
Abstract
Background. Epidural stimulation of the spinal cord can reorganize and change the excitability of the neural circuitry to facilitate stepping in rats with a complete spinal cord injury. Parkinson’s disease results in abnormal supraspinal signals from the brain to the spinal cord that affect the functional capacity of the spinal networks. Objective. The objective was to determine whether epidural stimulation (electrical enabling motor control, eEmc) of the lumbosacral spinal cord can reorganize the spinal networks to facilitate hindlimb stepping of rats with parkinsonism. Methods. A unilateral 6-OHDA (6-hydroxydopamine) lesion of the nigrostriatal pathway was used to induce parkinsonism. Sham rats (N = 4) were injected in the same region with 0.1% of ascorbic acid. Stimulation electrodes were implanted epidurally at the L2 and S1 (N = 5) or L2 (N = 5) spinal levels. Results. The 6-OHDA rats showed severe parkinsonism in cylinder and adjusting step tests and were unable to initiate stepping when placed in a running wheel and dragged their toes on the affected side during treadmill stepping. During eEmc, the 6-OHDA rats initiated stepping in the running wheel and demonstrated improved stepping quality. Conclusion. Stepping was facilitated in rats with parkinsonism with spinal cord stimulation. The underlying assumption is that the normal functional capacity of spinal networks is affected by supraspinal pathology associated with Parkinson’s disease, which either generates insufficient or abnormal descending input to spinal networks and that eEmc can appropriately modulate spinal and supraspinal networks to improve the motor deficits.
Introduction
The triad of akinesia/bradykinesia, tremor, and rigidity reflects the motor deficits typical of Parkinson’s disease (PD), but postural instability, failure to initiate stepping and abnormal gait also add to patient’s disability. Presently
We propose that improved stepping can occur in rats with parkinsonism when the spinal networks are engaged appropriately. The underlying assumption is that the supraspinal pathology associated with PD either generates insufficient or abnormal descending input to spinal networks, leading to abnormal stepping. Can some advantage be gained from direct neuromodulation of the excitability of selected spinal networks? Evidence for this possibility is demonstrated by observations that even without any supraspinal input to the spinal cord, full weight-bearing locomotion can be achieved in adult rats with a complete spinal cord transection at a mid-thoracic level when eEmc and pharmacological facilitation are used to enable stepping.6,7 In contrast to Fuentes et al, 3 we found that tonic stimulation at 40 Hz, but not at higher frequencies such as 300 Hz, facilitated stepping quality in these spinal rats.6,7 A relevant underlying principle to consider is that the extensive redundancy in the sources of interactive control of the motor system provides a potential strategy to minimize the impact of the loss of a single source of control. In addition, in our initial studies of human subjects with a complete spinal cord injury for more than a year we observed recovery of standing and stepping and several autonomic functions in response to eEmc,8,9 illustrating the integrative nature of multiple physiological systems as adaptive responses emerge. The objective of the present study was to determine the potential of eEmc of the lumbosacral spinal cord to enable rats with 6-OHDA-induced motor deficits to initiate and improve quality of stepping.
Methods
Fourteen adult male Sprague-Dawley rats (200-250 g body weight) were assigned randomly to a 6-OHDA (N = 10) or sham (N = 4) group. The rats were housed individually with food and water provided ad libitum in a room with a 12:12-hour light-dark cycle and maintained at ~25°C. All surgical procedures were conducted aseptically with the rats deeply anesthetized (isoflurane gas, 1.5%-2%, administered via facemask). All experimental procedures complied with the guidelines of the National Institute of Health Guide for the Care and Use of Laboratory Animals and were conducted in accordance with protocol approved by the Animal Care Committee at the University California, Los Angeles, CA.
6-OHDA Injections
Thirty minutes prior to surgery, the noradrenergic reuptake inhibitor desipramine (25 mg/kg, Sigma Chemicals, St Louis, MO, USA) was injected intraperitonealy. A small opening was drilled in the skull at 5.6 mm posterior to bregma, 2 mm lateral to the midline, and with the skull leveled between bregma and lambda. Four microliters of 0.2% 6-OHDA (Sigma) in 0.1% of ascorbic acid (6-OHDA group) or 4 µL of 0.1% ascorbic acid (sham group) was injected into the right substantia nigra 7.4 mm ventral to the brain surface, using a 10 µL Hamilton syringe pump at 0.5 µL/min (KDS Model 310 Plus Series, kdScientific, Holliston, MA, USA).
Head Connector, Spinal Epidural Electrode, and EMG Electrode Implantation
Head connector, epidural electrode, and electromyography (EMG) implantation procedures have been described previously.6,10 The epidural electrode was implanted at the L2 (N = 5) or the L2 and S1 spinal cord levels. The rats with both L2 and S1 epidural electrodes also were implanted with EMG recording electrodes in the soleus and tibialis anterior (TA) muscles bilaterally as described previously. 10
Data Collection
We characterized the response of several motor tasks over a period of up to 4 weeks to determine the potential efficacy of spinal stimulation using several locomotor assays. Given that none of these specific motor tasks had been tested in the PD model, testing the efficacy of spinal stimulation on these tasks at the various time points provided us with a more complete view on the overall locomotor abilities.
Epidural Stimulation Parameters
The stimulation threshold was determined as the minimum voltage producing a palpable contraction in any hindlimb muscle. A subthreshold voltage (~90% of threshold) was used during stimulation. 11 The stimulation protocols used were (1) tonic stimulation (repeated pulses) at 40 Hz or 333 Hz with a pulse width of 0.2 ms and (2) train stimulation, that is, 1 train/s for 250-ms train duration at 40 Hz with a pulse width of 0.2 ms. The direction of the stimulation was from L2 to S1 (N = 5) or L2 to ground (N = 5).
Running Wheel Test
At 2 and 4 weeks post-lesion, all rats were placed in a cage with a running wheel the night before the test for acclimatization. The running wheel is passive, that is, it has no motor drive and only rotates when the animal is attempting to run. For each test, the rat was placed in the running wheel without eEmc for about 10 to 30 seconds and then with eEmc. Each rat received 10 to 15 bouts of stimulation with 5 to 30 seconds (depending on the duration of the previous bout) of no stimulation between each bout of stimulation. The rats were videotaped from underneath the running wheel. The incidences of initiation of stepping with the stimulation “off” or “on” were determined from the video (a light showing time of stimulation) and reported as the percent of successful initiations of stepping (number of initiations of stepping/number of bouts × 100). The duration of stepping during each bout with the stimulation “off” or “on” was expressed as a percent (stepping time [seconds]/bout [seconds] × 100). The experimenter was blinded to the group identification but not the light indicating when the stimulation was on or off.
Bipedal Treadmill Stepping Test
Bipedal stepping ability was determined at 1 week (N = 5, with stimulation between L2 and S1) or at 4 weeks (N = 5, with stimulation between L2 and ground) post-lesion with and without eEmc. During the test, the rats were supported in a body harness over the treadmill in a semi-erect position to facilitate bipedal stepping (13.5 cm/s) and kinematics was recorded as described previously. 12 Some of the animals showed abnormal foot placements such as dragging the toe at the beginning of the swing phase and/or laterally rotating the foot during stance. EMG was recorded in 5 rats 4 weeks post-lesion at a sampling frequency of 2 kHz using a custom-made LabView program. EMG burst amplitudes, durations, and integrals for the 10 steps were determined as described previously. 10 The dragging and asymmetric foot placement are common and clinically significant deficits occurring after a neuromuscular dysfunction.13,14 The experimenter performing the data analysis was blinded to the group assignment of each rat.
Immunohistochemical Analyses
Four weeks after brain injection, the animals were anesthetized deeply with sodium pentobarbital (100 mg/kg, intraperitoneally) and intracardially perfused with 0.1 M phosphate buffered saline (PBS, pH 7.4) at room temperature, followed by 4% paraformaldehyde in 0.1 M PBS (pH 7.4). The brains were removed quickly, postfixed in the same fixative overnight at 4°C, cryoprotected in 30% sucrose in 0.1 M PBS, frozen on powdered dry ice, and stored at −80°C. Coronal brain sections (40 µm) were cut on a Leica CM 1800 cryostat (Deerfield, IL, USA) and stored in cryoprotectant at −20°C. Striatal sections at the level of bregma 2.20 mm, 0.48 mm, and −0.92 mm were stained for tyrosine hydroxylase (TH). The immunostaining for TH was performed as described previously. 15 Images of the striatum were taken using a Zeiss Axioskop microscope (Gottingen, Germany) with a Spot digital camera (Diagnostic Instruments, Inc, Sterling Heights, MI, USA). Optical density (OD) measurements of TH immunoreactivity in the striatum were performed using Image J (Image J 1.45s, National Institutes of Health). The adjacent section from the same animal were stained without the primary anti-TH antibody was served as control section. After calibration using a step tablet, the OD in each striatum was determined. OD measurements from control sections were deducted to eliminate any background signal. If the OD.value was lower than 0, 0 was assigned as the final value. TH data are presented as a percentage of TH OD for the ipsilateral side compared to the contralateral side with a minimum of 0. Due to technical difficulties, the brain tissue from 2 animals were unusable. Thus, TH analysis were done only on 8 6-OHDA rats.
Statistical Analyses
SigmaPlot 12 (Systat Software Inc) was used for all statistical analyses. The Mann-Whitney test was used to determine group differences in TH OD. Two-way repeated-measures analysis of variance followed by the Bonferroni test was used to compare group differences in the success rates of initiation of stepping, stepping time/bout, percent of abnormal foot placement, and EMG kinematics. Significant differences were determined at P < .05.
Results
Confirmation of Unilateral Lesion
Animals injected with 6-OHDA showed significant loss of TH immunoreactivity on the lesioned side compared with sham-treated animals indicating a loss of dopamine in this area (P = .012, Mann-Whitney test, Figure 1). The TH immunoreactivity ranged from 27% to 100% in the 6-OHDA rats. In both of cylinder test and adjusting step test the 6-OHDA rats showed significant asymmetry on the limbs on the injured side (Supplementary Figures 2 and 3, available online).
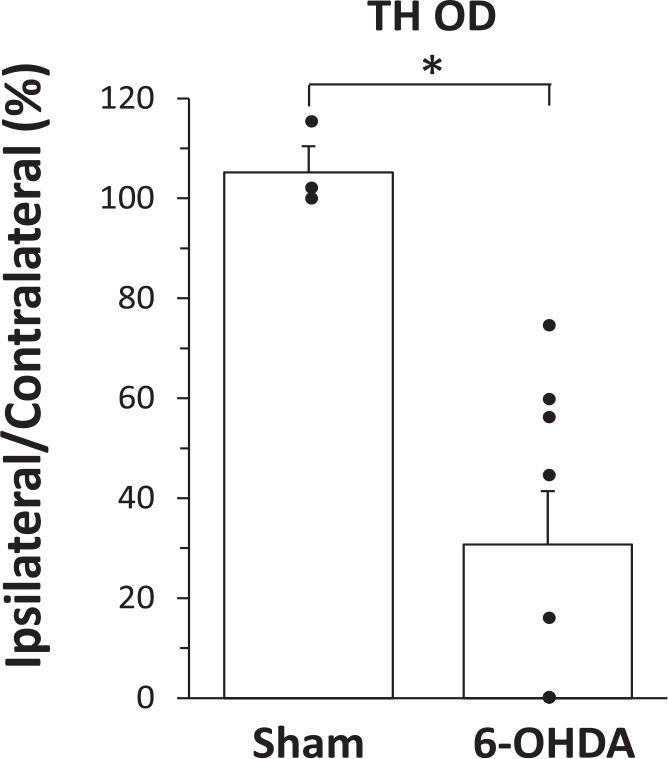
Optical density measurement of tyrosine hydroxylase (TH OD) in the striatum of sham and 6-OHDA (6-hydroxydopamine)-treated rats. Data are expressed as percentage of the ipsilateral/contralateral side. *Significantly different from the sham rats, P < .05. N = 3 (sham) N = 8 (6-OHDA, note, OD values are 0 in 3 rats).
eEmc Enables the Initiation of Stepping in PD Rats
We observed clear differences in the ability of sham versus 6-OHDA and of 6-OHDA rats with versus without eEmc when placed in a running wheel. The data of stepping initiation were similar for different stimulation patterns (tonic vs train) and sites (Supplementary Figure 1), and therefore were combined for analyses. Sham rats initiated stepping 40% to 50% of times when placed in the running wheel and this was similar with and without eEmc at 2 (Figure 2A) and 4 (Figure 3A) weeks post-lesion. This indicated that the eEmc did not make the sham rats step more on running wheels. In contrast, the 6-OHDA rats did not initiate any stepping at 2 weeks post-lesion (Figure 2A) and in only 2% of the cases at 4 weeks without eEmc (Figure 3A). With eEmc, however, the 6-OHDA rats significantly improved the percent successful initiation of stepping at 2 weeks (51%) and at 4 weeks (41%) to levels of the sham rats (Supplementary Video 1). Since the rats received sub-motor threshold stimulation at the lumbosacral spinal levels, neither did it induce stepping in either the forelimbs or hindlimbs nor cause a startle response. The sub-motor threshold neuromodulation of the spinal networks increases the level of network excitability close to a threshold to increase the susceptibility (probability) of generating functional movements. Thus, we refer to the higher excitable physiological state of the relevant motor pools as an enabling function.
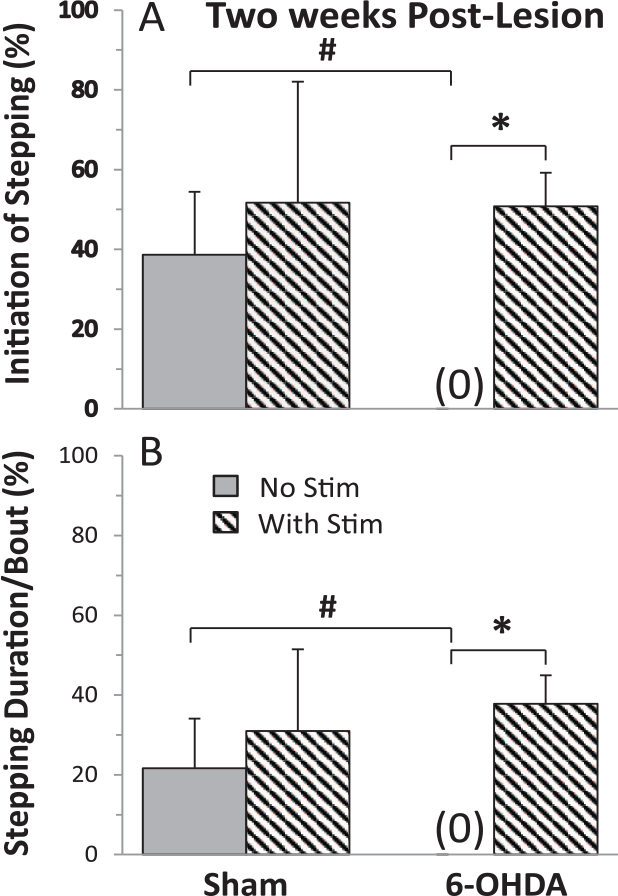
The percentage of successful initiation of stepping (A) and the duration (B) that the rats stepped during each bout of stimulation (Stim) when the rats were placed in a running wheel with and without stimulation at 2 weeks post-lesion. Values are mean ± SEM (10-15 bouts of stimulation/rat) for 4 and 10 rats for the sham and 6-OHDA (6-hydroxydopamine) groups, respectively. *Significantly different from No Stim within a group and #significantly different from sham rats with No Stim, at P < .05.
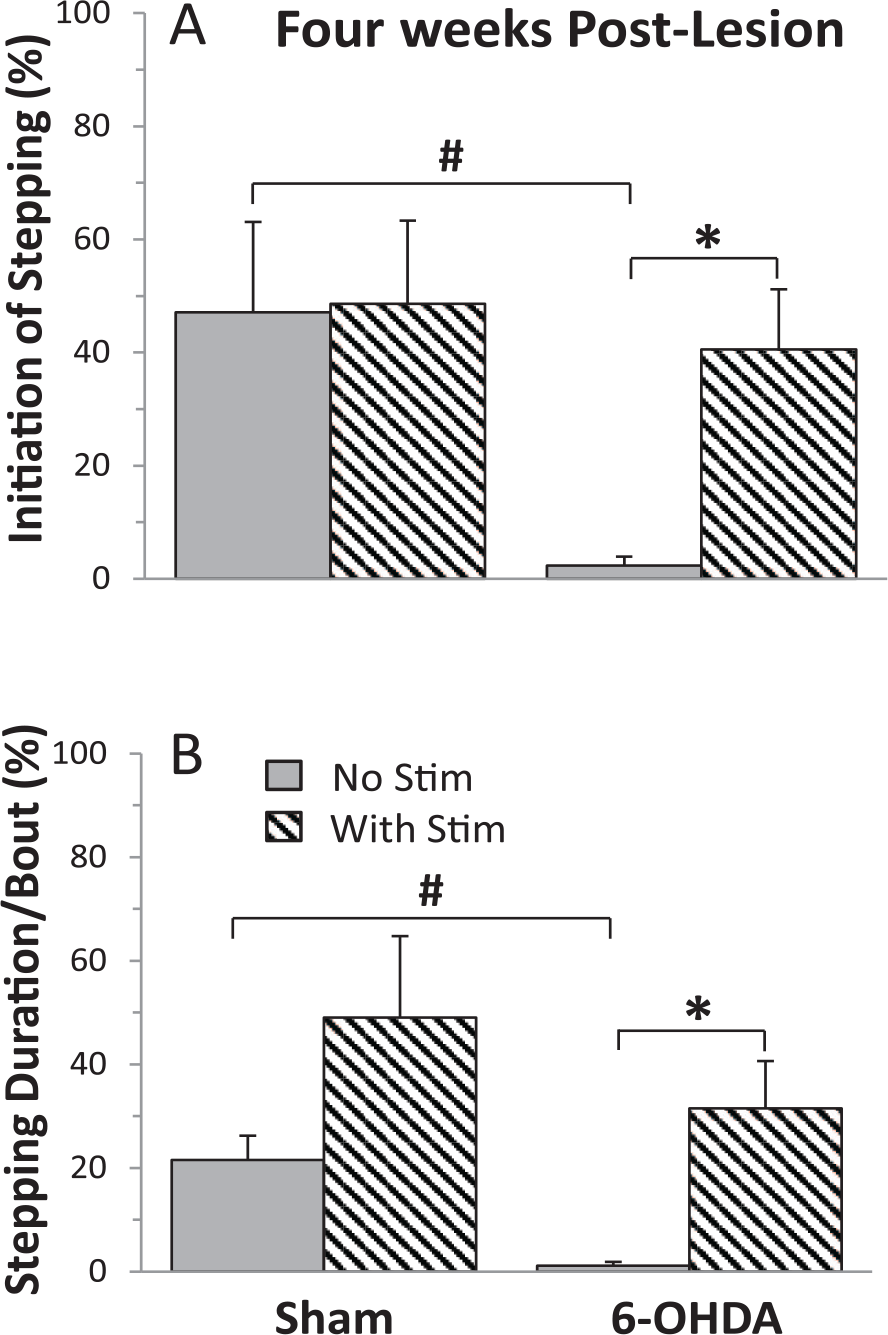
The percentage of successful initiation of stepping (A) and the duration (B) that the rats stepped during each bout of stimulation (Stim) when the rats were placed in a running wheel with and without stimulation at 4 weeks post-lesion. Values are mean ± SEM (10-15 bouts of stimulation/rat) for 4 and 10 rats for the sham and 6-OHDA (6-hydroxydopamine) groups, respectively. *Significantly different from No Stim within a group and #significantly different from sham rats with No Stim, at P < .05.
With no eEmc, the percent of the time running in the wheel for the sham rats was ~21% at 2 and 4 weeks post-lesion (Figures 2B and 3B). With eEmc these values increased to 31% and 49% but were not significantly different from no stimulation. The 6-OHDA rats initiated stepping only ~1% of the time without eEmc (P = .0012, compared with sham rats) but this time was significantly increased to 38% and 32% with eEmc at 2 and 4 weeks (Figures 2B and 3B). The incidences of initiation of stepping of the 6-OHDA treated rats with eEmc was not related to the severity of the brain lesion at 2 weeks (r2 = 0.014, P = .78) or 4 weeks (r2 = 0.417, P = .08) post-lesion.
eEmc Improves the Quality of Stepping in PD Rats
One of the purposes of this study is to determine whether the eEmc improves the motor deficits. Sham rats were able to step normally with and without eEmc based on EMG and kinematics analyses. Abnormalities in the kinematics were observed in 6-OHDA rats (Figure 4). Dragging the paw and abnormal foot placement of contralateral hindlimbs are also observed in multiple neurological conditions.7,14 With no eEmc, the 6-OHDA rats dragged their paw in the beginning of the swing phase and/or rotated their foot laterally during the stance phase in the hindlimb contralateral to the lesion side in 54% and 82% of the steps at 1 and 4 weeks post-lesion, respectively. No such abnormalities were observed in the ipsilateral hindlimb (Supplementary Video 2). With tonic stimulation at 40 or 333 Hz (L2 to S1), there were no abnormalities in the contralateral hindlimb at 1 week post-lesion. At the higher frequency of tonic stimulation 2.5% of the steps were abnormal in the ipsilateral hindlimb at 1 week post-lesion. At 4 weeks post-lesion, abnormalities in the kinematics in the contralateral limb occurred in 56% of the steps with tonic stimulation (40 Hz, L2 to ground). With trains of stimulation at the same frequency (40 Hz with 0.2-ms burst duration and cycle frequency of 1 train/s, L2 to ground) the number of abnormalities in the contralateral limb was reduced from 82% to 7% (Figure 4B). Combined, these data indicate that eEmc improved the quality of stepping of 6-OHDA rats including reduced dragging and abnormal foot placement of the affected hindlimb.
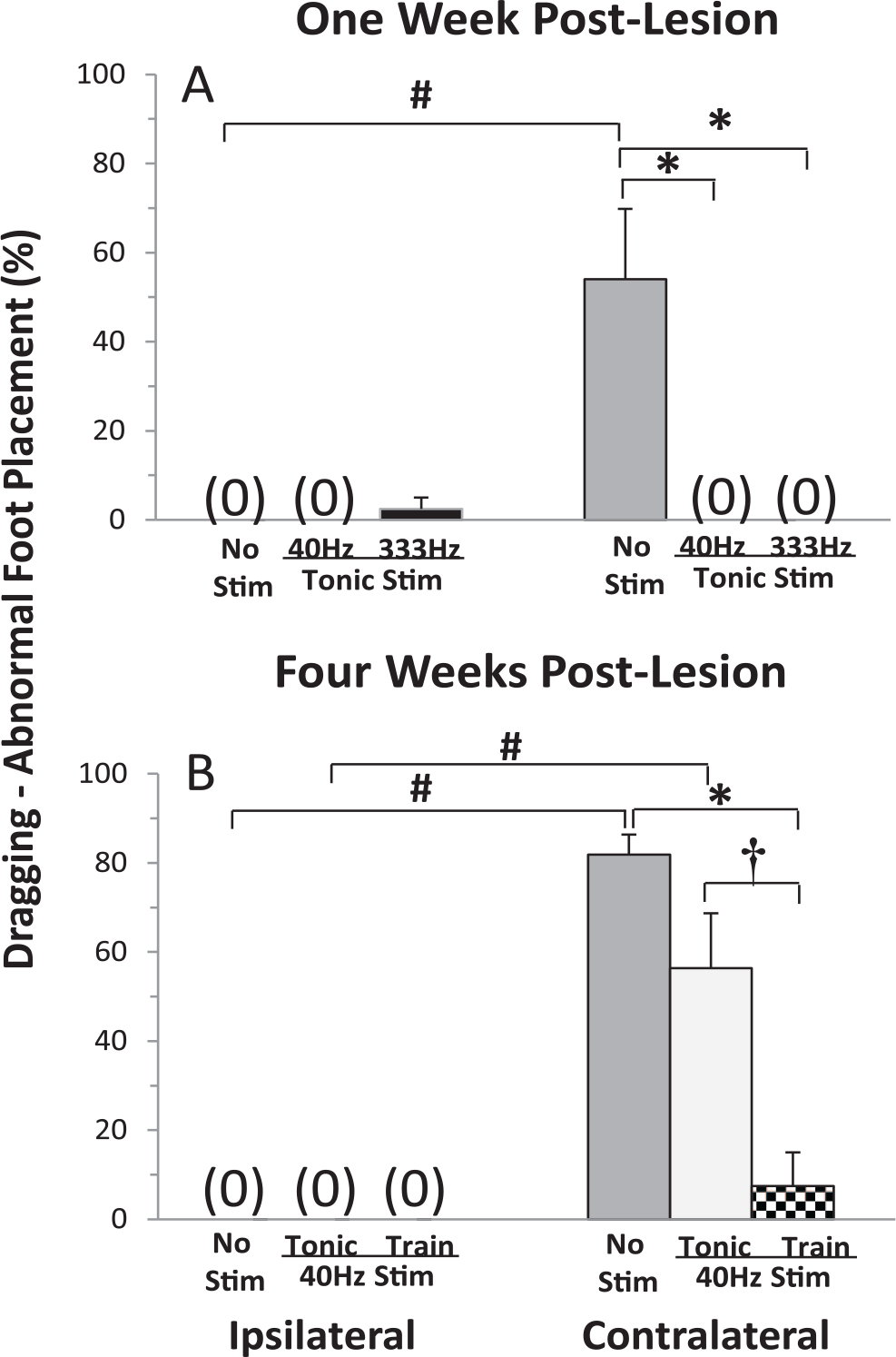
The percentage of steps showing dragging and/or abnormal foot placement of the ipsilateral and contralateral hindlimbs in the 6-OHDA (6-hydroxydopamine) rats during bipedal stepping on a treadmill at 13.5 cm/s with and without stimulation. Values are mean ± SEM for 5 rats at each time point (10 steps/rat). *,†Significantly different from No Stim and Tonic Stim within same side, respectively and #significantly different from the same condition on the ipsilateral side at P < .05. Note: The 1 week post-lesion group (N = 5) was stimulated between L2 and S1 spinal cord level; the 4 weeks post-lesion group (N = 5) was stimulated between L2 and ground.
The mean trajectories of the paw and measures of interlimb coordination for 10 consecutive steps under each condition for a representative sham rat and a 6-OHDA rat at 4 weeks post-lesion are shown in Figure 5. The paw trajectories in the X-Y plane and interlimb coordination in the horizontal (x) and vertical (y) planes in the sham rats were not different with and without eEmc. In contrast, there were contralateral-ipsilateral differences in the 6-OHDA rats. The trajectories and interlimb coordination patterns most closely resembling those in sham rats at 4 weeks post-lesion with train stimulation (Figure 5C). Detailed analysis of the foot trajectory demonstrates the impact of eEmc during the transition from the stance phase to the swing phase, resulting in reduced dragging (Figure 5E). In addition, these kinematics observations are consistent with the TA EMG firing pattern in late stance and early swing during treadmill stepping compared with no stimulation. Note the minimal change in soleus EMG between no stimulation and trains of stimuli as compared with the TA, correlating to the marked improvement primarily during the transition between the stance and swing phases (Figure 5F).
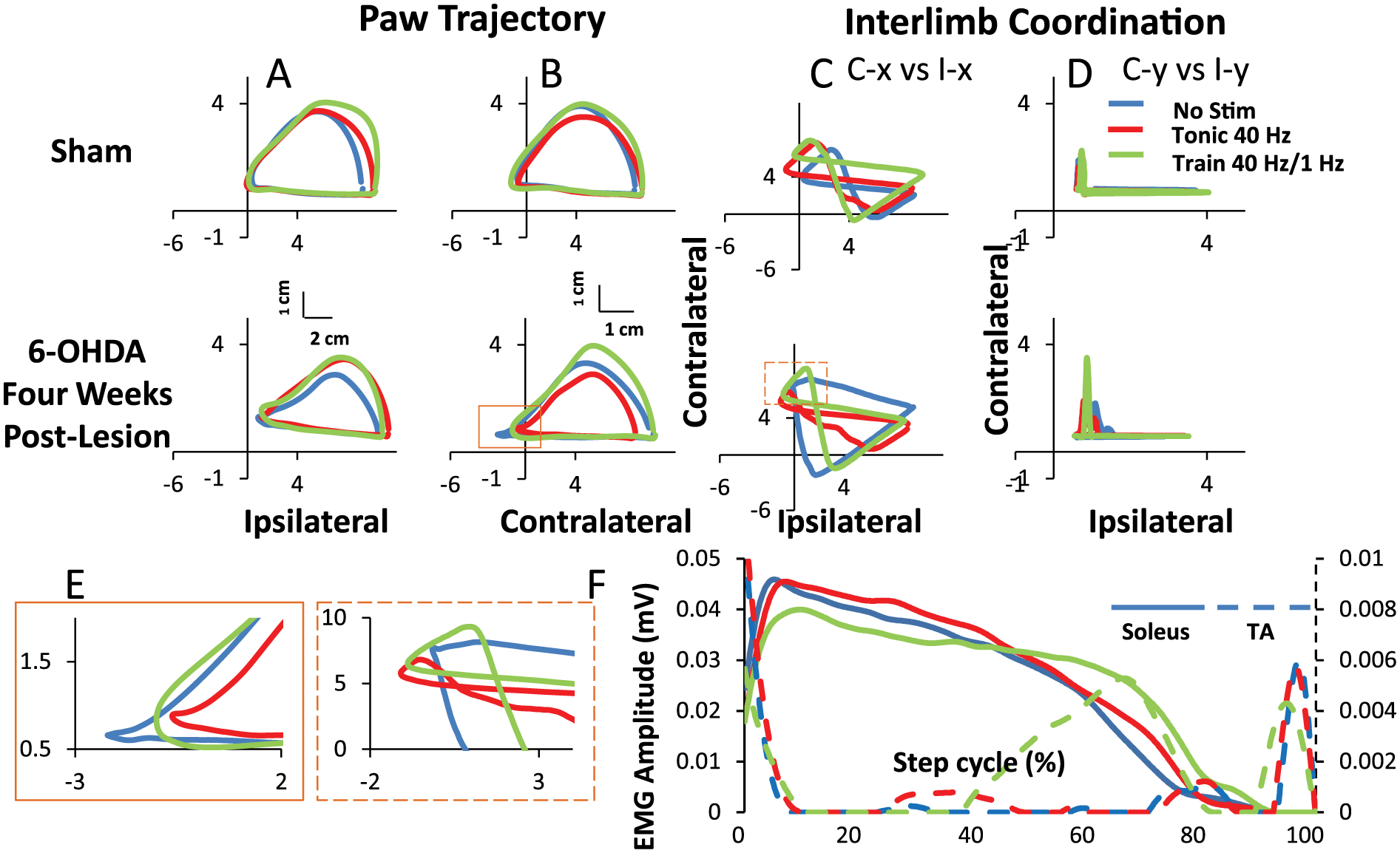
Trajectory of the paw for the ipisilateral (A) and contralateral (B) hindlimbs and interlimb coordination (C and D) during bipedal stepping on a treadmill at 13.5 cm/s from a representative sham and 6-OHDA (6-hydroxydopamine) rat at 4 weeks post-lesion with and without stimulation (No Stim). Line plots represent 10 steps for each rat for each condition. C-x, contralateral x axis; I-x, ipsilateral x axis; C-y, contralateral y axis; I-y, ipsilateral y axis. E represents a zoomed in view of the boxes. F, Average EMG (n = 5) over a normalized step cycle starting at heel strike comparing with and without stimulation.
The soleus and TA mean EMG integrals and burst durations during bipedal treadmill stepping were not different for the contralateral-ipsilateral hindlimbs of the sham rats (Figure 6). This also was true for the 6-OHDA rats, except that there was a tendency (P = .057) for the EMG burst duration to be higher in the TA of the contralateral with stimulation compared to no stimulation. The mean integral EMG’s were not significantly different when stepping was facilitated with either a tonic or train pattern of stimulation in the TA of both the ipsilateral and contralateral limbs. The average EMG patterns (Figure 5F) are consistent with the overall EMG analysis of burst durations and integrals (Figure 6).
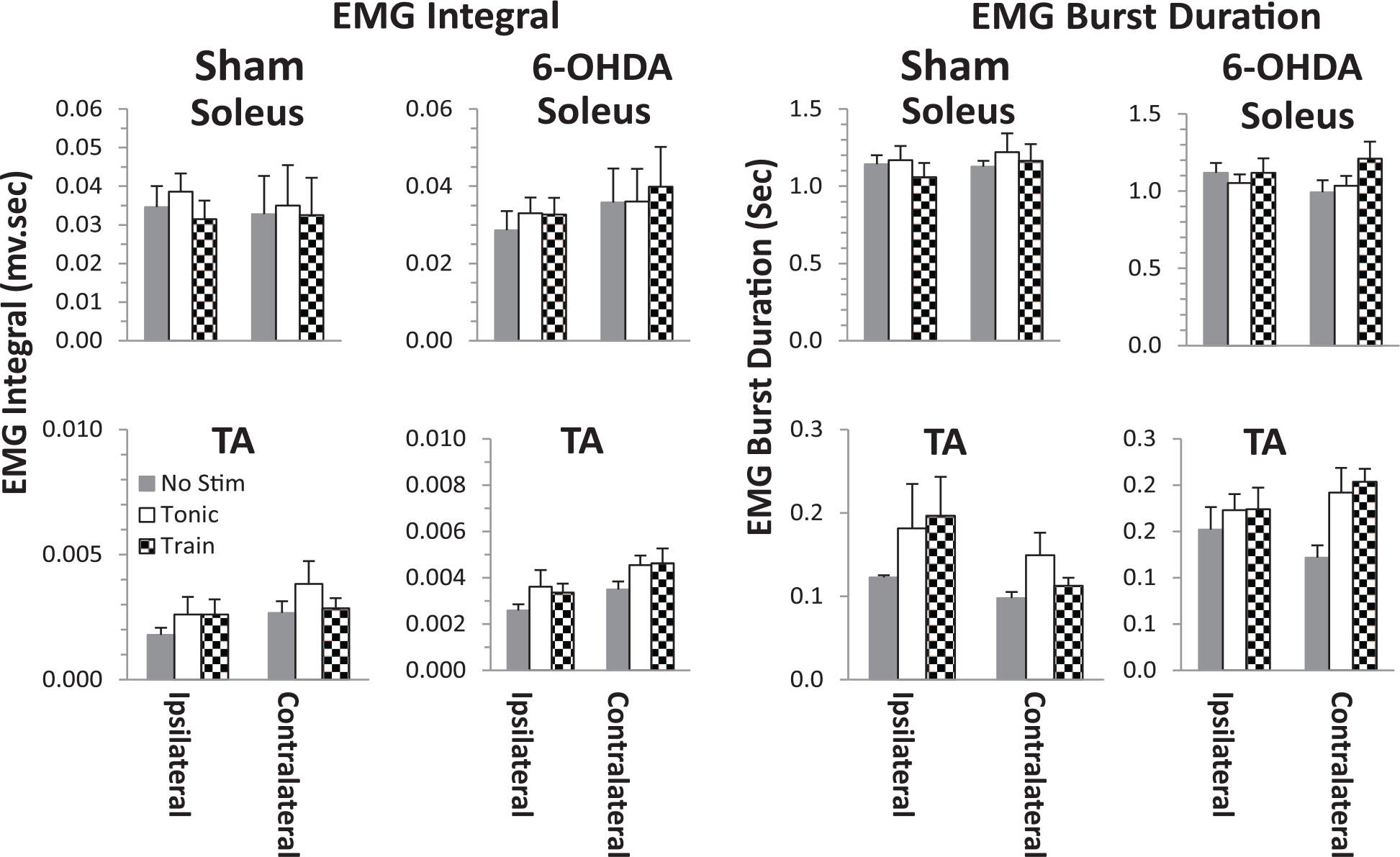
Soleus and tibialis anterior (TA) EMG integrals and mean burst duration for the Sham and 6-OHDA (6-hydroxydopamine) rats during bipedal stepping at 13.5 cm/s at 4 weeks post-lesion with (tonic and trains of 40 Hz) and without (No Stim) stimulation. Values are mean ± SEM for 10 steps in 4 and 5 rats in the Sham and 6-OHDA groups, respectively.
Discussion
Novelty of the Observations and Potential Mechanisms
Several potential motor assays were used to assess the effects of the 6-OHDA model of PD over a period of 1 to 4 weeks postinjury. This time period for studying the motor performance of 6-OHDA rats was similar to that which has been studied previously.16-18 The behavioral, kinematics, and EMG data demonstrated that spinal locomotor networks in unilaterally 6-OHDA-lesioned rats had difficulty in initiating stepping in a running wheel, although a rhythmic gait pattern could be generated. The gait pattern, however, was abnormal. With eEmc of the lumbosacral spinal cord, the 6-OHDA-lesioned rats initiated stepping in the running wheel with a frequency similar to that of uninjured rats and showed improvement in the quality of bipedal stepping on a treadmill. Previous studies exploring the possibility of stimulating the spinal cord to counter the progression of sensory and motor deficits associated with PD suggest that there is considerable potential in this interventional approach to improve motor function in PD.3,4,16,19-25 The mechanism for the improvement in those studies is generally to be via activation of the spinal dorsal somatosensory pathways, which project mainly to the thalamus.26,27 Given that spinal stimulation also facilitates hindlimb posture and locomotion in animals after a complete mid-thoracic spinal transection, we examined whether similar neromodulatory strategies could improve motor function after 6-OHDA treatment. We reasoned that (1) neuromodulation of the physiological state of spinal networks could compensate for the abnormal descending signals from the pathological regions of the brain and/or (2) some combination of both spinal and supraspinal mechanisms could have additive or even synergistic effects.
The present data demonstrate the possibility that the mechanism(s) underlying the improved motor function using spinal cord stimulation could be attributed, at least in part, to the neuromodulation of the spinal networks to a physiological state that enables the spinal interneurons to process extensive, but highly organized, proprioceptive input specific to a given motor task. We have reported that the constantly changing proprioceptive ensembles that are processed in real time can control very complicated movements with excellent coordination. 28 Although the actual mechanisms among the networks involved in the spinal control of movement are not well defined, some basic principles of control at the systems level are emerging. Based on numerous studies the efficacy of neuromodulation of spinal networks to regain stepping in absence of any input from the brain is substantial.6,7,13,29 The present results are consistent with the possibility that the improved motor functions observed could be attributed, at least in part, to direct effects on spinal networks. If the functional integrity of both spinal and supraspinal networks can be improved with neuromodulation, perhaps a synergistic optimization may lead to greater functional outcomes.
One of the unique motor assessments that we used in this study was to determine whether eEmc could alleviate the difficulty in initiating stepping in 6-OHDA rats. Kravitz et al 30 demonstrated that a “freezing” behavior could be controlled optogenetically in vivo in mice by activating the direct and indirect pathways of medium spiny projection neurons. Activation of indirect pathways induced a parkinsonian state, whereas stimulating the direct pathways reduced the freezing effect and increased locomotion. The authors concluded that the basal ganglia circuitry has a critical role in the bidirectional regulation of movement. Subsequent results suggest that a subset of basal ganglia neurons serve as an inhibitory gate over motor output that can be opened or closed by striatal direct and indirect pathways, respectively, resulting in movement initiation or motor suppression. 31 All these data are consistent with the possibility of a significant interaction between ascending signals derived from locomotor-related proprioception in activating circuits in the sensorimotor regions of the basal ganglia and the sensorimotor cortex. These findings would be consistent with a spinal-supraspinal interaction that would occur as seen with exercise interventions having marked immediate and chronic effects in improving motor control. 32 During lower limb exercises there are major increases in proprioception that amplify both spinal and supraspinal networks. For example, proprioceptive and cutaneous input can control hindlimb stepping without any input from the brain after complete spinal transection. 33 Uncertainty of the mechanisms of action during spinal stimulation, remains, however, when ascending connections remain intact as in the 6-OHDA model. At the same time, it has been documented numerous times the same stimulation parameters are shown to be effective without any input from the brain. Together, these findings are consistent with the possibility that spinal stimulation could alleviate parkinsonism via direct effects on spinal networks and/ or supraspinal networks. Taking the present results combined with previous studies reflecting activity-dependent plasticity, future studies are needed. The effects of chronic spinal neuromodulation on both supraspinal and spinal networks and the specific behavioral outcomes that could be attributable to the modulation of one or both networks in response to acute and chronically applied interventions while being trained for specific tasks need to be studied.
Behavioral Responses Are Tightly Linked to the Specific Combination of the Stimulation Parameters and the Physiological State of the Networks Being Neuromodulated
Based on a range of experimental models, the site and frequency of stimulation of the previous studies, and the parameters used in the present study, it is rather clear that a concerted effort is needed to develop spinal cord stimulation parameters and methodologies to optimize the potential of this intervention. Epidural stimulation (130 and 300 Hz) at a high cervical region of the spinal cord had no effects for relieving akinesia or restoring locomotor ability in 2 PD patients. 24 Stimulation at T9-T10 (130 Hz), which is closer to the upper lumbar segments that are known to be important in controlling locomotion, alleviated motor symptoms including tremor, rigidity, and bradykinesia in 1 PD patient, although gait was not modified. 20 An independent study reported improvement in gait in 4 PD patients with stimulation at T2-T4 (300 Hz). 34 Whereas, a recent study in 5 patients with epidural stimulation at T8-T10 showed that the best stimulation frequencies were 30 to 130 Hz in gait improvements. 25
We have explored stimulation patterns using low-frequency monopolar and bipolar stimulation or local versus global stimulation that have resulted in different kinematics patterns in animal models35,36 and in human subjects 9 indicating that along with location and frequency, the site of stimulation needs to be tested to optimize the functional response. In addition, the intensity of stimulation also must be considered. The stimulation parameters that are more effective in enabling initiation of stepping may not be the more effective one for optimizing the kinematics of posture and locomotion. In the present study, a sub-motor threshold intensity of stimulation was used, that is, an intensity that does not produce a direct response of the motoneurons projecting to the associated muscles. In many cases once stepping was initiated, it continued until the stimulation was stopped, consistent with an enabling of the initiation of rhythmic patterns of activation rather than a startle response. Also, the absolute voltage or current that we used induced little to no detectable startle-like responses. We have studied the response of many able bodied subjects to spinal transcutaneous stimulation of the lumbosacral segments which routinely induces involuntary rhythmic stepping motions without complaints of discomfort.37,38 The feasibility of stimulating the spinal networks with parameters that are similar to those used for spinal injury and can facilitate motor function in Parkinson’s subjects has been demonstrated. For example, Samotus et al 25 observed in 5 Parkinson’s subjects with an average age of 71 years and 14 years of PD symptoms improving in multiple measures of locomotion. After 6 months of daily stimulation for 12 to 15 hours per day with mid-thoracic epidural stimulation (60 Hz at 3% to 5% above sensory threshold) the subjects freezing episodes were reduced from 16 to 0 per day in absence of stimulation, while remaining on levodopa medication.
A novel feature of the present study is the development and use of an experimental assessment tool to specifically address the difficulty in initiating a movement such as stepping phenomenon, a behavior similar to that observed in many individuals with PD. Uninjured rats initiated stepping 40% to 50% of times placed in the running wheel and this was similar with and without eEmc. There was a near absence of the initiation of stepping in the 6-OHDA-treated rats without stimulation. The incidence of initiation of stepping in the presence of eEmc, however, was similar to sham rats. The duration of stepping was similar between sham and experimental animals with eEmc. Thus, the stimulation did not cause an abnormal amount of stepping in the sham animals and also the amount of stepping in the experimental animals was similar to the sham. The longer duration of the stepping once it was initiated in the present study is consistent with this response not simply being a supraspinal startle response.
While the present observations are not necessarily incongruent with the spinal eEmc effects being mediated in part via supraspinal methods, neither is there compelling evidence to exclude the likelihood that the regaining of function as observed in the present study be attributed directly to the facilitation of spinal networks. While these data generated from modeling of the structures activated with spinal epidural stimulation has been interpreted primarily based on the large size of ascending axons
39
of the dorsal columns, the effects have been attributed to supraspinal rather than spinal sites. The limitations of the modeling, however, is based largely on anatomical features of axons. It does not include anatomical or physiological properties of dendrites and their filamentous features. Neither does it include the continuous facilitating actions on dendritic filamentous structures and synapses on these dendritic structures, which for most neurons makes up more than 90% of a neurons surface. These facilitating effects have been reported to represent a dominating effect on the physiology of neurons in vivo.40,41 The significance of the limitations inherit in the modeling of neuronal actions as noted above essentially assumes that the neuronal control only occurs once an axon is generates an action potential. Based on extensive observations of epidural and transcutaneous spinal stimulation as well as pharmacologically, it is clear that behavioral features are highly dependent on facilitating mechanisms. Therefore, the data derived from modeling alone may be of marginal relevance to the issue of our stimulating procedures acting only via ascending axons versus spinal networks or possibly via both mechanisms. In either case, it seems that the spinal stimulation intervention should eventually be combined with pharmacological approaches that could enable the lowering of
What Might Be the Advantages of Spinal Cord Stimulation as Opposed to Deep Brain Stimulation and Pharmacological Treatment of PD?
There are several potential advantages of spinal cord stimulation relative to the more conventional interventions. The first level intervention usually attempted with the onset of PD is pharmacological, that is, prescribing
As PD advances, falls and abnormal gait become a major source of disability for patients. These symptoms are not well managed with pharmacotherapy or DBS targets. 42 Spinal cord stimulation on the other hand targets neural networks primarily responsible for control of locomotion and has the capability to function with some level of independence of supraspinal input. Thus, by targeting these intrinsic spinal networks, one could have better temporal control over the locomotor patterns as compared with more distant sources of control using DBS. Our newly developed noninvasive transcutaneous method of spinal cord stimulation 43 also may have positive results and be more readily accepted by severely impaired patients and could be used in conjunction with a pharmacological intervention.44,45 Compared with DBS results, transcutaneous spinal cord stimulation is safer, less invasive, less costly, and may be as effective and more adaptable to a given patient, especially the geriatric population.
Activity Dependence as a Factor in Enabling Functional Improvements Among Spinal Networks in PD
Another feature of spinal cord stimulation having an effect on spinal networks that could prove to be of further benefit is the activity dependence of spinal network plasticity with spinal learning being part of this plasticity. Because learning is a component of this activity-dependent plasticity, it seems probable that the impaired motor output emanating from the pathology of the brain in individuals with PD imposes dysfunctional motor signals on the spinal networks which become learned. The positive side of this possible negative phenomenon, however, is that, perhaps, the learned dysfunction may be overcome by neuromodulation of the spinal networks in a manner that negates the abnormal signals descending from the brain. The activity-dependent nature of individuals even with severe PD symptoms has been demonstrated by almost immediate improvement in motor function with repetitive exercise.46-48 Thus, a logical next step is to combine motor training with spinal cord stimulation over a period of weeks or even months as has been done after a spinal cord injury.5,7,9,25
Limitations of the Study
While the present results suggest some obvious directions to pursue in developing more effective interventional strategies for PD, one cannot assume that the mechanisms for PD-related deficits are the same as those of 6-OHDA-related deficits. The data pertaining to the alleviation of the phenomenon of having difficulty in initiating stepping in the 6-OHDA parkinsonian model, however, are definitive. Further, the work of Samotus et al 25 in 5 Parkinson’s subjects showing PD symptoms improving in multiple measures of locomotion after 6 months of daily epidural stimulation for 12 to 15 hours per day demonstrates the feasibility and probability that neuromodulation can be an effective intervention. None of these studies to date, however, provide clear mechanistic effects regarding the relative impact that can be attributed to spinal cord stimulation effects on spinal networks versus via ascending projections to the brain. Further studies will be needed to determine the spinal and supraspinal mechanisms that might contribute to improvement of motor function with spinal stimulation.
Implications
The present results suggest that stepping can be initiated, and the quality of stepping can be improved with eEmc in adult rats with parkinsonism due to unilateral nigrostriatal degeneration.
Supplemental Material
SuppFig_1 – Supplemental material for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism
Supplemental material, SuppFig_1 for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism by Hui Zhong, Chunni Zhu, Yoshihiko Minegishi, Franziska Richter, Sharon Zdunowski, Roland R. Roy, Bryce Vissel, Parag Gad, Yury Gerasimenko, Marie-Francoise Chesselet and V. Reggie Edgerton in Neurorehabilitation and Neural Repair
Supplemental Material
SuppFig_2 – Supplemental material for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism
Supplemental material, SuppFig_2 for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism by Hui Zhong, Chunni Zhu, Yoshihiko Minegishi, Franziska Richter, Sharon Zdunowski, Roland R. Roy, Bryce Vissel, Parag Gad, Yury Gerasimenko, Marie-Francoise Chesselet and V. Reggie Edgerton in Neurorehabilitation and Neural Repair
Supplemental Material
SuppFig_3 – Supplemental material for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism
Supplemental material, SuppFig_3 for Epidural Spinal Cord Stimulation Improves Motor Function in Rats With Chemically Induced Parkinsonism by Hui Zhong, Chunni Zhu, Yoshihiko Minegishi, Franziska Richter, Sharon Zdunowski, Roland R. Roy, Bryce Vissel, Parag Gad, Yury Gerasimenko, Marie-Francoise Chesselet and V. Reggie Edgerton in Neurorehabilitation and Neural Repair
Footnotes
Acknowledgements
We thank Victoria Leon for her technical assistance.
Supplementary material for this article is available on the Neurorehabilitation & Neural Repair website along with the online version of the article.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: PG, RRR, YG, and VE, researchers on the study team hold shareholder interest in NeuroRecovery Technologies and hold certain inventorship rights on intellectual property licensed by The Regents of the University of California to NeuroRecovery Technologies and its subsidiaries. MFC received grants from NIH and Michael J. Fox Foundation during the conduct of the study. She also receives personal fees from Michael J. Fox Foundation outside the submitted works. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by Michael J. Fox Foundation, NIH NS062009-01A1, U01 EB007615 and Dana & Albert R Broccoli Charitable Foundation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.