
Abstract
Background. Cortical electrical stimulation of the motor cortex in combination with rehabilitative training (CS/RT) has been shown to enhance motor recovery in animal models of focal cortical stroke, yet in clinical trials, the effects are much less robust. The variability of stroke location in human patient populations that include both cortical and subcortical brain regions may contribute to the failure to find consistent effects clinically. Objective. This study sought to determine whether infarct location influences the enhanced motor recovery previously observed in response to CS/RT. The efficacy of CS/RT to promote improvements in motor function was examined in 2 different rat models of stroke that varied the amount and location of cortical and subcortical damage. Methods. Ischemic infarctions were induced by injecting the vasoconstricting peptide endothelin-1 either (1) onto the middle cerebral artery (MCA) producing damage to the frontal cortex and lateral striatum or (2) into a subcortical region producing damage to the posterior thalamus and internal capsule (subcortical capsular ischemic injury [SCII]). Daily CS/RT or RT alone was then given for 20 days, during which time performance on a skilled reaching task was assessed. Results. Animals with MCA occlusion infarctions exhibited enhanced improvements on a skilled reaching task in response to CS/RT relative to RT alone. No such enhancement was observed in animals with SCII infarctions across the 20 days of treatment. Conclusions. The efficacy of CS for enhancing motor recovery after stroke may depend in part on the extent and location of the ischemic infarct.
Keywords
Introduction
The combination of epidural electrical stimulation of the motor cortex and motor rehabilitation (CS/RT) has been demonstrated to enhance motor recovery in preclinical animal studies of focal cortical ischemia.1-7 This improved motor function is accompanied by increases in synapse density 2 and enhanced synaptic responses 7 in the perilesional cortex. In addition, CS/RT also results in an expansion and reorganization of ICMS-derived forelimb movement representations in the ipsilesional motor cortex beyond that observed with RT alone.4-6 Despite the robust effects in animal studies, the efficacy of CS/RT for ameliorating poststroke motor impairments in clinical studies has proven to be inconsistent. A recent large-scale clinical trial failed to demonstrate any significant improvements in motor function in chronic stroke patients 4 weeks post-CS/RT treatment. 8 In this same study, a positive treatment response was, however, observed in a subset of patients from whom movements could be elicited in response to epidural electrical stimulation. The efficacy of CS as a stroke treatment may thus depend on the functional integrity of residual neural tissue, including the corticospinal tract. 9
The nature and persistence of functional deficits after stroke can vary because of vast differences in infarct size and location. 10 Preclinical animal studies of stroke have predominantly used experimental models that primarily induce cortical damage.11-14 Clinically, however, damage often includes or may even be restricted to subcortical structures.10-17 Indeed, enduring motor impairments can manifest as a result of damage to subcortical strokes, including descending motor tracts. 18 Damage within subcortical white matter can result in cortical deafferentation, causing dysfunction within widespread cortical areas outside of the infarction.19,20 Small infarctions within the internal capsule, for example, result in severe motor impairments and poor motor recovery.21-23
Lesion location–dependent differences in the amount of motor recovery and cortical reorganization raise the possibility that the efficacy of any given treatment may depend on the specific pattern of brain damage in individual patients. The success of CS/RT for enhancing motor recovery in animal studies may be a result of the fact that the stroke models used have targeted cortical rather than subcortical areas. In the present study, we tested the efficacy of CS/RT to promote improvements in motor function in 2 different rat models of stroke, producing 2 different patterns of brain injury.
Methods
Animals
A total of 40 adult male Long-Evans hooded rats (350-420 g) were pair housed (2 animals per cage) in standard laboratory cages on a 12:12 hour light-dark cycle in the University of Florida’s Communicore Research Building vivarium. All experimentation occurred during the light cycle. Rats were given Lab Diet 5001 (PMI Feeds, St Louis, MO) and water ad libitum and were cared for in accordance with the National Institutes Health Guide for the Care and Use of Laboratory Animals and with the approval of the University of Florida’s Institutional Animal Care and Use Committee.
Reach Training
The single-pellet reaching task was used as previously described.4,5,24 Animals were trained to retrieve food pellets (45 mg; Bio-Serv) with their preferred limb for approximately 2 weeks (20 min/d). A successful reach was scored when the animals grasped the food pellet and brought it to their mouth without dropping it. The percentage of successful reaches [(Number of successful retrievals/Total number of reaches) × 100]—that is, reaching accuracy—was then calculated. All training sessions were videorecorded. Animals were sorted by prelesion performance to create groups with comparable baseline levels of reaching accuracy.
Infarction
Ischemic damage was induced by injecting the vasoactive peptide endothelin-1 either onto the proximal branches of the middle cerebral artery (MCA) along the lateral aspect of the frontal cortex or into the subcortical territory within and surrounding the internal capsule, as previously described.11,12,25 The hemisphere contralateral to each animal’s preferred reaching paw was targeted for injury. Animals were anesthetized with ketamine hydrochloride (70 mg/kg, intraperitoneally [ip]) and xylazine (5 mg/kg, ip), with supplemental isofluorane (0.15%) and ketamine (20 mg/kg, ip). Under sterile conditions, a midline incision was made and burr holes created over the injection sites. Endothelin-1 (0.2 µg/µL American Peptide, Sunnyvale) was delivered by the Nanolitre injection system (World Precision Instruments, Sarasota, FL) using a SYS-Micro 4 Controller (World Precision Instruments, Sarasota, FL). Stereotaxic coordinates of the injection site for MCA occlusion (MCAo) were anteroposterior, +0.9 mm; mediolateral, −5.2mm; and dorsoventral, −8.6 mm, with respect to bregma, 25 whereas for subcortical capsular ischemic injury (SCII), they were anteroposterior, −3.0 mm; mediolateral, −3.0 mm; and dorsoventral, −7.0 mm, with respect to bregma.11,12 Doses of endothelin-1 were 3 µL for MCAo (240 pMol dissolved in 0.9% sterile saline) and1 µL for capsular injury (80 pMol dissolved in 0.9% sterile saline).
Cortical Electrode Implantation
Directly following MCAo or SCII (32 animals total), 9-pin electrode carriages (ABS Plug, Ginder Scientific Inc, Ottawa, Canada) were implanted over the motor cortex in the injured hemisphere. 4 The surface electrode was placed over the exposed cortex between 1 mm posterior to 5 mm anterior to bregma and 0.5 to 5.5 mm lateral to midline. A return lead was fixed to the skull posterior to lambda and the craniotomy filled with gel foam. Both the electrode and gel foam were covered in nonexothermic PolyWave dental acrylic and cured with brief ultraviolet light. The electrode was fixed to skull screws with standard dental acrylic, and the animals were given warm ringers solution (4 cc; subcutaneously [sc]) and metacam (0.10 mg/kg; sc).
Movement Thresholds (MTs)
Three days after implantation, all animals had their individual CS MTs determined and then began motor rehabilitation. MTs were assessed on postlesion training days 1, 10, and 19. MTs were defined for each animal as the minimum current to cause an involuntary motor response. The animals were placed into a transparent cylinder and observed while 3-s trains of 1-ms, 100-Hz monopolar cathodal pulses were given. The current was gradually increased by 5% increments until a movement of the contralateral forelimb could be clearly detected. CS MTs for all animals were identified by one investigator who was blinded to the treatment groups. Care was taken during CS MT testing because preliminary studies found that intensities of continuous CS set at greater than 50% threshold resulted in undesired evoked movements during CS/RT.
Cortical Electrical Stimulation and Rehabilitative Training
CS/RT or RT (20 min/d) was initiated 3 days after surgery and continued for 20 days. CS during rehabilitation training was then delivered at 50% of each animal’s CS MT. RT was performed using the single-pellet reaching task, and all sessions were videorecorded for analysis of reaching accuracy and reach attempts. Animals receiving CS/RT were stimulated via the Vertis Stimulation System during these sessions. The cortical electrode was connected to a remote stimulator controlled by a personal computer. CS/RT was delivered as continuous monopolar cathodal stimulation with a frequency of 100 Hz and a current intensity dictated by each animal’s MT. Each pulse was biphasic, charge balanced, and asymmetric consisting of a square phase lasting 100 ± 10 µs and a decaying exponential phase lasting ~9900 ± 10 µs. Half of the animals that received MCAo were given RT alone (MCAo-RT; n = 8), whereas the other half received the combination of CS/RT (MCAo-CS/RT; n = 8). Half of the animals that received SCII were given RT alone (SCII-RT; n = 8), whereas the other half received CS/RT (SCII-CS/RT; n = 8). A group of animals with no injury were included as healthy controls (n = 8) and were trained on the same task on all days.
Histology and Lesion Verification
Following the rehabilitation, animals were given pentobarbital and then transcardially perfused with 0.1 M sodium phosphate buffer followed by 4% paraformaldehyde solution in 0.1 M sodium phosphate buffer. Serial 50-µm coronal sections were taken using a microtome; 10 sections spaced 600 µm apart and spanning approximately 2.7 mm anterior and 3.3 mm posterior to bregma were sampled for lesion verification. The sampled sections were stained with toluidine blue and digitally scanned (Epson Perfection V500 Photo Scanner, Long Beach, CA). The area of spared tissue was traced using Image J software, 26 and cortical volumes were estimated with Cavalieri’s unbiased estimator method 27 :
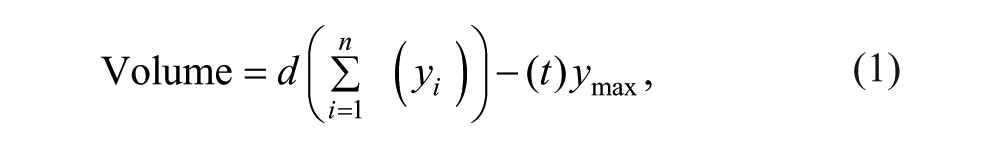
where yi is the cross-sectional area of the ith section through the morphometric region, d is the distance between sections (600 µm), and n is the total number of sections (12). ymax Is the maximum value for the area of 1 section, and t is the section thickness (50 µm); their product was subtracted as a correction. Estimated volumes were analyzed as the percentage affected to unaffected side [(Volume lesioned hemisphere/Volume nonlesioned hemisphere) × 100] to account for individual differences. Each group contained 8 animals.
Data Analysis
Skilled reaching performance was assessed by measuring the percentage change in both number of reaching attempts and reaching accuracy across the 20 days of training. For each animal, the percentage was calculated by subtracting the average of the first 5 rehabilitation days from the average of the final 5 rehabilitation days, then normalizing to preinjury values: [(Days 16 to 20 − Days 1 to 5)/Preinjury] × 100. Motor performance and CS MTs were analyzed using 1-way ANOVA for percentage changes or 2-way repeated-measures ANOVA, and planned mean comparisons across treatment condition within each lesion model were conducted using Fisher’s Protected Least Significant Difference (FPLSD). Spared tissue estimates were analyzed using 1-way ANOVA to assess differences across treatment conditions, and planned mean comparisons across treatment condition within each lesion model were conducted using FPLSD. Data are presented as mean ± SE, and significance was set at P < .05.
Results
Reaching Attempts
A 2-Way repeated-measures ANOVA for training days 1, 5, 10, 15, and 20 for the number of attempts (Figure 1A) revealed a significant Condition × Training Day interaction [F(20, 175) = 2.019; P = .0085] as well as a significant effect of condition [F(4, 35) = 12.62; P < .0001] and training day [F(5, 175) = 23.38; P < .0001]. Planned mean comparisons were used to assess differences between CS/RT and RT within each lesion model. No significant differences between MCAo+RT and MCAo+CS/RT conditions on number of reach attempts were detected on any of the training days: preinjury (P = .76; FPLSD), day 1 (P = .94; FPLSD), day 5 (P = .99; FPLSD), day 10 (P = .25; FPLSD), day 15 (P = .86; FPLSD), and day 20 (P = .56; FPLSD). Animals given SCII+RT and SCII+CS/RT did not significantly differ in the number of reach attempts on any of the training days: preinjury (P = .62; FPLSD), day 1 (P = .81; FPLSD), day 5 (P = .80; FPLSD), day 10 (P = .36; FPLSD), day 15 (P = .61; FPLSD), and day 20 (P = .66; FPLSD).
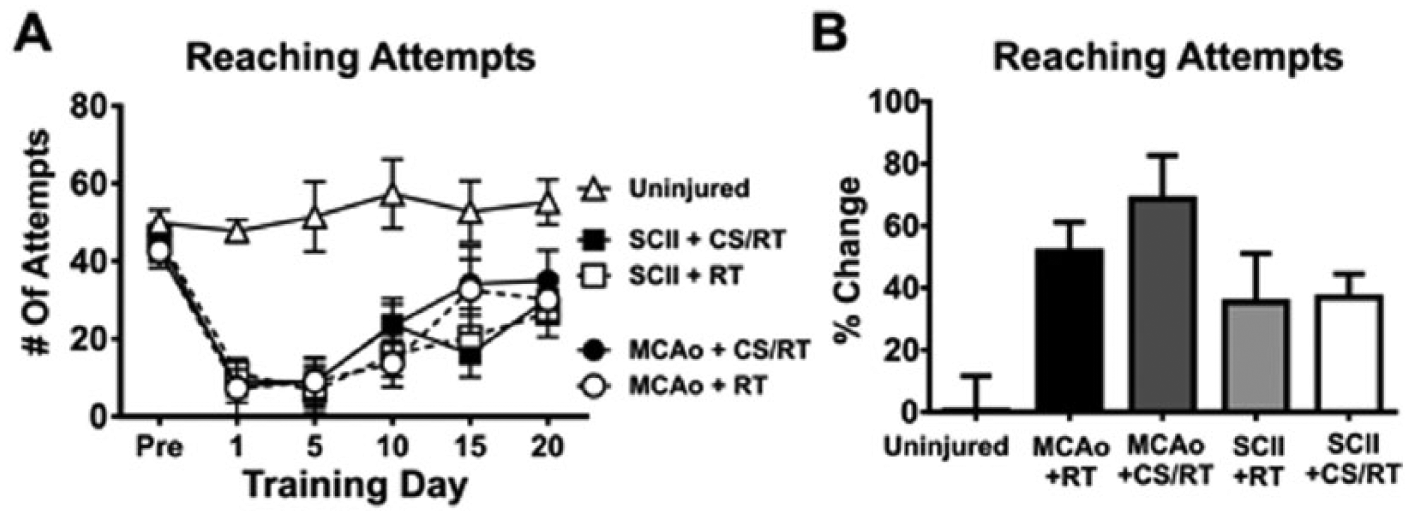
Number of reaching attempts during preinjury and at 1, 5, 10, 15, and 20 days of rehabilitative training (RT) or cortical stimulation with RT (CS/RT). A. Line graph comparing the number of reach attempts during preinjury and RT. No significant differences were detected in the number of reach attempts between RT and CS/RT treatments for animals given either middle cerebral artery occlusion (MCAo) or subcortical capsular ischemic injury (SCII) during preinjury or rehabilitation days 1, 5, 10, 15, and 20. B. Histogram comparing percentage change in the number of reach attempts during preinjury and RT. No significant differences were detected in the percentage change in the number of reach attempts between RT and CS/RT treatments for animals given either MCAo or SCII.
For each animal, a percentage change in the number of reach attempts was calculated by subtracting the average of the first 5 rehabilitation days from the average of the final 5 rehabilitation days then normalizing to each preinjury number of attempts: [(Days 16 to 20 − Days 1 to 5)/Preinjury] × 100 (Figure 1B). A 1-way ANOVA for percentage change of reach attempts detected a significant overall difference between groups [F(4, 35) = 4.46; P = .0051]. Animals given MCAo+RT and MCAo+CS/RT did not differ in percentage change in the number of reach attempts (P = .32; FPLSD). Similarly, animals given SCII+RT and SCII+CS/RT did not differ in percentage change in the number of reach attempts (P = .93; FPLSD).
Reaching Accuracy
The percentage reach accuracy of the animals was analyzed to assess their ability to produce skilled forelimb movements during RT. A 2-way repeated-measures ANOVA for training days 1, 5, 10, 15, and 20 on reaching accuracy (Figure 2A) revealed a significant Condition × Training Day interaction [F(20, 175) = 4.18; P < .0001] as well as a significant effect of condition [F(4, 35) = 18.29; P < .0001] and training day [F(5, 175) = 46.47; P < .0001]. Planned mean comparisons were used to assess differences between CS/RT and RT within each lesion model. Animals given MCAo+RT and MCAo+CS/RT did not significantly differ in reaching accuracy on the following training days: preinjury (P = .91; FPLSD), day 1 (P > .99; FPLSD), day 5 (P = .58; FPLSD), day 10 (P = .31; FPLSD), or day 15 (P = .074; FPLSD). In contrast, the MCAo+CS/RT condition exhibited significantly greater reaching accuracy relative to the MCAo+RT condition on day 20 (P = .0090; FPLSD). Animals given SCII+RT and SCII+CS/RT did not significantly differ in reaching accuracy on any of the training days: preinjury (P = .82; FPLSD), day 1 (P = .77; FPLSD), day 5 (P = .65; FPLSD), day 10 (P = .18; FPLSD), day 15 (P = .51; FPLSD), and day 20 (P = .83; FPLSD).
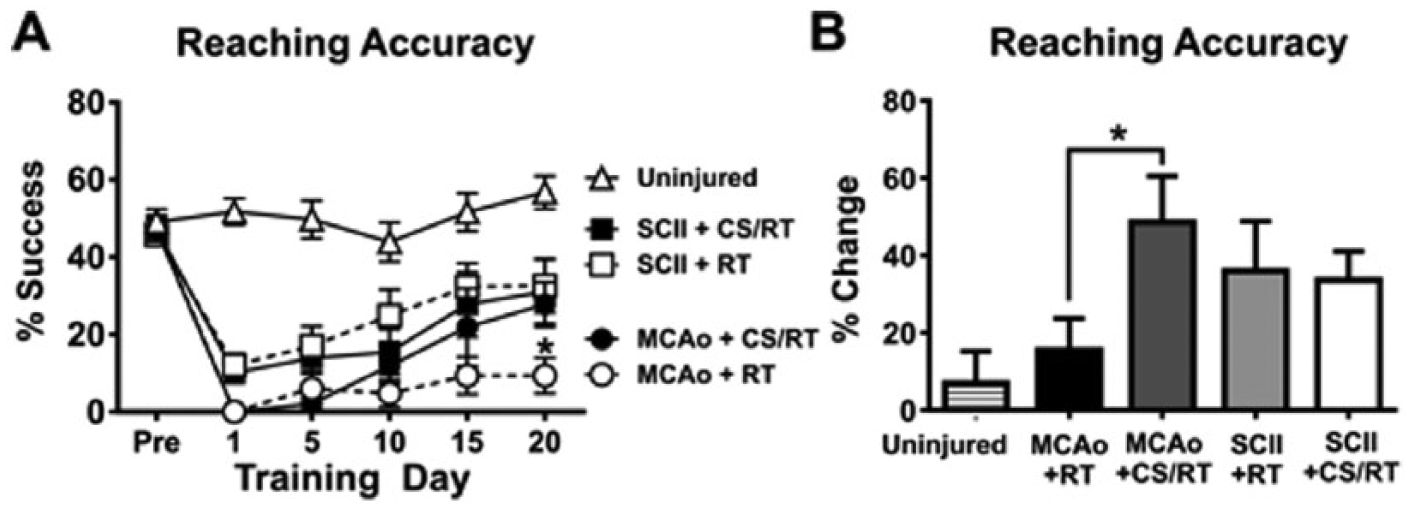
Percentage successful reaching accuracy during preinjury and at 1, 5, 10, 15, and 20 days of rehabilitative training (RT) or cortical stimulation with RT (CS/RT). A. Line graph comparing reaching accuracy during preinjury and RT for animals given either middle cerebral artery occlusion (MCAo) or subcortical capsular ischemic injury (SCII). No significant differences were detected in reaching accuracy between animals given SCII+RT and SCII+CS/RT. Animals given MCAo+CS/RT had significantly greater reaching accuracy on rehabilitation day 20 (*P < .05), whereas nonsignificant differences were detected between these groups at the other time points. B. Histogram comparing percentage change in reaching accuracy during preinjury and RT. Animals given MCAo+CS/RT exhibited a significantly greater percentage increase in reaching accuracy relative to animals given MCAo+RT (*P < .05), whereas this difference was not observed in SCII animals.
A separate 2-way repeated-measures ANOVA for training days 1, 5, 10, 15, and 20 on reaching accuracy revealed a significant Lesion type × Training day interaction [F(10, 185) = 6.77; P < .0001] as well as a significant effect of lesion type [F (2, 37) = 35.87; P < .0001] and training day [F (5, 185) = 29.14; P < .0001]. Planned mean comparisons comparing MCAo and SCII models detected that animals given SCII exhibited significantly greater reaching accuracy relative to animals given MCAo on rehabilitation days 1 (P = .0264; FPLSD), 5 (P = .022; FPLSD), 10 (P = .017; FPLSD), 15 (P = .0039; FPLSD), and 20 (P = .0077; FPLSD).
For each animal, a percentage change in reaching accuracy was calculated by subtracting the average of the first 5 rehabilitation days from the average of the final 5 rehabilitation days then normalizing to each preinjury accuracy: [(Days 16 to 20 − Days 1 to 5)/Preinjury] × 100. A 1-way ANOVA for percentage improvement detected a significant overall main effect of condition [F(4, 35) = 2.83; P = .039]. Planned mean comparisons comparing CS/RT and RT within each lesion model (Figure 2B) detected that the MCAo+CS/RT animals exhibited a significantly greater percentage change in reaching accuracy relative to animals given MCAo+RT (P = .024; FPLSD). In contrast, no difference was detected between animals given SCII+RT and SCII+CS/RT (P = .86; FPLSD).
These results demonstrate that whereas the combined therapy of CS/RT magnified motor improvements following MCAo, no CS treatment effect was detected in animals following SCII.
Cortical Stimulation Movement Thresholds
CS MTs were used to set the stimulation intensity for each CS/RT animal and to provide insight into the integrity and plasticity of corticospinal pathways. A 2-way repeated-measures ANOVA on MTs during the 3 assessments (Figure 3A) detected significant effects of rehabilitation day [F(2, 56) = 130.80; P < .0001], group [F(3, 28) = 13.10; P < .0001], and the interaction [F(6, 56) = 17.50; P < .0001]. Planned mean comparisons were used to assess differences between CS/RT and RT within each lesion model. Animals given MCAo+RT were not significantly different on movement assessments relative to animals given MCAo+CS/RT on the first 2 assessments: MT1 (P = .61; FPLSD) or MT2 (P = .25; FPLSD). In contrast, the MCAo+CS/RT condition exhibited significantly lower MTs relative to the MCAo+RT condition on the third assessment: MT3 (P = .037; FPLSD). Animals given SCII+RT did not significantly differ in MTs relative to animals given SCII+CS/RT during any of the assessments: MT1 (P = .66; FPLSD), MT2 (P = .45; FPLSD), or MT3 (P = .73; FPLSD). Animals given SCII exhibited significantly greater MTs relative to animals given MCAo on all 3 assessments: MT1 (SCII+RT vs MCAo+RT, P = .00030, or SCII+CS/RT vs MCAo+CS/RT, P = .0055; FPLSD), MT2 (SCII+RT vs MCAo+RT, P < .0001, or SCII+CS/RT vs MCAo+CS/RT, P < .0001; FPLSD), or MT3 (SCII+RT vs MCAo+RT, P < .0001, or SCII+CS/RT vs MCAo+CS/RT, P < .0001; FPLSD).
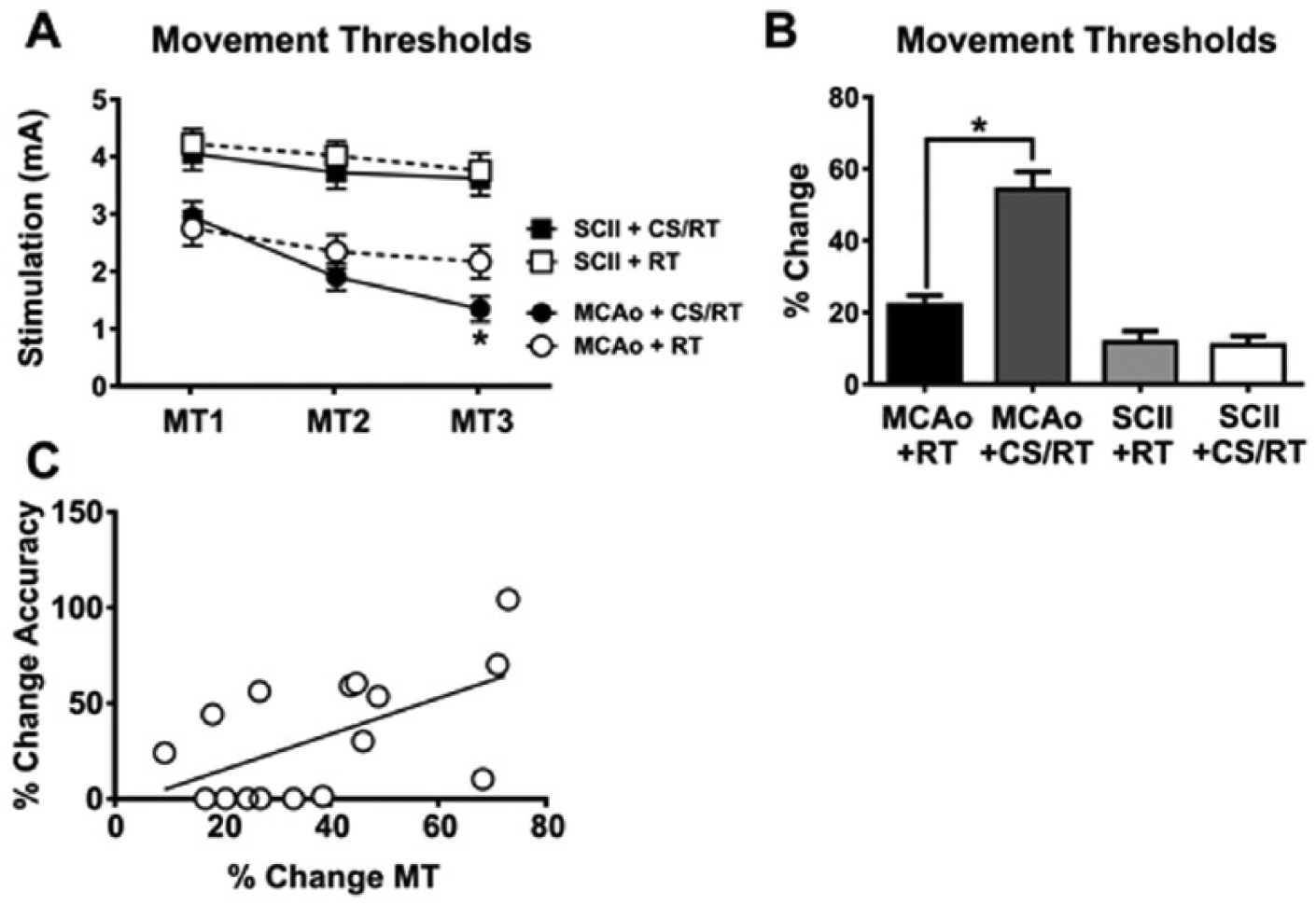
Minimum thresholds (MTs) to evoke involuntary movement with cortical stimulation (CS). CS MTs (MTs 1-3) were assessed on rehabilitation days 1, 10, and 19, respectively. A. Line graph comparing MTs for animals given either middle cerebral artery occlusion (MCAo) or subcortical capsular ischemic injury (SCII). No significant differences were detected in MTs between animals given SCII+ rehabilitative training (RT) and SCII+CS/RT. Animals given MCAo+CS/RT exhibited significantly lower MTs relative to animals given MCAo+RT on MT3 (*P < .05), whereas nonsignificant differences were detected between these groups at the other time points. B. Histogram comparing percentage change in MTs between MT1 and MT3. Animals given MCAo+CS/RT exhibited a significantly greater percentage reduction in MTs relative to animals given MCAo+RT (*P < .05), whereas this difference was not observed in SCII animals. C. For animals given MCAo, a significant positive correlation was detected between percentage change in reaching accuracy and percentage change in MTs (r2=0.33; P<.05).
The percentage change in MTs was calculated (Figure 3B) by subtracting the threshold at the first assessment from the threshold at the final assessment and then normalizing to the threshold at the first assessment [MT3 − MT1/MT1 × 100]. A significant change in MT was detected between groups [F(3, 28) = 35.14; P < .0001]. Animals given MCAo+CS/RT exhibited a significantly greater percentage reduction in MT in comparison to animals given MCAo+RT (P < .0001; FPLSD). In contrast, animals given SCII+CS/RT exhibited no difference in percentage change in MT in comparison to animals given SCII+RT (P = .87; FPLSD).
Separate correlations were used to test for relationships between the percentage change in MT and the percentage change in reaching accuracy for either MCAo+CS/RT or SCII+CS/RT animals. A significant positive correlation was detected between percentage change in MT and percentage change in reaching accuracy for animals given MCAo (r2 = 0.33; P = .021; Figure 3C). No significant correlations between change in motor threshold and reaching accuracy were found in any of the other treatment conditions.
Spared Cortical Tissue
Spared cortical tissue was estimated by calculating the percentage cortical volume from the injured hemisphere relative to the volume of the uninjured hemisphere: (Ipsilateral volume/Contralateral volume) × 100. See Figure 4A.
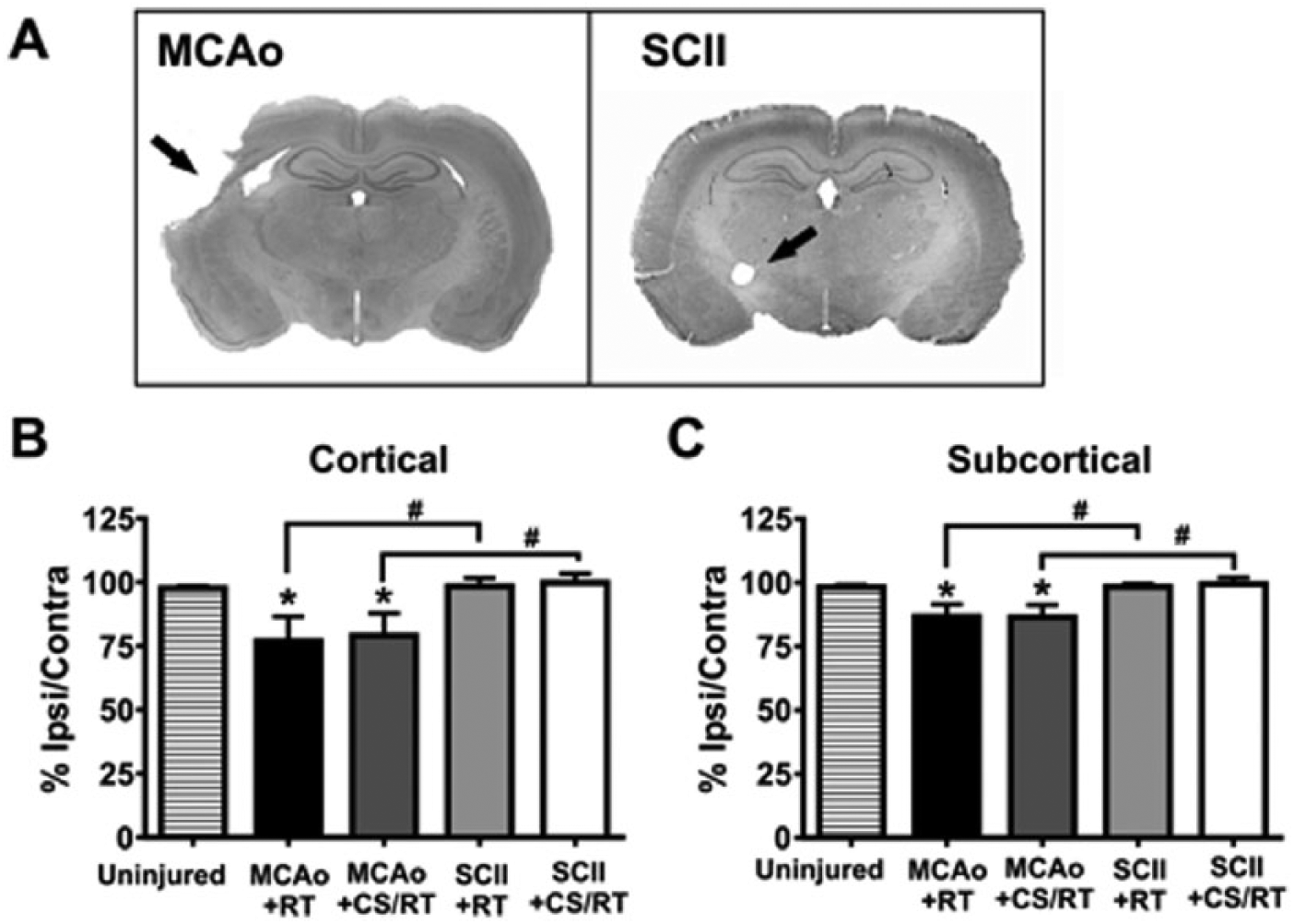
Spared tissue estimates following middle cerebral artery occlusion (MCAo) and subcortical capsular ischemic injury (SCII). A. Representative Nissl-stained coronal sections from animals given experimental MCAo or SCII. Black arrows are used to indicate regions of tissue damage. B. Histogram comparing percentage spared cortical tissue in MCAo and SCII animals. Animals given MCAo exhibited significantly less spared cortical tissue relative to uninjured control animals (*P < .05) or animals given SCII (#P < .05). C. Histogram comparing percentage spared subcortical tissue in MCAo and SCII animals. Animals given MCAo exhibited significantly less spared subcortical tissue relative to uninjured control animals (*P < .05) or animals given SCII (#P < .05).
A 1-way ANOVA revealed a significant main effect of condition (Figure 4B) on the percentage of spared cortical tissue [F(4, 35) = 3.30; P = .021]. Planned mean comparisons were used to assess differences between CS/RT and RT within each lesion model. Animals given MCAo presented with significantly less spared cortical tissue in comparison to uninjured control animals (control vs MCAo+RT, P = .025; control vs MCAo+CS/RT, P = .043; FPLSD). The 2 groups of animals given MCAo did not differ from one another in the amount of spared cortical tissue (MCAo+RT vs MCAo+CS/RT, P = .81). The 2 groups of animals given SCII did not differ from one another in the amount of spared cortical tissue (SCII+RT vs SCII+CS/RT, P = .87; FPLSD). Animals given SCII did not significantly differ in the amount of spared cortical tissue in comparison to uninjured control animals (control vs SCII+RT, P = .93; control vs SCII+CS/RT, P = .81; FPLSD). The MCAo model exhibited significantly less spared cortical tissue relative to the SCII model (MCAo+RT vs SCII+RT, P = .020, or MCAo+CS/RT vs SCII+CS/RT, P = .025).
Spared Subcortical Tissue
A 1-way ANOVA revealed a significant main effect of condition (Figure 4C) on the percentage of spared subcortical tissue [F(4, 35) = 4.046; P = .0084]. Planned mean comparisons were used to assess differences between CS/RT and RT within each lesion model. Animals given MCAo presented with significantly less spared subcortical tissue in comparison to uninjured control animals (control vs MCAo+RT, P = .019; control vs MCAo+CS/RT, P = .017; FPLSD).The 2 groups of animals given MCAo did not differ from one another in the amount of spared subcortical tissue (MCAo+RT vs MCAo+CS/RT, P = .96; FPLSD). The 2 groups of animals given SCII did not differ from one another in the amount of spared subcortical tissue (SCII+RT vs SCII+CS/RT, P = .81; FPLSD). Animals given SCII did not significantly differ in the amount of spared subcortical tissue in comparison to uninjured control animals (control vs SCII+RT, P = .98; control vs SCII+CS/RT, P = .78; FPLSD). The MCAo model exhibited significantly less spared subcortical tissue relative to the SCII model (MCAo+RT vs SCII+RT, P = .018, or MCAo+CS/RT vs SCII+CS/RT, P = .0084; FPLSD).
Discussion
The present study tested the efficacy of CS/RT to enhance motor recovery after stroke in 2 different models of cerebral ischemia that produced 2 different patterns of brain injury. The MCAo model produced damage to the lateral frontal and parietal cortex as well as the dorsolateral striatum. The SCII model produced damage that was restricted to the lateral aspect of the posterior thalamus, including the internal capsule. The results demonstrated that CS/RT differentially affected both poststroke motor performance and cortical physiology across these 2 injury models. The MCAo+CS/RT animals showed a significant increase in reaching accuracy as compared with the MCAo+RT animals. No such differences were observed between the SCII+CS/RT and SCII+RT animals. Furthermore, animals in the MCAo+CS/RT condition showed significant reductions of motor thresholds as compared with the MCAo+RT animals, and these reductions correlated with improvements in reaching accuracy. Again, no such differences were found between the SCII+RT SCII+CS/RT animals. Together, the results demonstrate that CS/RT treatment in animals with cortical infarctions augmented motor function, as evidenced by the increase in reaching accuracy, and altered cortical physiology, as evidenced by the reduction in motor thresholds.
There are several potential explanations for the differential effects of CS/RT in the experimental models. First, the 2-stroke models used here resulted in different overall lesion sizes. MCAo animals had larger cortical and subcortical lesions than both control and SCII animals. Furthermore, there was no significant difference in cortical or subcortical lesion size between SCII animals and controls. This pattern of results emerges as a result of the way in which the infarctions were created. The MCAo lesion method used here primarily resulted in damage to the mediolateral frontal and parietal cortices but also damaged the dorsal and lateral aspects of the anterior striatum. The SCII lesion method resulted in very focal lesions that were limited to the lateral aspect of the posterior thalamus, including the internal capsule. The method used to measure lesion size in this study may not have been sensitive enough to detect the small infarctions in SCII animals. However, the SCII animals did show impairments on the reaching task and had significantly higher motor thresholds than the MCAo animals, indicating that there was tissue damage. The larger infarctions in the MCAo animals were also associated with greater poststroke motor impairments when compared with the SCII animals. With daily CS/RT, the MCAo animals achieved a performance comparable to the SCII+RT and SCII+CS/RT animals. The reduced impairment levels may have masked any potential additional improvements in SCII animals resulting from the CS/RT. A second potential explanation may be related to lesion location. CS is presumed to enhance motor recovery by encouraging the recruitment/reorganization of residual cortical areas. 5 Damage to the internal capsule might hinder the effects of any such plasticity because the output of those cortical areas would be impaired. This is consistent with the observation that SCII animals had significantly higher motor thresholds than the MCAo throughout the 20 days of treatment.
In clinical studies, reductions in the magnitude or absences of motor-evoked potentials in response to cortical transcranial magnetic stimulation within days of stroke are associated with poor motor recovery.28,29 In the present study, animals with SCII required significantly greater stimulation currents to evoke movement than animals given MCAo. This suggests a greater disruption in the integrity of the descending motor tracts in SCII animals relative to the MCAo animals. The fact that MTs were present in SCII animals suggests some sparing of internal capsule fibers. This partial sparing coupled with absence of cortical damage may have supported the observed improvements in reaching accuracy.
Motor rehabilitation following cortical ischemia results in motor improvements that are accompanied by an expansion and reemergence of movement representations in the motor cortex that are specific to the training 30 and are likely mediated by synaptic plasticity.31,32 CS/RT is thought to augment these endogenous plasticity processes and is associated with greater reorganization of motor maps, increased density of synapses, 2 and increased synaptic responses 7 than RT alone.4-6 CS/RT may also have neuroprotective, angiogenic, anti-inflammatory, or growth factor–releasing properties 33 that also contribute to enhanced motor recovery. Subcortical infarctions that damage the corticospinal tract would then limit the impact of any treatment, such as CS/RT, intended to enhance plasticity within the motor cortex. This is consistent with the results of the recent Everest trial, where no differences in the overall percentage of patients that met the primary efficacy end point were found between CS/RT patients (32%) and controls (29%) 4 weeks posttreatment. 8 Yet in the 13 patients who did exhibit movements in response to the CS, 69% met the primary efficacy end point. The results highlight the importance of determining the specific pattern of brain damage resulting from stroke prior to prescribing adjuvant therapies such as CS.
Conclusions
The present findings demonstrate that the efficacy of CS for enhancing motor recovery after stroke may depend in part on the extent and location of the ischemic infarct. The results further highlight the importance of understanding the interaction between the infarct location and extent, and the use of brain stimulation to treat functional impairments after stroke.
Footnotes
Authors’ Note
No part of this work has been published.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: supported by National Institutes of Health Grant U54 NSO48126 (JAK).