
Abstract
Background. Cerebrolysin is a neuropeptide preparation with neuroprotective and neurotrophic properties. Our previous study demonstrates that cerebrolysin significantly improves functional recovery in rats after mild traumatic brain injury (mTBI). Objective. To determine histological outcomes associated with therapeutic effects of cerebrolysin on functional recovery after TBI. Methods. In this prospective, randomized, blinded, and placebo-controlled study, adult Wistar rats with mild TBI induced by a closed head impact were randomly assigned to one of the cerebrolysin dose groups (0.8, 2.5, 7.5 mL/kg) or placebo, which were administered 4 hours after TBI and then daily for 10 consecutive days. Functional tests assessed cognitive, behavioral, motor, and neurological performance. Study end point was day 90 after TBI. Brains were processed for histological tissue analyses of astrogliosis, axonal injury, and neurogenesis. Results. Compared with placebo, cerebrolysin significantly reduced amyloid precursor protein accumulation, astrogliosis, and axonal damage in various brain regions and increased the number of neuroblasts and neurogenesis in the dentate gyrus. There was a significant dose effect of cerebrolysin on functional outcomes at 3 months after injury compared with saline treatment. Cerebrolysin at a dose of ⩾0.8 mL/kg significantly improved cognitive function, whereas at a dose of ⩾2.5 mL/kg, cerebrolysin also significantly improved sensorimotor function at various time points. There were significant correlations between multiple histological and functional outcomes 90 days after mTBI. Conclusions. Our findings demonstrate that cerebrolysin reduces astrogliosis and axonal injury and promotes neurogenesis, which may contribute to improved functional recovery in rats with mTBI.
Keywords
Introduction
Traumatic brain injury (TBI) is a leading cause of death and disability. Mild TBI (mTBI) is often unnoticed or misdiagnosed because of missing visible physical signs without obvious tissue lesions in the brain. 1 Although mTBI symptoms typically resolve within days to weeks, more than 15% of patients with mTBI have a measurable cognitive deficit at 1 year following an injury. 2 The pathologies underlying cognitive deficits after mTBI are poorly understood, and most clinical trials involve moderate to severe injury, neglecting efforts to treat mTBI. 3 The failure of clinical trials for TBI may, in part, be a result of heterogeneity of the population of TBI patients and variability in treatment approaches. 4 Another important aspect is that most strategies to date have used drugs in clinical trials targeting a single pathophysiological mechanism that contributes to early cell death. 5
Cerebrolysin is a neuropeptide preparation derived from purified brain proteins with both neuroprotective and neurotrophic properties that target multiple pathways to improve functional recovery after neurological diseases and injuries. 6 Cerebrolysin consists of amino acids (80%) and small (<10 Da) peptides (20%) with mass spectrometry analysis. These small peptides mimic the effect of neurotrophic factors, including ciliary neurotrophic factor, fibroblast growth factor 2, brain-derived neurotrophic factor (BDNF), glial cell line–derived neurotrophic factor (GDNF), nerve growth factor, and insulin-like growth factor (IGF).7,8 Cerebrolysin promotes neurite outgrowth and cholinergic fiber regeneration in vitro. 9 It reduces the blood-brain barrier damage in TBI rats 10 and inflammatory responses in rats with intracranial hemorrhage. 11 Cerebrolysin has beneficial effects on cognitive impairment by reducing amyloid precursor protein (APP) accumulation and promoting the preservation of synaptic terminals in mouse models of Alzheimer disease (AD). 12 Besides these protective effects, cerebrolysin promotes neurogenesis in animal models of AD, 13 stroke,14,15 and TBI.16,17 Cerebrolysin increases neuronal survival in the hippocampus, preserves axonal integrity in the striatum, enhances vascular patency and integrity after moderate TBI, 18 and reduces APP levels and astrogliosis in rats after mTBI. 16 Collectively, these multiple beneficial effects resulting from cerebrolysin treatment may in concert contribute to improvement of neurological outcome after brain injury.
Neurotrophic factors, such as BDNF, IGF-1, and GDNF, are neuroprotective in models of brain and spinal cord injuries. 19 However, neurotrophic factors with high molecular weights do not pass the blood-brain barrier in significant amounts. 20 Cerebrolysin with low molecular weights falls into the category of drugs that have been approved in countries outside of the United States mainly for treatment of cognitive impairment in dementia, stroke, and TBI and mainly in Europe, Asia, and South America. Recent clinical studies have shown the therapeutic value of cerebrolysin by improving functional recovery, decreasing the mortality rate, and increasing the favorable outcome in TBI patients with severe disability 21 as well as improving cognitive function in patients with mTBI. 22 Our previous study demonstrates a dose-related improvement of functional recovery with cerebrolysin in rats after mTBI. 23 The therapeutic mechanisms remain elusive. This prospective, randomized, blinded, placebo-controlled study extended our previous study to investigate histological outcomes associated with therapeutic effects of cerebrolysin on functional recovery, specifically, with a focus on determining long-term histological changes of pathological hallmarks and the correlation with functional outcome parameters.
Materials and Methods
Animal Model of mTBI
mTBI was induced by the impact acceleration model that mimics diffuse axonal injury.23,24 All experimental procedures were carried out in accordance with the NIH Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committee (IACUC#1431) of Henry Ford Hospital. The study was performed in male Wistar rats (417 ± 30 g, 3-4 months old, Charles River Breeding Company, Wilmington, MA). The same animals in this study for histological analyses were used in a recently published behavioral outcome study. 23 Anesthesia was induced with 4% isoflurane and maintained with 1.0% to 1.5% isoflurane in 70% N2O and 30% O2 throughout the surgical period. Rectal temperature was maintained at 37°C ± 0.5°C throughout the surgical procedure using a feedback-regulated water heating system. The skin over the cranial vault was shaved and swabbed with an antiseptic (Betadine) and 70% alcohol. A 2-cm midline incision was made using a scalpel, and the skull was exposed. The stainless steel disk (10 mm in diameter with 3 mm thickness) was coated with cyanoacrylic glue (Krazy Glue) and mounted on the parietal bone midline between Bregma and Lambda, straddling the sagittal suture. The animal was placed prone on the foam bed and secured by strapping surgical tape. Closed-head injury was induced by dropping the cylindrical column of segmented brass (450 g) through the Plexiglas tube from a distance (1 m) onto the disc fixed to the skull vault of the animal. The metal helmet was removed and the incision closed with a sterile 4-0 suture.
Animal Groups and Treatment
We designed a prospective, randomized, placebo-controlled, and blinded study for evaluating the effects of cerebrolysin therapy in rats with mTBI. Study personnel involved in treatment and outcome assessment were blinded. The sample size, 12 rats per group, was prespecified assuming α =.05, a 2-sided test, and possible mortality to ensure an effect size of 1.43 and power of 80%. Randomization schema was generated using nQuery3.0. The randomization was based on a mixed block size. Male Wistar rats (417 ± 30 g, 3-4 months old, Charles River Breeding Company, Wilmington, MA) were randomized to 1 of 3 cerebrolysin doses (0.8, 2.5, or 7.5 mL/kg) or vehicle (saline; n = 12 rats per group). Cerebrolysin was provided by EVER Pharma in preprepared ampoules; each milliliter of cerebrolysin contains 215.2 mg of the active cerebrolysin concentrate in an aqueous solution (EVER Neuro Pharma GmbH, Unterach, Austria). The drug doses were chosen based on our previous studies in TBI 18 and stroke. 17 Treatment with study drug was initiated 4 hours after injury and was administered intraperitoneally once daily for 10 consecutive days. Study medication was prepared by a designated investigator (who was not involved in any other part of this study) according to animal body weight, 1 day prior to treatment based on the randomization scheme. All experimental drugs were prepared in opaque syringes at a constant volume of 3 mL (adjusted with saline) and administered at the scheduled injection times. The first treatment was administered 4 hours after mTBI and repeated once daily for a total of 10 consecutive days. Laboratory personnel who performed mTBI, drug injection, and outcome assessment were blinded to the treatment. Animals were closely monitored for any adverse events after they were enrolled. Cerebrolysin was prepared fresh every day. Age-matched controls with sham surgery without injury were used as shams (n = 12). For labeling proliferating cells, 5′-bromo-2′-deoxyuridine (BrdU, 100 mg/kg) was injected intraperitoneally into rats daily for 7 days, starting 1 day after mTBI. Animals were killed humanely 90 days after mTBI.
Evaluation of Functional Outcomes
Cognitive performance was assessed by using the Morris water maze (MWM) daily for 5 days on all rats at 3 months (days 86-90 after injury). 16 At 1 and 3 months after injury, behavior was assessed by the novel object recognition (NOR) test rating the spontaneous tendency of rodents to spend more time exploring a novel object 25 and by the 3-chamber social interaction test rating the tendency to spend more time interacting with a novel rat versus one they have previously encountered. 26 Motor function was assessed by the adhesive removal test assessing sensorimotor deficits at 1, 7, 14, 21, 30, 60, and 90 days after injury. Neurological functions were assessed by the modified neurological severity score (mNSS), a composite of motor, sensory, reflex, and balance tests at 1, 7, 14, 21, 30, 60, and 90 days after injury. 27 The functional tests were performed by a researcher blinded to the status of group and treatment according to the detailed protocols published in our previous study. 23 We did not report the functional data in this study because we have recently published a behavioral outcome study using the same animals. 23
Evaluation of Histological Outcomes by Immunohistochemistry
Rats were anesthetized intraperitoneally with ketamine (100 mg/kg) and xylazine (10-13 mg/kg) and perfused transcardially with saline solution, followed by 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS), pH 7.4. Rat brains were removed and immersed in 4% paraformaldehyde for 2 to 4 days. Using a rat brain matrix (Activational Systems, Inc, Warren, MI), each forebrain was cut into 2-mm thick coronal blocks for a total of 7 blocks from Bregma 5.2 mm to Bregma −8.8 mm per animal. The tissues were embedded in paraffin and a series of 6-µm-thick slides were cut. Antigen retrieval was performed by boiling sections in 10 mM citrate buffer (pH 6.0) for 10 minutes. After washing with PBS, sections were incubated with 0.3% H2O2 in PBS for 10 minutes, blocked with 1% BSA (bovine serum albumin) containing 0.3% Triton-X 100 at room temperature for 1 hour, and incubated with mouse antidoublecortin (anti-DCX; 1:200, Santa Cruz Biotechnology, Santa Cruz, CA), rabbit polyclonal antiglial fibrillary acidic protein (anti-GFAP; 1:1000, Dako, Glostrup, Denmark), mouse anti-NeuN monoclonal antibody (1:300; MAB 377, Chemicon, Temecula, CA), phosphorylated neurofilament H (1:1000; pNfH, Covance SMI 31R) antibody, and monoclonal mouse anti-APP A4 (1:400; MAB348, Millipore Sigma, Burlington, MA) at 4°C overnight. For negative controls, primary antibodies were omitted. After washing, sections were incubated with biotinylated antimouse, antirabbit, or antichicken antibodies (1:400; Vector Laboratories, Inc, Burlingame, CA) at room temperature for 30 minutes. After an additional washing, sections were incubated with an avidin-biotin-peroxidase system (ABC kit, Vector Laboratories, Inc), visualized with diaminobenzidine (Sigma) and counterstained with hematoxylin. Cell counting and analyses were performed in the cortex (CT), corpus callosum (CC), Cornu Ammonis (CA)1, CA3, dentate gyrus (DG), and thalamus (TH) to assess astrogliosis (GFAP), and axonal injury (APP and phosphorylated neurofilament heavy subunit [pNfH]). Data on neuronal nuclei [NeuN] for mature neurons and neurogenesis (DCX) for immature newborn neurons were collected.
Evaluation of Neurogenesis by Double Immunofluorescent Staining
Double fluorescent staining with NeuN and BrdU was performed to identify newborn mature neurons in the DG. Briefly, after being deparaffinized and rehydrated, brain sections were boiled in 10 mM citric acid buffer (pH 6) for 10 minutes. After washing with PBS, sections were incubated in 2.4 N HCl at 37°C for 20 minutes. Sections were incubated with 1% BSA containing 0.3% Triton-X-100 in PBS. Sections were then incubated with mouse anti-NeuN antibody (1:200; Chemicon) at 4°C overnight. For negative controls, primary antibodies were omitted. FITC-conjugated antimouse antibody (1:400; Jackson ImmunoResearch, West Grove, PA) was added to sections at room temperature for 2 hours. Sections were then incubated with rat anti-BrdU antibody (1:200; Dako) at 4°C overnight. Sections were then incubated with Cy3-conjugated goat antirat antibody (1:400; Jackson ImmunoResearch) at room temperature for 2 hours. Each of the steps was followed by three 5-minute rinses in PBS. Tissue sections were mounted with Vectashield mounting medium (Vector laboratories).
Cell Counting and Quantitation
Cell counting and quantitation were performed by observers blinded to the individual treatment status of the animals. The fields of interest were digitized under a light microscope (Nikon, Eclipse 80i) at a magnification of either 200 or 400 using CoolSNAP color camera (Photometrics) interfaced with MetaMorph image analysis system (Molecular Devices), as described in detail previously. 18 The immunoreactive cells or area of positive staining was calculated and divided by the measured areas and presented as numbers per square millimeter or percentage of area. Astroglial cells were identified by GFAP immunostaining. To maintain consistency with our previous study, 16 we counted GFAP+ cells. Cell counting and area measurements were performed by observers blinded to the individual treatment status of the animals.
Data Analysis
For measurements of APP, NeuN, GFAP, DCX, and pNfH staining, generalized linear mixed models using PROC GLIMMIX (SAS version 9.4) were used with study subject as the random effect and treatment dose/specimen region as fixed effects. First, the model started testing for Treatment × Region interactions. If there was a significant Treatment × Region interaction, we used pairwise dose comparison for each region or a main dose effect at a critical value of 0.05. Bonferroni correction was applied to adjust the 4 dose-group comparisons. The pairwise dose comparison at each region would be considered exploratory if neither the interaction nor the main dose effect was detected. For measurements of DCX and BrdU/NeuN staining, generalized linear models using PROC GENMOD (SAS version 9.4) were used in an approach similar to that described above. The dose effect was detected if adjusted P values were <.05 after Bonferroni correction. Pearson partial correlation with correction for multiple comparisons was used to test correlation between effects of cerebrolysin on each staining with each function at 3 months, controlling for possible dose differences.
Results
In the present study, a total of 60 animals were randomly assigned into 1 of 5 groups, including a sham group and 4 groups of rats with mTBI and treated with cerebrolysin at 0 (saline as placebo), 0.8, 2.5, or 7.5 mL/kg, with 12 animals included in each group. No skull fractures or apneas were observed immediately after impact. In general, all the animals in each group gained weight over time (P < .01). However, there was no Dose × Time interaction or effect on weight. No other adverse effects (ie, death, seizures, dehydration, etc), even at the highest dose of cerebrolysin (7.5 mL/kg), were observed during the 90-day study, which is consistent with the safety profile for this agent in animals and humans. No animals died during the 90-day study.
Cerebrolysin Significantly Reduced Activation of Astrocytes After mTBI
Immunohistochemical staining of GFAP for detecting reactive astrocytes has shown a significantly reduced number of GFAP+ astrocytes for the individual cerebrolysin doses in all brain regions in comparison to placebo (CC, DG, CA1, CA3, TH; Figure 1 and Supplementary Figure S1). In the CT, the higher cerebrolysin doses (2.5 and 7.5 mL/kg) had a significant effect on astrogliosis in comparison to placebo. However, there was no significant difference in effects of cerebrolysin cortical astrogliosis among 3 doses (0.8, 2.5, and 7.5 mL/kg). Pairwise comparison showed a significant superiority of the higher doses (2.5 and 7.5 mL/kg) versus the 0.8-mL/kg dose in the CC and of the 2.5-mL/kg dose over 0.8 mL/kg in the DG. Moderate to strong correlation was observed between astrogliosis and social interaction, and neurogenesis. The correlation with cognitive performance, NOR, and sensorimotor functions (mNSS and adhesive removal) was in the weak to moderate range (Tables 1 and 2).
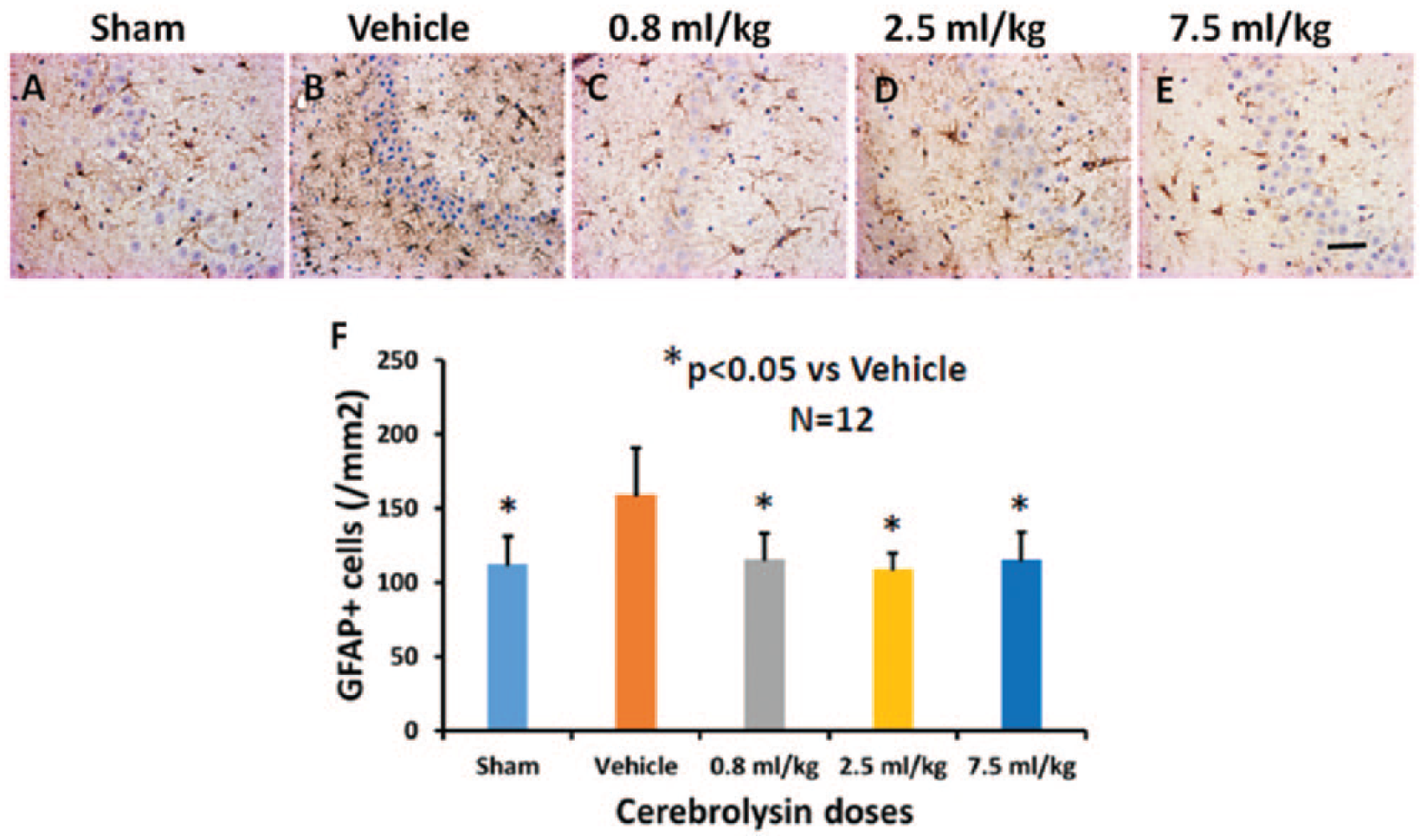
The effects of cerebrolysin on astrocyte activation. GFAP staining was performed to detect activation of astrocytes 90 days after mTBI. Some expression of GFAP was observed in the CA3 region of the hippocampus of sham animals (A). Cerebrolysin treatment at 0.8 mL/kg (C), 2.5 mL/kg (D), and 7.5 mL/kg (E) significantly decreases GFAP+ cells in the CA3 region 90 days after mTBI compared with the vehicle group where prominent astrogliosis exists (B). The data on the number of GFAP+ cells are shown in the bar graph (F). Scale bar = 25 µm (E).
Correlation of the Number of GFAP+ Cells in Each Region With Functional Tests at Day 90 After mTBI. a
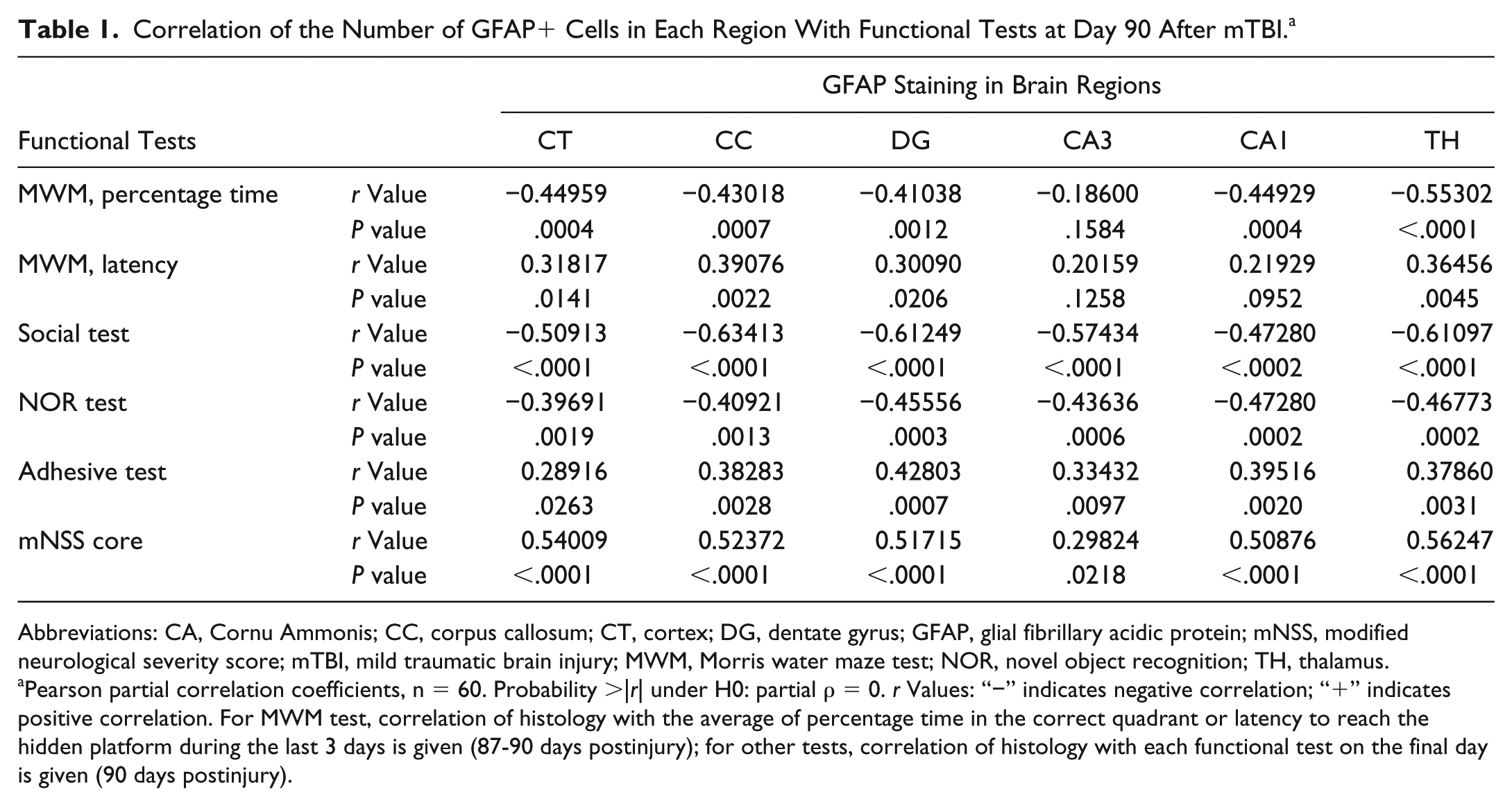
Abbreviations: CA, Cornu Ammonis; CC, corpus callosum; CT, cortex; DG, dentate gyrus; GFAP, glial fibrillary acidic protein; mNSS, modified neurological severity score; mTBI, mild traumatic brain injury; MWM, Morris water maze test; NOR, novel object recognition; TH, thalamus.
Pearson partial correlation coefficients, n = 60. Probability >|r| under H0: partial ρ = 0. r Values: “−” indicates negative correlation; “+” indicates positive correlation. For MWM test, correlation of histology with the average of percentage time in the correct quadrant or latency to reach the hidden platform during the last 3 days is given (87-90 days postinjury); for other tests, correlation of histology with each functional test on the final day is given (90 days postinjury).
Correlation of the Number of DCX+ and BrdU/NeuN+ Cells With Immunostaining in the DG and Functional Tests at Day 90 After mTBI. a
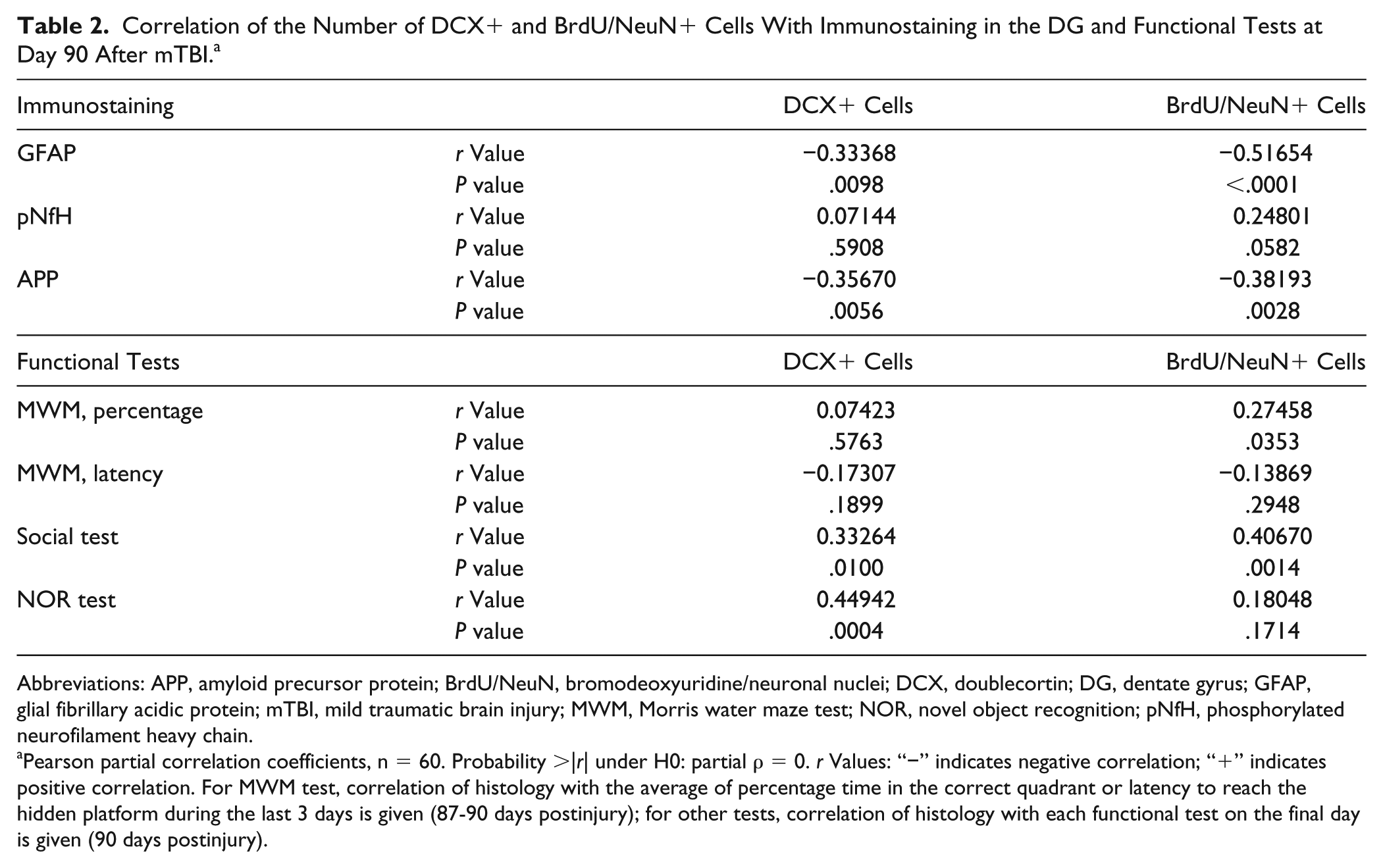
Abbreviations: APP, amyloid precursor protein; BrdU/NeuN, bromodeoxyuridine/neuronal nuclei; DCX, doublecortin; DG, dentate gyrus; GFAP, glial fibrillary acidic protein; mTBI, mild traumatic brain injury; MWM, Morris water maze test; NOR, novel object recognition; pNfH, phosphorylated neurofilament heavy chain.
Pearson partial correlation coefficients, n = 60. Probability >|r| under H0: partial ρ = 0. r Values: “−” indicates negative correlation; “+” indicates positive correlation. For MWM test, correlation of histology with the average of percentage time in the correct quadrant or latency to reach the hidden platform during the last 3 days is given (87-90 days postinjury); for other tests, correlation of histology with each functional test on the final day is given (90 days postinjury).
Cerebrolysin Significantly Reduced Axonal Damage After mTBI
APP staining has shown a significantly reduced APP accumulation for all individual cerebrolysin doses in all brain regions in comparison to placebo (CT, DG, CA1, CA3, Figure 2 and Supplementary Figure S2). The CC was not affected by APP accumulation; no differences were seen between the 5 study groups. Pairwise comparison showed a significant superiority of the 2.5-mL/kg dose versus the 0.8-mL/kg dose in the CA3 region. Moderate to strong correlation was observed between APP accumulation and social interaction. Correlation with cognitive performance, NOR, and sensorimotor functions was in the weak to moderate range (Tables 2 and Supplemental Table 1).
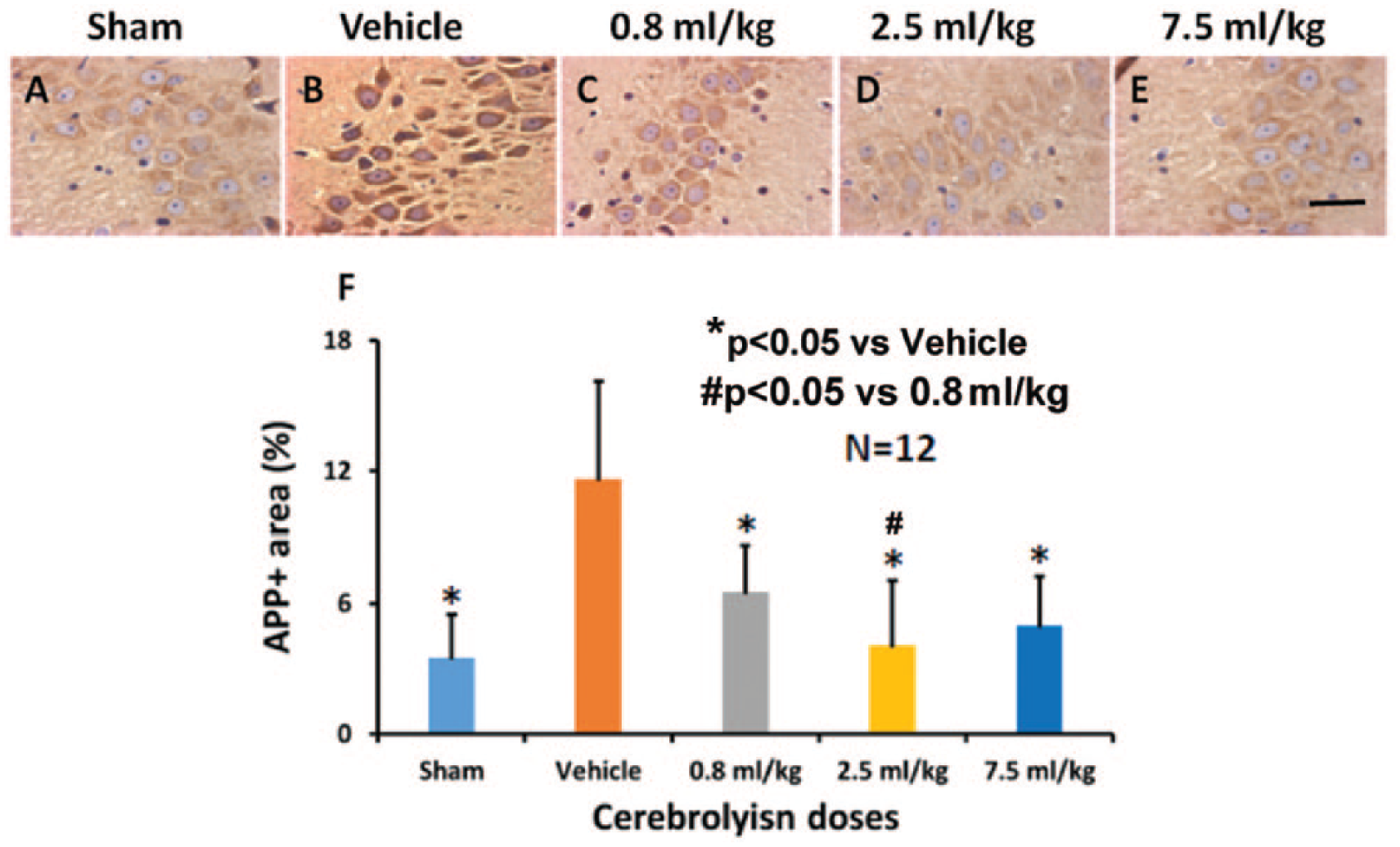
The effects of cerebrolysin on brain APP accumulation. APP is weakly expressed in the CA3 region of sham animals (A). Compared with the vehicle-treated group (B), cerebrolysin treatment at doses of 0.8 mL/kg (C), 2.5 mL/kg (D), and 7.5 mL/kg (E) significantly reduces APP accumulation in the CA3 region 90 days after mTBI. The data on APP+ area are shown in the bar graph (F). Scale bar = 25 µm (E).a
Staining of pNfH has shown a significant increase of pNfH+ neuronal processes for all individual cerebrolysin doses in all brain regions (CT, CC, CA1, CA3, DG, TH) in comparison to placebo (Figure 3 and Supplementary Figure S3). However, the effect of the lowest cerebrolysin doses (0.8 mL/kg) was not significant in the CC. Pairwise comparison showed a significant superiority of the higher doses (2.5 and 7.5 mL/kg) versus the 0.8-mL/kg dose in the CT and CC. Moderate to strong correlation was observed between pNfH+ neuronal processes and both social interaction and neurological functions. Weak to moderate correlation was observed with cognitive performance (Table 2 and Supplemental Table 2).
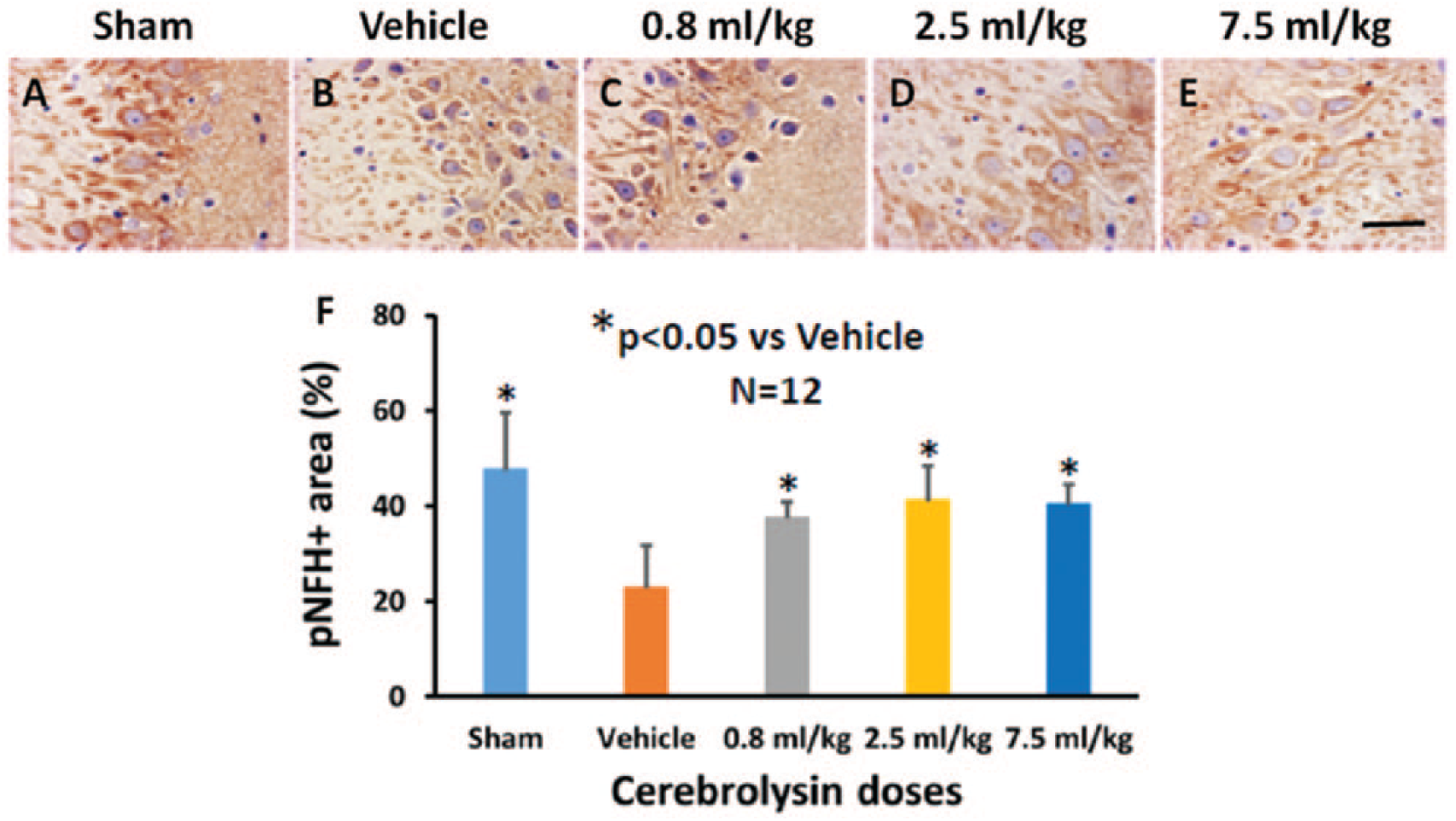
The effects of cerebrolysin on percentage of pNfH+ areas. Compared with brain regions of sham animals (A), there is a significant decrease in percentage of pNfH+ areas in the CA3 region from the vehicle-treated group (B). Cerebrolysin treatment at doses of 0.8 mL/kg (C), 2.5 mL/kg (D), and 7.5 mL/kg (E) significantly increases the percentage of pNfH+ areas in the CA3 region 90 days after mTBI. The data on pNfH+ areas are shown in the bar graph (F). Scale bar = 25 µm (E).
Cerebrolysin Significantly Increased the Number of Neuroblasts and Mature Neurons in the DG After mTBI
Immunohistochemical staining of DCX in the DG has shown a significant increase in the number of neuroblasts for the individual cerebrolysin doses in comparison to placebo (Figure 4). Pairwise comparison showed no significant differences between individual cerebrolysin doses. A weak to moderate correlation was observed between neuroblasts and behavior (social interaction and NOR), APP accumulation, and astrogliosis. Double immunostaining of BrdU/NeuN in the DG has shown a significant increase in the number of new mature neurons in comparison to placebo overall and for the individual cerebrolysin doses (Figure 4). Pairwise comparison showed no significant differences between individual cerebrolysin doses. A moderate correlation was observed with GFAP and a weak to moderate correlation with APP accumulation, cognitive function, and social interaction (Table 2). This mTBI did not cause neuronal loss in any brain region with NeuN staining (Supplementary Figure S4).
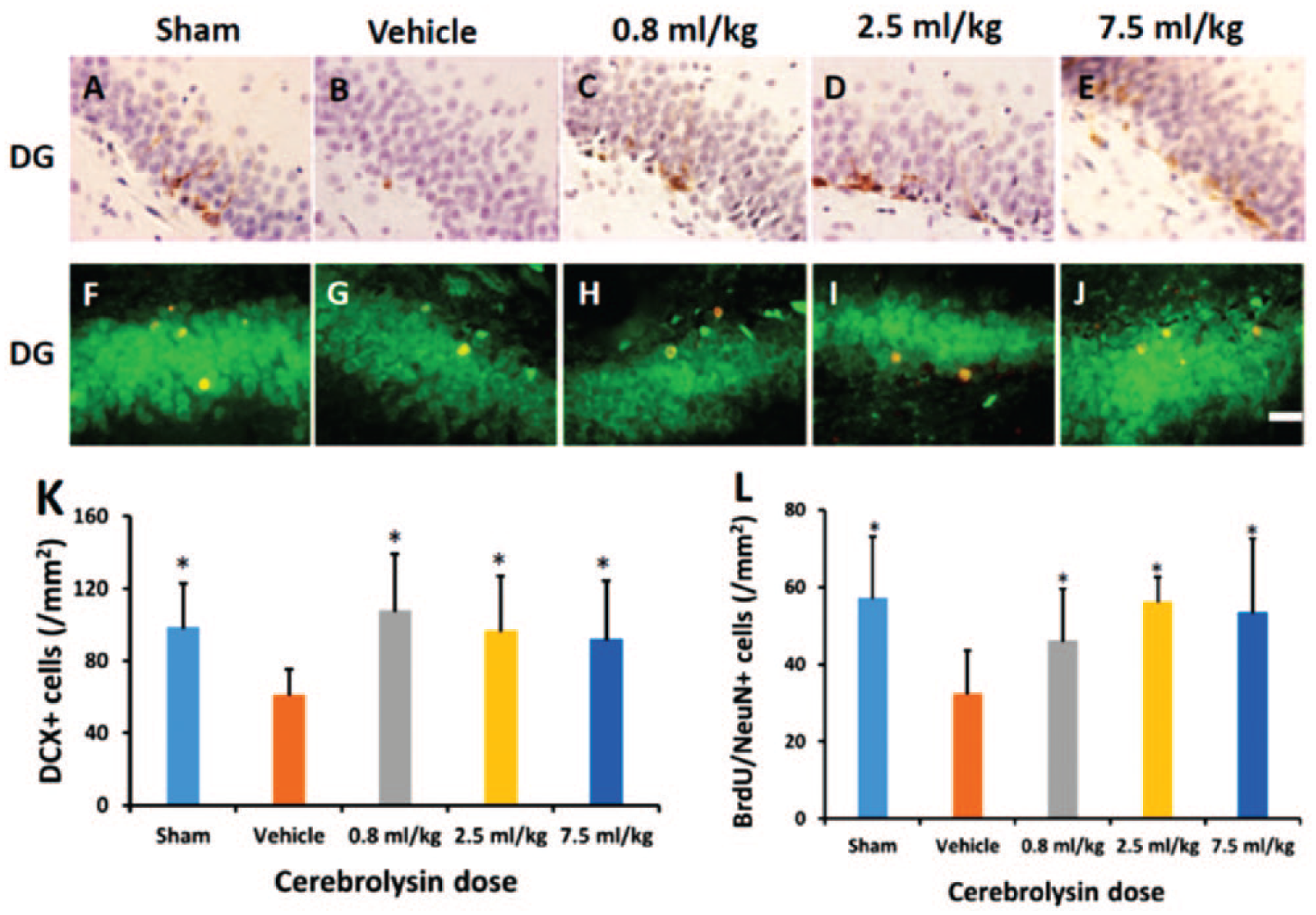
The effects of cerebrolysin on the number of DCX-expressing neuroblasts and BrdU/NeuN+ newborn mature neurons in the DG. DCX staining was used to identify neuroblasts in the DG, (A-E). Compared with the vehicle-treated group (B), cerebrolysin treatment (C-E) significantly increases DCX+ neuroblasts in the DG 90 days after mTBI, which is comparable to sham level (A). The bar graph (K) shows the number of DCX+ neuroblasts. Compared with the vehicle-treated group (G), cerebrolysin treatment (H-J) significantly increased newborn mature neurons identified with BrdU/NeuN double immunofluorescent staining (merged in yellow) in the DG, at 90 days postinjury. The bar graph (L) shows the number of BrdU/NeuN+ newborn mature neurons. Scale bar = 25 µm.
Discussion
In the previous study with a delayed and long-term treatment, cerebrolysin was given 24 hours postinjury for 28 days in rats after mTBI. 16 In the present study, with acute and short-term treatment, cerebrolysin was given 4 hours postinjury for 10 days in rats after mTBI. More important, this study was a prospective, randomized, blinded, and placebo-controlled preclinical experiment, with the intention to optimize protocols for future clinical studies. To maximize the translational power of the study, we followed current guidelines on the design and reporting of experimental animal studies. Our data of an absence of a difference in NeuN staining between sham and vehicle groups are consistent with data on rats subjected to a mild TBI resulting from a 450 g × 1 m impact that demonstrated no significant neuronal death. 28
Our data indicate that compared with placebo, cerebrolysin significantly increased neurogenesis and the number of neuroblasts in the DG, and reduced APP accumulation, astrogliosis, and axonal damage in various brain regions. There were significant correlations between multiple histological and functional outcomes 90 days after mTBI, as detected by Pearson partial correlation analyses. Higher doses (⩾2.5 mL/kg) resulted in better functional recovery 23 ; however, differences in histological outcomes between these effective doses were not significant. These data indicate that in addition to its beneficial histological effects, cerebrolysin may also alter other mechanisms at signaling pathways and electrophysiological activity we did not investigate for improvement of functional recovery in rats after mTBI.
Cerebrolysin Attenuates Activation of Astrocytes After mTBI
mTBI is a poorly understood health problem and can result in short- and long-term cognitive, behavioral, and emotional impairments without obvious gross brain damage. 29 In patients with mild and moderate TBI, GFAP (biomarker for activation of astrocytes) was detectable in sera within 1 hour after concussion, 30 and inflammation may persist for many years after a single TBI event. 31 Reactive gliosis has been described as constitutive, graded, multistage, and evolutionary conserved defensive astroglial reaction. 32 Whereas astrocytes are vital for the normal functioning of the brain, 33 chronic reactive astrocytes have detrimental effects also and might negatively influence long-term cognitive performance 34 and immature neurons. 35 In the current study, there is a strong correlation of cerebrolysin-reduced astrogliosis and improved neurogenesis. Increased neurogenesis may be related to the reduced astrogliosis induced by cerebrolysin. Our previous data indicate that higher doses of cerebrolysin (2.5 and 7.5 vs 0.8 mL/kg) provide better efficacy in terms of improving cognitive performance, 23 which is at least partially related to cerebrolysin-reduced astrogliosis in all brain regions and enhanced neurogenesis in the DG observed in this study.
Cerebrolysin Reduces Axonal Injury After mTBI: APP Accumulation
The soluble α form of APP (sAPPα) is neuroprotective and promotes neuronal plasticity. 36 Intracerebroventricular administration of sAPPα in rats at 30 minutes after severe diffuse TBI significantly reduced axonal injury and apoptosis and improved motor and cognitive outcomes. 36 However, alternative processing or overexpression of APP might result in detrimental accumulation of APP, which occurs rapidly after TBI and persists in both animals and humans.37-40 In an AD model, cerebrolysin decreased amyloid β (Aβ) load, which was accompanied by an amelioration of cognitive performance and a reduced synaptic pathology. 12 In our study, APP accumulation was detected mainly in the soma of neurons 90 days after mTBI, which may be one of the reasons why APP accumulation was not observed in the CC where axons are the main component. Our data indicate that cerebrolysin at all doses significantly reduces APP accumulation and shows a significant dose-response difference, suggesting that cerebrolysin at the low dose (0.8 mL/kg) is potent enough to reduce APP level in the brain after mTBI. However, the specific pathway remains elusive but might include activation of α secretase and/or promotion of APP processing toward neuroprotective sAPPα.
Cerebrolysin Reduces Axonal Injury After mTBI: pNfH Staining
Neurofilament phosphorylation has been intimately tied to myelination and axonal transport. 41 However, in sera/cerebrospinal fluid (CSF), pNfH has been identified as a possible early biomarker for measuring the extent of axonal injury in humans. 42 Increased serum levels were also observed in rats at 1 and 3 days after mTBI induced by rotational impact 43 and in moderate and severe focal TBI induced by controlled cortical impact but not in mild focal TBI. 44 In the present study, we did not measure the pNfH level in serum or CSF but found that pNfH+ staining was reduced in brain regions 90 days after mTBI. More important, cerebrolysin significantly increased the percentage of the pNfH+ area in multiple brain regions compared with the saline group, suggesting amelioration of axonal injury on treatment. To better monitor the efficacy of cerebrolysin, measurements of serum pNfH after mTBI and the effects of cerebrolysin treatment on brain and serum pNfH at multiple early and late time points are warranted.
Cerebrolysin Stimulates Neurogenesis in the DG After mTBI
Adult mammalian brains harbor neural stem/progenitor cells, 45 which have the potential to become functional neurons and contribute to cognitive recovery after TBI.46,47 A large proportion of newly generated cells in rodent DG die within 1 month after TBI.48,49 The majority of BrdU+ cells, which survive for 10 weeks, become dentate granule neurons in TBI rats. 50 Some of these BrdU+ cells are colocalized with synaptophysin, suggesting that they may receive synaptic input and associate with cognitive recovery. 50 In the present study, cerebrolysin increased the number of newborn immature (DCX+ neuroblasts) and mature (BrdU/NeuN+ cells) neurons in the DG examined at 90 days after mTBI. Furthermore, the Pearson’s correlation analyses (Table 2) showed a significant, positive correlation of the number of DCX+ neuroblasts with social and NOR tests as well as a significant, positive correlation of the number of newborn mature neurons (BrdU/NeuN+ cells) with MWM (percentage time spent in the correct quadrant) and social tests. The mechanisms underlying cerebrolysin-enhanced neurogenesis after TBI largely remain elusive. Our previous studies indicate that the Sonic hedgehog signaling pathway mediates cerebrolysin-enhanced neurogenesis, oligodendrogenesis, and axonal remodeling in the peri-infarct area after stroke, 14 and the PI3K/Akt pathway mediates cerebrolysin-augmented proliferation, differentiation, and migration of adult neural progenitor cells in the subventricular zone after stroke. 15 A recent study demonstrates that cerebrolysin is capable of protecting the grafted neural stem cells in an APP transgenic mouse model of AD and increases BDNF. 51 We did not measure the effect of cerebrolysin on BDNF in this study. Further investigation of cerebrolysin on BDNF in injured brain is warranted because of the importance of BDNF in the neuroprotection and regulation of neurogenesis after TBI. 52
Limitations
The adult brain is capable of initiating widespread structural and functional reorganization in response to brain injury and therapeutic treatment.53,54 We do not exclude the possibility that other brain regions also participate in the functional recovery induced by cerebrolysin treatment after mTBI. In addition, cerebrolysin may target multiple pathways to improve functional recovery after neurological diseases and injuries.6,55 Functional benefits from cerebrolysin could be attributed to many other mechanisms, including synaptogenesis, oligodendrogenesis, and dendritic remodeling, which were not investigated in this study. Further investigation of the expected time course of gliosis and neurogenesis post-TBI is warranted for better understanding of the rationale for treatment timing and duration. Based on the demographic data, TBI primarily occurs in young, adult male civilians and deployed military. 56 However, controversy exists regarding the role of sex in TBI outcome and response to TBI treatments.57,58 It is, therefore, warranted to determine efficacy of cerebrolysin in female rats subjected to TBI. In addition, although we did not observe seizure in rats with mild TBI treated with cerebrolysin in our study, it should be noted that drugs that promote plasticity and neurite outgrowth may increase susceptibility to seizure.59,60
Conclusions
Treatment with cerebrolysin over 10 days at doses of 0.8 to 7.5 mL/kg and initiated 4 hours after injury had beneficial effect on the pathophysiological hallmarks of mTBI in rats. Cerebrolysin significantly reduced the astrocyte activation (GFAP) and axonal damage (reduced APP and increased pNfH) and increased neurogenesis (DCX, BrdU/NeuN). Our data in Tables 1 and 2 show that APP accumulation/axonal injury/GFAP activation in CT and the hippocampus as well as neurogenesis in DG are significantly correlated to both social behavior (the 3-chamber social interaction test) and cognition (MWM and NOR). These data indicate that the impact acceleration produces a diffuse injury as evidenced by different immunostainings in various brain regions without noticeable cortical and hippocampal cell loss. The diffuse injury leads to social and cognitive deficits. However, there is some difference in P values and correlation coefficient r values between social behavior and cognition with different immunostainings, with a tight correlation for social behavior rather than cognition. The present study suggests that the social test may be more sensitive to mTBI than cognitive tests. However, additional studies are warranted. The significant correlation between histology and functional outcome found in this study suggests that the structural and plasticity effects stimulated by cerebrolysin may in concert drive functional improvement. Effects of the individual dosage groups were comparable; however, they were most pronounced in the 2.5-mL/kg group, corresponding to a human dose of 30 mL. Our data on functional recovery reported in a previous study 23 and histopathology in this study together with previous preclinical and clinical studies support the efficacy of cerebrolysin for the treatment of mTBI.
Supplemental Material
Supplemental_material – Supplemental material for Cerebrolysin Reduces Astrogliosis and Axonal Injury and Enhances Neurogenesis in Rats After Closed Head Injury
Supplemental material, Supplemental_material for Cerebrolysin Reduces Astrogliosis and Axonal Injury and Enhances Neurogenesis in Rats After Closed Head Injury by Yanlu Zhang, Michael Chopp, Zheng Gang Zhang, Yi Zhang, Li Zhang, Mei Lu, Talan Zhang, Stefan Winter, Edith Doppler, Hemma Brandstäetter, Asim Mahmood and Ye Xiong in Neurorehabilitation and Neural Repair
Footnotes
Acknowledgements
The authors would like to thank Susan MacPhee-Gray for editorial assistance.
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. The authors disclose receipt of the following financial support for the research, authorship, and/or publication of this article: Yanlu Zhang and Ye Xiong had some salary and travel to meetings supported by EVER Neuro Pharma GmbH. Mei Lu and Talan Zhang received some salary support by EVER Neuro Pharma GmbH for statistical analyses. Dr Chopp received travel support from EVER Neuro Pharma GmbH for presenting data at scientific meetings.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was funded by EVER Neuro Pharma GmbH.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.