvironment. 4,5
Therapeutic cancer vaccination based on DCs was developed to activate DCs using tumor antigens in vitro, which can prime naive T cells and subsequently overcome the weak immune response against cancer. Currently, ex vivo therapeutic DCs are usually generated by the in vitro stimulation of peripheral blood mononuclear cells (PBMCs) with a cytokine cocktail consisting of granulocyte–macrophage colony-stimulating factor (GM-CSF) and interleukin (IL)-4, followed by tumor antigen loading and inflammatory signal stimuli, such as lipopolysaccharide or tumor necrosis factor (TNF), which contribute to the maturation of DCs. 6,7 The practical implementation of DC-based cancer therapy has shown some significant clinical outcomes in several types of cancer. 8,9 However, only marginally clinical benefits were observed in most types of cancer due to the low capacity of the vaccines to generate tumor-specific T-cell responses. 10,11 A clinical study has suggested that no more than 5% of mature DCs can reach draining LNs after intradermal injections, 12 which was further confirmed by another study performed by Verdijk et al. 13 These results suggest that the ineffective migration of DCs to the LNs could, to some extent, be responsible for the low capacity of the vaccines to generate tumor-specific T-cell responses.
It has been reported that low-dose radiation (LDR) can induce a hormesis effect on the immunological network in both animal and human populations through enhancing the activity of T cells and natural killer (NK) cells. 14 –17 Furthermore, our previous study found that a single exposure of DCs that were derived from human PBMCs to LDR at a dose of 0.2 Gy significantly promoted the production of IL-12, which plays an important role in the differentiation of naive T cells into Th1 cells and the activation of CD8+ CTLs, 18 –20 and enhanced the ability of the DCs to induce T-cell proliferation. 21 In addition, our study showed that the migratory ability of DCs derived from human PBMCs also significantly increased after exposure to LDR at a dose of 0.2 Gy. 22 These results indicate that LDR can also enhance the function of DCs, which indicates that exposure to LDR may be a promising approach to improve the outcomes of DC-based cancer vaccines by enhancing the migration of DCs, as well as IL-12 secretion.
In the present study, we investigated the effect of a single exposure to LDR at a dose of 0.2 Gy on the migration of DCs derived from mice bone marrow and further examined whether LDR could enhance the in vitro and in vivo antitumor effects of DC-based cancer vaccines.
Material and Methods
Cell Lines
The Lewis lung carcinoma (LLC1) cells were obtained from Dr Cheng Wu (The Second Artillery General Hospital) as a gift and maintained in Dulbecco’s modified Eagle medium (DMEM) with 10% fetal bovine serum (FBS).
Mice
Eight-week-old male C57BL/6 (B6) mice were purchased from Vital River Laboratory Animal Technology Co, Ltd (Beijing, China). All animals were maintained in accredited facilities, and the animal protocols were approved by the Animal Care Committee of the Second Artillery General Hospital.
Dendritic Cell Generation, Culture, and Exposure to LDR
Dendritic cells were generated from murine bone marrow cells by Inaba’s protocol 23 with minor modifications. Briefly, bone marrow cells were flushed from the femurs of mice, followed by incubation with red blood cell (RBC) lysing buffer (Sigma-Aldrich, St Louis, Missouri) to deplete the RBCs. The cells were cultured in DMEM supplemented with 10% FBS, 100 U/mL penicillin, 100 mg/mL streptomycin, 8 ng/mL recombinant murine (rm) GM-CSF (PeproTech, Rocky Hill, New Jersey), and 2 ng/mL rmIL-4 (PeproTech). On day 1 of culturing, nonadherent cells were gently removed by gently shaking the dishes, and fresh medium was added. On day 3 of culturing, the LLC1 cell lysate prepared by 5 cycles of freezing and thawing of LLC1 cells were added into cultures, which contained 1 × 106 DCs, to obtain a final concentration of 5 mg/mL protein according to the study by Thumann et al. 24 On day 5 of culturing, the cultures were supplemented with 5 ng/mL recombinant human tumor necrosis factor-α (rmTNF-α) (PeproTech) to induce the maturation of the DCs, which were subsequently exposed to X-ray radiation at a dose of 0.2 Gy the next day. The X-rays were given using a precision linear accelerator (Elekta, Stockholm, Sweden) with a dose rate of 442.89 cGy/min.
Migration Assay of DCs
Dendritic cells were harvested immediately after exposure to LDR. For the in vitro assay, a total of 5 × 104 DCs in 200 μL serum-free DMEM was placed into the upper chamber of Boyden chamber (Corning, New York). Then, chemoattractant medium containing 20% FBS, 20 ng/mL rmCCL19 (PeproTech), and 15 ng/mL rmCCL21 (PeproTech) was placed into the lower chamber of the Boyden chamber. To normalize the migration rates of the DCs, a total of 5 × 104 DCs with or without exposure to LDR in chemoattractant medium were directly seeded into an independent lower chamber of the Boyden chamber to quantify the absorbance of the total cells. The system was incubated at 37°C for 6 hours. After incubation, the DCs in the lower chamber were detected with Cell Counting Kit-8 (CCK-8, KeyGen Biotech, Nanjing, China), according to the manufacturer’s instructions. The absorbance of each well at 450 nm was detected with a microplate reader (Bio-Rad, Hercules, California). The migration rates of the DCs with or without exposure to LDR were shown as the absorbance ratio between the DCs translocated from the upper chamber and the DCs directly seeded into the lower chamber.
For the in vivo migration assay, DCs were stained with 5 μM 5- or 6-(N-succinimidyloxycarbonyl) fluorescein 3´,6´-diacetate (CFSE; Dojindo Molecular Technologies, Kumamoto, Japan) for 20 minutes at 37°C and then washed 3 times with phosphate-buffered saline (PBS) at room temperature. Then, 2 × 106 CFSE-labeled DCs in 100 μL PBS were subsequently injected into the hind footpad of C57BL/6 (B6) mice. Forty-eight hours after injection, the popliteal LNs were removed, mechanically disaggregated, and digested with 1 mg/mL collagenase A to obtain a single-cell suspension, which was analyzed by flow cytometry to identify CFSE-positive cells.
Mixed Lymphocyte Reaction
The mixed lymphocyte reaction (MLR) was performed to assess the ability of the DCs to stimulate T lymphocytes. Briefly, spleens of C57BL/6 (B6) mice were removed and mechanically disaggregated on a 74-μm cell strainer to obtain a single-cell suspension. The single-cell suspension was treated with RBC lysis buffer and passed through a Nylon Wool Fiber column to enrich the T cells. The purified T cells then were co-cultured with DCs at a DC to T cell ratio of 1:10 in round-bottom 48-well microtiter plates for 36 hours. At the end of 36 hours, T-cell proliferation was measured by a CCK-8 assay as described above and expressed as the fold change over the untreated cells.
Detection of the Cytotoxic Effect of CTLs Primed by DCs
T cells isolated from the spleen were co-cultured with DCs and were loaded with whole LLC1 cell lysate at a DC to T cell ratio of 1:10 for 3 days to generate LLC1 cell-specific CTLs, which were subsequently harvested as nonadherent cells. The LLC1 cells (target cells) were seeded into 48-well plates in 0.5 mL culture medium at 2 × 104/well. After static cultivation for 2 hours, 4 × 104 LLC1 cell-specific CTLs (killer cells) were added into the 48-well plates to co-culture with LLC1 cells for 24 hours. At the end of 36 hours, the cell number was detected by a CCK-8 assay as described above. The killing rate of CTLs was calculated by the flowing formula: KR (%) = [1 − (OD(T+K) − ODK)/ODT] × 100% (where KR, killing rate; OD(T+K), the absorbance of the well with both target cells and killer cells; ODK, the absorbance of the well added with killer cells alone; and ODT, the absorbance of the well with target cells alone).
In Vivo Experiment
In this experiment, 5 × 106 of LLC1 cells in 200 µL PBS were subcutaneously injected into the armpit of C57BL/6 (B6) mice. About 2 weeks after injection, the mice were killed, and the tumor masses were surgically resected and cut into tumor blocks that weighted (20 [2]) mg. The tumor blocks were subcutaneously transplanted into the armpit of 8-week-old male C57BL/6 (B6) mice under anesthesia to establish the murine transplantation tumor model of LLC1 cells. The tumor diameters were measured every other day, and each tumor volume in cubic millimeters was calculated by the following formula: V = 0.5 × D × d 2 (where V, volume; D, longitudinal diameter; and d, latitudinal diameter). One week after the tumor challenge, when the tumor volumes reached approximate 600 mm3, the mice were divided into 3 groups (n = 10 mice), namely, the control, the DC, and the DC-LDR groups. For the DC and DC-LDR groups, the mice were subcutaneously injected near the inguinal LN with 1 × 107 DCs or LDR-exposed DCs, respectively, in a total volume of 200 μL PBS twice with a 1-week interval. The mice in the control group were subcutaneously injected with 200 μL PBS in parallel with the mice in the DC and DC-LDR groups. The survival time of the mice was recorded.
For immunohistochemical staining and the detection of cytokines, 15 mice bearing transplanted tumors were divided into 3 groups (n = 5), and they received the treatment twice as described above. Blood from the tail vein was collected on days 7 and 14 after the initial treatment. On day 14 after the initial treatment, all the mice were euthanized, and the tumor tissues were collected.
Enzyme-Linked Immunosorbent Assay
The contents of IL-12 and interferon (IFN)-γ in serum were detected by mouse enzyme-linked immunosorbent assay kits for IL-12 and IFN-γ (Boster Biological Technology Co, Ltd, Wuhan, China), respectively, according to the manufacturer’s instructions.
Immunohistochemical Staining
Tumor tissue was fixed in 10% formalin, embedded in paraffin, and sectioned at 5 µm thickness. The tissue sections were treated with an anti-CD8 antibody (Zhongshanjinqiao Co, Ltd, Beijing, China) at a dilution of 1:100 for 1 hour, followed by incubation with an HRP-conjugated secondary antibody for 1 hour at room temperature. Diaminobenzidine (DAB) was applied to the tissue sections for color development. The sections were subsequently photographed using a microscope, and dark brown staining was considered positive. Positive cells were quantified in a high-power field (×400) using Image-Pro Plus 6.0. The values from sections of at least 3 different tumors were averaged.
Terminal Deoxynucleotidyl Transferase–Mediated dUTP Nick End Labeling Assay
Apoptotic cells in the tumor tissue were detected by terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining according to the manufacturer’s protocol of the in situ cell death detection kit (Roche, Shanghai, China). Briefly, after deparaffinization and rehydration, the sections of tumor tissue were stained for TUNEL with an ApopTag Peroxidase In Situ Apoptosis Detection Kit (Chemicon, Temecula, California), as described in previous studies. Briefly, each slide was deparaffinized and dehydrated and treated with proteinase K (20 mg/L) for 15 minutes at room temperature. The slides were treated with 3% hydrogen peroxide for 5 minutes to quench endogenous peroxidases and then were incubated with a TUNEL reaction mixture containing terminal deoxynucleotidyl transferase and digoxigenin-11-dUTP at 37°C for 1 hour. Then, 3,3×-diaminobenzidine chromogen was applied. Methyl green was used as the counterstain. Positive cells were quantified in a high-power field (×400) using Image-Pro Plus 6.0. The values from sections of at least 3 different tumors were averaged.
Statistical Analysis
The experimental data collected from repeated experiments or multiple animals are presented as the means (standard deviation). The comparisons between different groups were performed by a 2-sided t test using the statistical software SPSS 19.0. For the survival analysis, Kaplan-Meier survival curves were constructed. A Cox proportional hazards model was used, and the hazard ratios between groups were tested by a Wald χ2 test. Differences were considered to be significant at P <.05.
Results
Exposure to X-Ray Radiation at a Dose of 0.2 Gy Promoted the Migration of DCs In Vitro and In Vivo
To investigate whether exposure to LDR could enhance the homing ability of DCs, we first examined the effect of LDR on DC migration in vitro. As shown in Figure 1A, exposure to X-ray radiation at a dose of 0.2 Gy enhanced the migration of DCs toward CCL19 and CCL21. The migration of DCs to LNs was further explored using an in vivo model, as described in the “Methods” section. The results showed that the percentage of CFSE-positive cells in popliteal LNs of mice injected with LDR-exposed DCs significantly increased compared to that of mice injected with unexposed DCs, indicating that exposure to X-ray radiation at a dose of 0.2 Gy promoted the DCs administered by hypodermic injection to migrate to LNs.

Exposure to LDR enhanced the migratory capacity of DCs. The migration of DCs with or without exposure to LDR was examined in vitro by a transwell migration assay combined with a cell number count using a CCK-8 assay (A). After labeling with CFSE, the in vivo migration of DCs with or without exposure to LDR from the injection site (footpad) to the draining LN (popliteal lymph node) was examined using flow cytometry. The representative data of 3 independent experiments are shown (B), followed by a statistical analysis (C). * P < .05 versus untreated. LDR indicates low-dose radiation; DC, dendritic cell; CFSE, 5- or 6-(N-succinimidyloxycarbonyl) fluorescein 3´,6´-diacetate; LN, lymph node.
Exposure to X-Ray Radiation at a Dose of 0.2 Gy Enhanced the T-Cell-Stimulating Activity and Antitumor Effect of DCs In Vitro
After migrating to LN, the T-cell-stimulating capacity of the DCs is critical for the generation of tumor-specific T-cell responses. We, therefore, investigated the effect of LDR on the ability of DCs to stimulate T-cell proliferation using an MLR assay. As shown in Figure 2A, the ability of DCs to induce T-cell proliferation increased significantly after the DCs were exposed to X-ray radiation at a dose of 0.2 Gy. The cytotoxic effect of CTLs was detected to further assess the target cell killing activity mediated by CTLs that were primed by target cell antigen-loaded DCs with or without exposure to LDR. As shown in Figure 2B, the killing rate of CTLs that were generated by DCs exposed to LDR at a dose of 0.2 Gy increased significantly compared to that of CTLs that were generated by DCs without exposure to LDR.

Exposure to LDR enhanced the activity of DCs. A, T cells derived from the spleen of C57BL/6 (B6) mice were added into cultures of DCs and subsequently co-cultured with DCs for 36 hours, followed by counting the number of T cells using a CCK-8 assay. B, The cytotoxicity of CTLs generated by whole LLC1 cell lysate-stimulated DCs with or without exposure to LDR was detected by a CCK-8 assay and expressed as a fold change over untreated cells. *P < .05 versus untreated. LDR indicates low-dose radiation; DC, dendritic cell; CTLs, cytotoxic T lymphocytes; CCK-8, cell counting kit-8.
Administration of a DC Vaccine Exposed to X-Ray Radiation at a Dose of 0.2 Gy Prolonged the Survival Time of Mice Bearing Transplanted Tumors
To investigate whether the LDR-enhanced migration and T-cell-stimulating capacity of DCs could result in increased antitumor effects of DC vaccines in vivo, an LLC1 xenograft model was established by transplanting tumor blocks of LLC1 cells into the armpits of mice. The tumor-bearing mice were subcutaneously injected with DCs twice with a 1-week interval between injections. As shown in Figure 3, the administration of DCs without exposure to LDR seemed to prolong the survival time of the tumor-bearing mice compared to that of mice with the PBS treatment; however, there was no significant difference in survival time after statistical analysis. In contrast, the survival time of tumor-bearing mice that received DCs exposed to LDR was significantly prolonged compared to that of mice with PBS treatment, which indicated that exposure to LDR improved the therapeutic antitumor effect of DC vaccines.
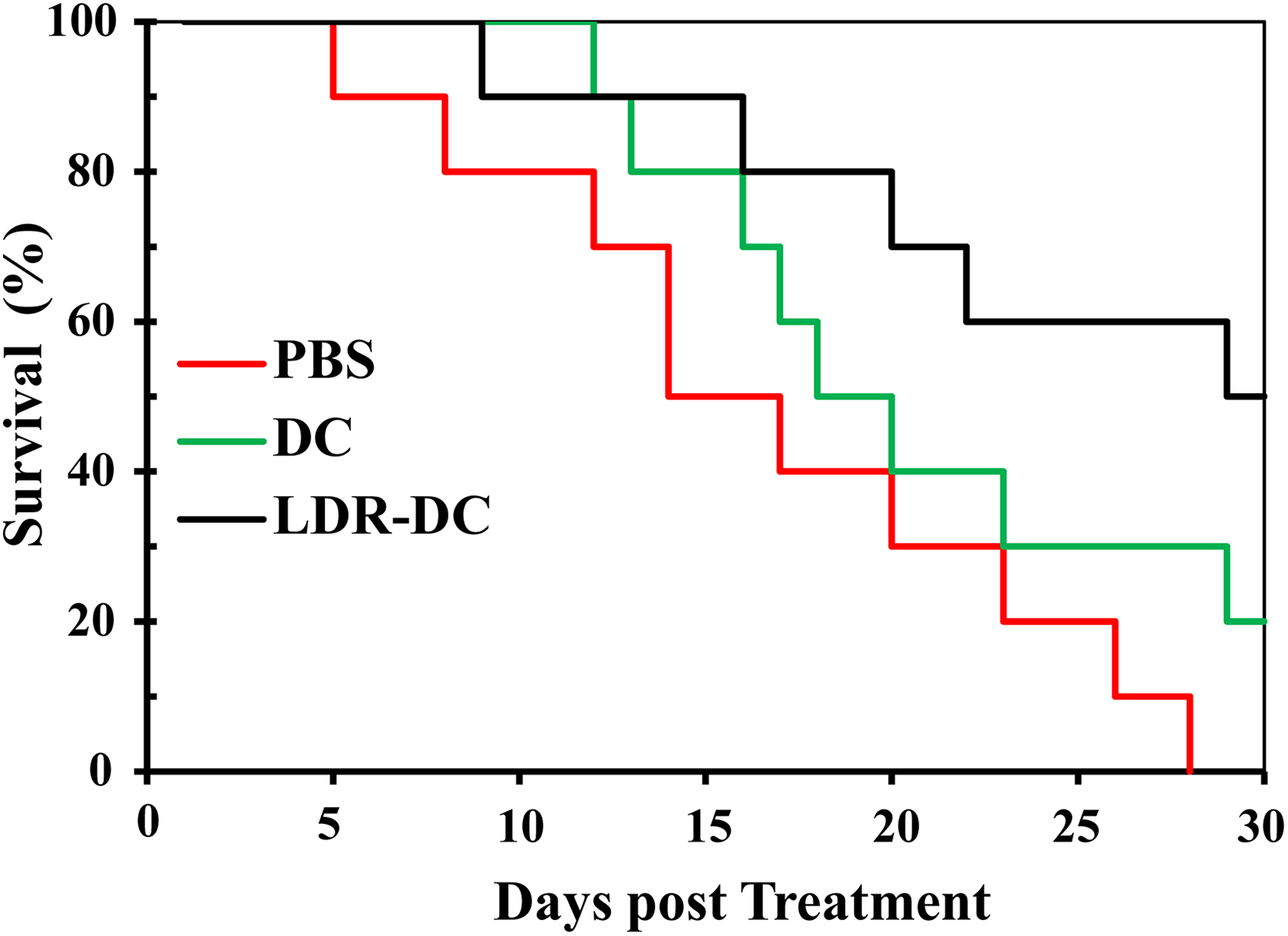
The survival time of mice-bearing transplanted tumors was significantly prolonged by treatment with a DC vaccine exposed to LDR. Mice-bearing transplanted tumors were subcutaneously injected near the inguinal lymph node with PBS, DCs, or LDR-exposed DCs suspended in PBS twice with a 1-week interval between injections. The survival time of the mice was recorded. Kaplan-Meier survival curves were plotted. The difference between LDR-DC and PBS, which was tested by a Wald χ2 test using the Cox proportional hazards model, was statically significant (P = .009), while differences between DC and PBS as well as LDR-DC were not statically significant (P = .169 and .149, respectively). DC indicates dendritic cell; LDR, low-dose radiation; PBS, phosphate-buffered saline.
Administration of a DC Vaccine Exposed to X-Ray Radiation at a Dose of 0.2 Gy Increased IL-12 and IFN-γ in the Serum of Mice
Based on our previous in vitro finding that DCs exposed to LDR at a dose of 0.2 Gy secreted higher levels of IL-12 than did DCs without exposure to LDR, 21 we further investigated whether an injection of a DC vaccine exposed to LDR could also result in increased IL-12 levels in the serum of mice compared with those of mice administered an unexposed DC vaccine injection. As shown in Figure 4A, the serum IL-12 of mice injected with a DC vaccine that was not to LDR increased significantly compared with that of control mice at both 7 and 14 days after treatment. Moreover, an injection of a DC vaccine that was exposed to LDR resulted in a further increase in serum IL-12 at both examined time points compared to that with the treatment with the unexposed DC vaccine.

Treatment with a DC vaccine exposed to LDR resulted in a significant increase in serum IL-12 and IFN-γ in tumor-bearing mice. The serum of mice was collected at both 7 and 14 days after an initial treatment with PBS or a DC vaccine with or without exposure to LDR. The contents of IFN-γ (A) and IL-12 (B) in serum were detected by ELISA. *P < .05 versus PBS; #P < .05 versus DC. DC indicates dendritic cell; LDR, low-dose radiation; IL-12, interleukin 12; IFN-γ, interferon γ; PBS, phosphate-buffered saline; ELISA, enzyme-linked immunosorbent assay.
In addition to serum IL-12, serum IFN-γ, which is a cytokine produced by CTLs generated by DC vaccines and plays a crucial role in protecting against tumors, was also detected. As shown in Figure 4B, an injection of a DC vaccine that was not exposed to LDR significantly increased the level of serum IFN-γ compared to that of the untreated control at both examined time points. Furthermore, an injection of a DC vaccine exposed to LDR at 0.2 Gy was found to elevate serum IFN-γ more efficiently than an injection of a DC vaccine without exposure, which was consistent with the changing pattern of serum IL-12.
Administration of a DC Vaccine Exposed to X-Ray Radiation at a Dose of 0.2 Gy Promoted the Tumor Infiltration of CTLs and Apoptosis of Tumor Cells
Theoretically, CTLs would reach the targeted tumor after DC vaccination, which is a key step for the DC vaccination to elicit immune responses against the tumor. We, therefore, examined the infiltration of CTLs in tumor tissues by immunohistochemistry with an anti-CD8 antibody. As shown in Figure 5A (upper panel) and Figure 5B, the administration of a DC vaccine exposed or unexposed to LDR significantly increased the infiltration of CTLs into tumor tissue compared to the untreated control. However, there were more CTLs in the tumor tissue of mice treated with the DC vaccination exposed to LDR than of mice treated with the DC vaccination that was not exposed to LDR.

Treatment with a DC vaccine exposed to LDR significantly enhanced the tumor infiltration of CTLs and tumor cell apoptosis. A, Tumor tissue was resected from mice 14 days after an initial treatment with PBS or a DC vaccine with or without exposure to LDR and was subsequently subjected to immunohistochemical staining using an anti-CD8 antibody and a TUNEL assay to detect the infiltration of CTLs (upper panel, ×400) and apoptotic tumor cells (lower panel, ×400) in tumor tissue, respectively. The CD8+ cells (B) and apoptotic cells (C) were quantified using Image-Pro Plus 6.0. * P < .05 versus PBS; # P < .05 versus DC. DC indicates dendritic cell; LDR, low-dose radiation; CTLs, cytotoxic T lymphocytes; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling; PBS, phosphate-buffered saline.
The secretion of IFN-γ and infiltration of CTLs into tumor tissue can both induce apoptosis of tumor cells though various mechanisms; the apoptotic cells were, therefore, detected by TUNEL staining. The results showed that there were more apoptotic tumor cells in the tumor tissue of mice injected with unexposed DC than in the tumor tissue of untreated control mice. More importantly, an injection of DC exposed to LDR resulted in more significant tumor cell apoptosis than the unexposed DC injection (lower panel of Figure 5A and C), which was similar to the changing patterns of serum IFN-γ and tumor infiltration of CTLs.
Discussion
The strong ability of DCs to induce an immune response against tumors makes DCs important for cancer vaccine development. Although several DC-based antitumor vaccines have entered clinical trials, their response rates are still unclear due to various issues, such as inefficient migration to LN and IL-12 production, which are associated, at least in part, with the current strategy for the ex vivo generation of DC vaccines. In the current study, we showed that exposure to X-ray radiation at a dose of 0.2 Gy during the preparation of DC vaccines enhanced the ability of DCs to migrate both in vitro and in vivo, stimulating T-cell proliferation and priming the antitumor response in vitro. After subcutaneous injections into mice-bearing tumors, the DC vaccine exposed to X-ray radiation at a dose of 0.2 Gy significantly prolonged the survival time of the mice, which was accompanied by increases in serum IL-12 and IFN-γ, the infiltration of CTLs, and cell apoptosis in tumor tissue compared to those of unexposed DC vaccine administration.
In our previous study using human PBMC-derived DCs that were exposed to X-ray radiation at dose of 0.05, 0.1, 0.2, or 0.5 Gy during the early (day 2) or late stage (day 6) of DC vaccine generation and lung adenocarcinoma cells, we found that treatment with 0.2 or 0.5 Gy of X-ray radiation on day 2 significantly inhibited the T-cell-stimulating activity, antitumor effect, and IL-12 secretion of DCs in vitro, while the exposure to LDR at a dose of 0.2 or 0.5 Gy on day 6 significantly promoted the T-cell-stimulating activity, antitumor effect, and IL-12 secretion of DCs in vitro. 21 However, exposure to LDR at a dose of 0.05 or 0.1 Gy at the early or later stage did not show any significant effect in regard to the T-cell stimulating activity, antitumor effect, and IL-12 secretion of DCs in vitro. 21 Considering that there was no significant difference in the promoting effects between exposures of 0.2 and 0.5 Gy, a single exposure at a dose of 0.2 Gy, which was expected to be safer than 0.5 Gy, on day 6 was selected for this study. In addition, a mouse lung adenocarcinoma cell line similar to that in our previous in vitro study 21 was also selected. To prime a robust immune response against tumors, a sufficient number of DCs in the vaccine must migrate from the injection site to the draining LN, where they activate immune cells. 25 However, a body of clinical studies indicated that less than 5% of injected DCs could migrate to the LN after subcutaneous injection, 12,13,26 which was considered to be one of factors that limited the clinical efficiency of DC vaccines. 27 Therefore, many efforts have been made to enhance the migratory capacity of DC vaccine through modifying the preparation procedure of DC vaccines. For instance, Rizzo et al 28 reported that pretreatment with low-molecular-weight hyaluronan during the preparation of DC vaccines enhanced the migratory capacity of DCs through increasing the expression of CCR7. Zhu et al 29 showed that the addition of CpG ODN and Poly (I: C) to the maturation cocktail could generate DC vaccines with higher migratory potential. In the present study, our results showed that exposure to LDR during the preparation of DC vaccines also promoted the migration of DCs. In an independent study by our group, we revealed that exposure to X-ray radiation at a dose of 0.2 Gy increased the expression of CCR7 by activating the ATM/NF-κB pathway in the immortalized mouse DC line JAWSII, which resulted in the enhancement of DC migration. 30
After migrating to LN, the ability of DCs to stimulate T cells is essential to initiate tumor immune responses. The current study showed that exposure to LDR enhanced the ability of DCs to stimulate T cells. Similarly, Shigematsu and his colleagues 31 showed that DCs could stimulate the proliferation of T cells more effectively after exposure to LDR. In contrast, Jahns et al 32 reported that exposure of DC precursors to LDR did not affect DCs in regard to their ability to stimulate T-cell proliferation. These inconsistent results may be caused by differences in the irradiation time point at which the DCs were in different developmental stages. In line with increased ability to stimulate the proliferation of T cells, the in vitro cytotoxicity of T cells induced by the LDR-exposed DC vaccine was significantly upregulated compared to that induced by the unexposed DC vaccine. More importantly, an in vivo study confirmed that the antitumor activity of the DC vaccine was significantly increased after exposure to LDR, as shown by the increased survival time of tumor-bearing mice; however, the DC vaccine without exposure to LDR did not significantly prolong the survival time of tumor-bearing mice compared to the untreated control.
The goal of DC vaccines is to activate CTLs, which will subsequently infiltrate the tumor and lead to tumor cell death through various mechanisms, such as the induction of cell apoptosis. 33 A previous clinical study showed that treatment with a DC vaccine resulted in robust CTL infiltration in tumor areas. 34 In the present study, treatment with a DC vaccine without exposure to LDR increased the number CTLs in the tumor compared to the that in the control, while the administration of a DC vaccine exposed to LDR further enhanced the infiltration of CTLs in the tumor.
It has been reported that apoptotic tumor cells can be observed at the site of CD8+ T-cell infiltration after treatment with a DC vaccine, which indicates that infiltrated CD8+ T cells kill tumor cells by inducing apoptosis. In the current study, a TUNEL assay showed that treatment with a DC vaccine exposed to LDR enhanced apoptosis of tumor cells compared to the administration of an unexposed DC vaccine, which was consistent with the results of CD8+ T-cell infiltration.
The IL-12 produced by DCs is crucial for Th1 differentiation and the activation of CTLs, 18,35 both of which play an important role in cancer rejection. Various clinical studies have demonstrated that the level of serum IL-12 was significantly increased after treatment with a DC vaccine, 36,37 which was consistent with our findings. However, the production of IL-12 by a DC vaccine generated by conventional methods was still insufficient, which was considered to be responsible for, at least in part, the unsatisfying clinical outcomes of using DC vaccines for cancer immunotherapy. 29,38 Dendritic cell vaccines with an increased ability to produce IL-12 have been proven to prime stronger CTLs in vitro and result in greater tumor growth inhibition in vivo. 38,39 However, the authors did not report whether the administration of such a DC vaccine could result in a further increase in serum IL-12. Our previous study showed that exposure to LDR at a dose of 0.2 Gy during the preparation of human DC vaccines significantly enhanced the production of IL-12 by DCs. 21 Further mechanism studies using JAWSII cells showed that, as with the increased migratory ability, the LDR-induced increase in IL-12 production was also mediated by the ATM/NF-κB pathway. 30 We also found that the activation of NF-κB by LDR continued until 90 minutes after LDR and that the increases in IL-12 and CCR7 could be detected up to 24 hours after LDR (unpublished data), which indicated that the stimulatory effect of LDR on DCs might be temporary. In the current study, we further confirmed that the level of IL-12 in the serum of tumor-bearing mice was further increased after the administration of DC vaccine exposed to LDR compared to that with the treatment with the unexposed DC vaccine. The increased level of systemic IL-12 would be beneficial to stimulate a burst of antitumor immune response, and this fact is supported by a clinical study in which a subcutaneous injection of IL-12 given to patients with metastatic melanoma increased the frequency of antitumor CTL precursors in peripheral blood, promoted the infiltration of CTLs in neoplastic tissue, and resulted in increased tumor necrosis and/or fibrosis. 40 Similar to IL-12, the level of IFN-γ in patient serum was also observed to be increased after treatment with a DC vaccine. 37 In line with this finding, the present study showed that treatment with an unexposed DC vaccine resulted in an increase in serum IFN-γ, while administration of a DC vaccine exposed to LDR led to a further increase in serum IFN-γ. It has been established that IFN-γ, which can be secreted by activated T cells, plays an important role in the immune defense against tumors through activating CTLs and NK cells or through directly affecting cancer cell survival. 41 –43 We therefore hypothesized that the increases in serum IL-12 and IFN-γ after treatment with a DC vaccine exposed to LDR were associated with enhanced CTL infiltration in the tumor. However, we did not further mechanistically investigate whether the in vivo effects of DCs exposed to LDR, including the increased survival time of tumor-bearing mice, elevated serum IL-12 and IFN-γ, and enhanced CTL infiltration in the tumor, could be abolished by pretreating DCs with inhibitors of ATM or NF-κB in the current study, which is a limitation of the study.
These experimental data support the conclusion that exposure to LDR at a dose of 0.2 Gy during the procedure of DC vaccine preparation may be a novel approach to generate DCs with superior antitumor activity. Further studies are needed to assess whether this novel approach of DC vaccine preparation is effective for other types of solid tumors, specifically for renal tumors in which DC-based immunotherapy has entered into a phase 2 study. 44
Footnotes
Authors’ Note
FL and QJ contributed to conceptualization. FL and QJ did funding acquisition. SW, HY, RH, XS, SC, NY, and WL did investigation. FL wrote the original draft. QJ wrote, reviewed, and edited the paper. Sinian Wang and Huijie Yu contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported in part by grants from the National Natural Science Foundation of China (81202151 and 31570854 to FL, 31770914 to QJ) and the Key Project of Military Logistics (AEP17J001 to QJ).