
Abstract
Background. The incidence of stroke in adulthood increases with advancing age, but there is little understanding of how poststroke treatment should be tailored by age. Objective. The goal of this study was to determine if age and task specificity of rehabilitative training affect behavioral improvement and motor cortical organization after stroke. Methods. Young and aged mice were trained to proficiency on the Pasta Matrix Reaching Task prior to lesion induction in primary motor cortex with endothelin-1. After a short recovery period, mice received 9 weeks of rehabilitative training on either the previously learned task (Pasta Matrix Reaching), a different reaching task (Tray Reaching), or no training. To determine the extent of relearning, mice were tested once weekly on the Pasta Matrix Reaching Task. Mice then underwent intracortical microstimulation mapping to resolve the remaining forelimb movement representations in perilesion motor cortex. Results. Although aged mice had significantly larger lesions compared with young mice, Pasta Matrix Reaching served as effective rehabilitative training for both age-groups. Young animals also showed improvement after Tray Reaching. Behavioral improvement in young mice was associated with an expansion of the rostral forelimb area (“premotor” cortex), but we failed to see reorganization in the aged brain, despite similar behavioral improvements. Conclusions. Our results indicate that reorganization of motor cortex may be limited by either aging or greater tissue damage, but the capacity to improve motor function via task-specific rehabilitative training continues to be well maintained in aged animals.
Introduction
Stroke affects nearly 800 000 Americans each year. The majority of strokes during adulthood occur after the age of 60 years and stroke risk continues to increase with advancing age. 1 Aged animal models of stroke are important for elucidating mechanisms of effective rehabilitative therapies. Following experimental induction of stroke, aged animals show long-lasting performance deficits on sensorimotor tasks. 2 Rehabilitative tasks can be effective in promoting improvements in forelimb function in older rats and monkeys,3-5 but these improvements are somewhat limited. While young animals’ wrist and digit movement patterns during grasping normalize over time, older animals’ do not,4,5 evidence that the aged brain does not recover from stroke as well as the young brain.
In young adult squirrel monkeys, motor cortical infarcts cause a reduction in the areal extent of the functional map and a loss of skilled forelimb use. 6 Rehabilitative training induces a beneficial reorganization of the perilesion motor map concurrent with behavioral improvement. 7 However, it is unknown what effect stroke has on the aged motor cortical representations of the forelimb, which we have previously shown have already lost some complexity and learning-related plasticity in the intact brain. 8 The goal of the current study was to determine how age and task specificity of rehabilitative training affects motor cortical organization and the ability to regain a previously learned motor skill following focal ischemic motor cortical lesions in a clinically relevant animal model.
Methods
Subjects
A total of twenty-five 3- to 7-month-old and twenty-four 16- to 20-month-old male C57BL/6 mice were used. All mice were obtained from Jackson Laboratories (Bar Harbor, ME) at 1 month of age, except for a subset of aged mice obtained as retired breeders at 9 months of age (n = 17). Animals were housed in groups of 3 to 4 except for retired breeders, who were housed singly to prevent aggressive behavior. All mice received standard cage supplementation. 9 Differences in housing had no effect on behavioral improvement after stroke—main effect of group, F(1, 22) = 1.45, P = .24; group by day interaction, F(10, 220) = 1.05, P = .41—or cortical representation size (t = −0.22 to 1.71, P = .10 to .89).
To ensure proper levels of motivation during the reaching tasks, mice were maintained on scheduled feeding, receiving a daily allowance of 2.5 to 3 g of rodent chow immediately following each training session. Seven young and 10 aged mice were omitted from the study (and the above animal numbers) because of postoperative mortality. Animal use was in accordance with a protocol approved by the University of Texas at Austin Animal Care and Use Committee.
Experimental Overview
Mice received daily training on the Pasta Matrix Reaching Task (PMRT) for 8 weeks preoperatively to ensure task mastery. 8 Following task acquisition, ischemic lesions of the forelimb motor cortex were induced with endothelin-1 (ET-1), a vasoconstricting peptide. 9 Postoperative reaching success on the PMRT was assessed 4 days after surgery. Young and aged mice were randomly divided into 3 treatment conditions: (1) No rehabilitation (No-Rehab; n = 8 young, n = 8 aged), (2) Tray Reaching rehabilitation (Tray-Rehab; n = 8 young, n = 8 aged), or (3) Pasta Matrix Rehabilitation (PM-Rehab; n = 9 young, n = 8 aged). All mice were tested once weekly (probe trials) on the PMRT, following a 5-day period of rehabilitation or control procedures. Rehabilitation and probe trials were continued in this fashion for 9 weeks total. Following the final probe trial, mice underwent a terminal intracortical microstimulation (ICMS) procedure to resolve remaining forelimb movement representations in the caudal forelimb area (CFA) of the primary motor cortex and the rostral forelimb area (RFA), which is putatively homologous to premotor cortex. 10 Experimenters were blinded to the training conditions of animals during ICMS procedures.
Preoperative Training
The PMRT requires mice to reach for and break small pieces of vertically oriented, uncooked capellini pasta pieces arranged in a matrix distal and lateral to the reaching chamber aperture. In order to successfully retrieve a piece, the mouse must break the pasta by grasping and pulling forward. Detailed methods were described previously. 9
Mice were trained to reach only with the preferred limb by filling only the half of the matrix contralateral to that limb (Figure 1A). Daily training sessions consisted of up to 100 reach attempts or 15 minutes, whichever occurred first. The number of pasta pieces successfully broken was recorded. Preoperative training was conducted 6 days per week, with 1 day off, for a total of 10 weeks. Mice received no preoperative training on the Tray Reaching Task (Figure 1B).
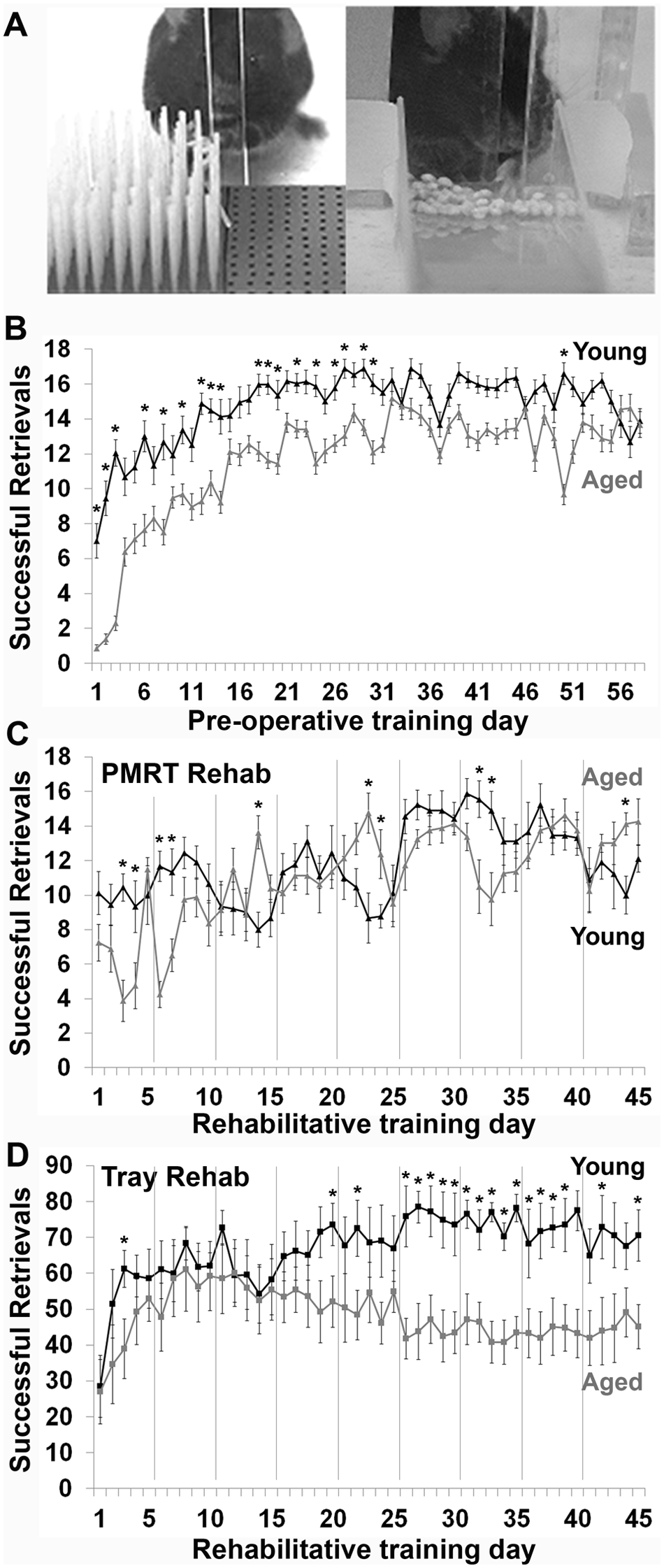
Aged mice are able to learn and perform skilled motor tasks. (A) Examples of mice performing the two rehabilitative training tasks: the Pasta Matrix Reaching Task (PMRT, left) and the Tray Reaching Task (right). (B) Results of postoperative PMRT training trials in young and aged mice. Successful retrievals indicate the number of pasta pieces broken and eaten per training session. (C) Results of PMRT rehabilitative training trials in young and aged mice. (D) Results of Tray Reaching rehabilitative training trials in young and aged mice. Successful retrievals indicate the number of millet seeds grasped and eaten per training session. Gray vertical lines in C, D indicate 2-day breaks in rehabilitative training to allow for probe trials on the PMRT and days off of training. *p < .05, young versus aged. Data are means ± standard error.
Endothelin-1 Lesions
Endothelin-1-induced lesion surgeries were performed as previously described. 9 Under anesthesia (100 mg/kg ketamine and 10 mg/kg xylazine, intraperitoneal), a small burr hole was drilled through the skull over the forelimb representation of primary motor cortex, at 1.5 mm lateral to midline and +0.3 mm anterior to Bregma. 11 The dura was punctured and a calibrated pipette with a tip diameter of ~50 µm was lowered into the cortex to a depth of 800 µm. For young mice, 4 µL of ET-1 (American Peptide; 320 pmol in saline) was infused into the cortex. Aged mice received a smaller volume of ET-1 (3 µL) based on a pilot study that indicated that the larger volume (and dose) increased mortality rates. The burrhole was then filled with gelfoam and covered with ultraviolet curing dental cement prior to suturing. Each animal was allowed to fully awaken in a heated chamber, and received buprenorphine (3 mL/kg, subcutaneous) before it was returned to the home cage. Postlesion mortality rates were not significantly different between young and aged mice (21% vs 29%, respectively); χ2(1, n = 66) = 0.49, P = .52.
Rehabilitative Training
Rehabilitative training began 5 days after lesion induction surgery, and was then conducted 5 days per week, followed by 1 probe trial day and 1 day off, for a total of 9 weeks. Probe trials of the PMRT were used to access deficits in, and relearning of, this previously acquired task. Postoperative PMRT sessions, including probe trials, were conducted exactly as preoperative sessions. Control procedures for mice in the No-Rehab condition consisted of eating pieces of pasta too small to handle from the floor of a reaching chamber for a maximum of 15 minutes per day.
Mice in the Tray-Rehab condition performed the Tray Reaching Task, in which they were allowed to reach for 100 millet seeds placed in an inclined glass tray (2.5 × 2.5 × 7.5 cm, with a 6-mm tall front lip; Figure 1A). Rehabilitation was restricted to the contralesional limb by aligning the edge of the tray with the lateral edge of the reaching slit, on the contralateral side of the reaching chamber. The close proximity of the reaching slit to the chamber wall (1.8 cm) forced a reaching angle in which mice were only able to successfully obtain seeds with the contralateral paw.
Intracortical Microstimulation
Mice underwent a terminal ICMS procedure 1 to 2 days following the final PMRT probe session (9 weeks postinfarct). Motor cortical representation areas were defined by the movements generated at the lowest stimulation thresholds, the approach traditionally used to characterize motor cortical maps in primates and rats6,7,12 and which we have previously refined for mice.8,11 Aged mice were more sensitive to the effects of anesthesia, and received a smaller anesthetic dose for the procedure (young mice, 150 mg/kg ketamine and 10 mg/kg xylazine; aged mice, 100 mg/kg ketamine and 4 mg/kg xylazine, intraperitoneal).
Intracortical microstimulation was conducted as previously described.8,11 A large craniotomy was made over the motor cortex. Intracortical penetrations with a glass microelectrode (15-µm tip) were made at 790- to 800-µm depths in 250-µm increments, bordering in all forelimb responsive sites with nonforelimb or nonresponsive sites. Penetrations were made throughout the typical extent of the mouse forelimb area 11 to ensure that all forelimb responsive sites were detected.
At each site, a 40-ms train of thirteen 200-µs monophasic cathodal pulses was delivered at 350 Hz from an electrically isolated, constant current stimulator (BAK Electronics) at a rate of 1 Hz. Stimulation was increased until a visible movement was evoked on the contralateral side of the body, up to a maximum of 100 µA. We used a higher maximal current than our previous studies in non-stroke affectedmice (60 µA)8,11 to compensate for potential dampening of cortical responses near the infarct. 13 The movement evoked at the lowest current (stimulation threshold) was recorded for each site. If no movement was seen at or less than 100 µA, the site was considered nonresponsive.
Histology and Infarct Analysis
Immediately following the ICMS procedure, mice were euthanized with an overdose of sodium pentobarbital (175 mg/kg, intraperitoneal) and perfused intracardially with 0.1 M phosphate buffer and 4% paraformaldehyde. Brains were postfixed in paraformaldehyde and sliced on a vibratome into 50-µm thick sections. Every sixth section was mounted onto gelatin-coated slides and Nissl stained with toluidine blue.
Coronal sections were viewed at a magnification of 50×. The cortical boundaries of 9 coronal sections per animal from approximately 2.0 mm anterior to 1.2 mm posterior to Bregma were traced using Neurolucida software. Cavalieri’s method was used to calculate total remaining cortical volume as the product of summed section areas and distance between sections. 14 Lesion volume was indirectly estimated as the difference between the volumes of the damaged and intact cortices. Remaining peri-infarct forelimb movement representation areas were estimated by comparing total map areas to previously published baseline cortical area values. 8
Statistics
SPSS software was used for all statistical analyses. A priori power analysis indicated that a sample size of 6 mice per group was necessary to detect significant effects in skilled reaching behavior and ICMS-evoked maps at 80% power with an α level of .05. Two-tailed t tests were used to compare cortical volumes and interhemispheric volume differences between young and aged mice. A repeated-measures analysis of variance (ANOVA) was used to compare preoperative learning curves of young and aged mice on the PMRT. Repeated-measures ANOVAs were conducted on postoperative behavioral data to determine (1) if either type of rehabilitative training improved reaching success on probe trials, relative to controls (PM-Rehab vs No-Rehab and Tray-Rehab vs No-Rehab) and (2) if one task was more effective in promoting improvement over the other (PM-Rehab vs Tray-Rehab), using separate analyses for young and aged animals and Bonferroni-corrected post hoc comparisons. Young and aged performance on initial postoperative and postrehabilitative training probe trials was compared with 2-tailed t tests. To directly assess the effect of age on behavioral improvement within each treatment condition, we used repeated-measures ANOVAs and Bonferroni-corrected post-hoc tests to compare the change from initial postoperative performance of young and aged mice.
Two-tailed t tests were conducted to determine whether (1) the remaining representation areas or ratio of proximal to distal forelimb representations were affected by either type of rehabilitative training (PM-Rehab vs No-Rehab and Tray-Rehab vs No-Rehab) and (2) if the 2 tasks resulted in differing cortical organization (PM-Rehab vs Tray-Rehab). Data from the CFA and RFA, as well as young and old mice, were analyzed separately.
Results
Preoperative Learning on the PMRT
Preoperative training was conducted 6 days per week for a total of 10 weeks, in order to ensure that both young and aged mice reached proficiency on the task prior to lesion induction (Figure 1B). Young mice initially performed better on the task, and started at a higher success level on the first day of training, t(47) = 5.99, P < .001. A significant main effect of age, F(1, 47) = 20.61, P < .001, and a day × age interaction, F(57, 2679) = 8.45, P < .001, suggest that the pattern of learning differed between young and aged mice. This was especially evident in the first month of training. However, over the course of preoperative training, aged mice were able to catch up to young mice. By the last 3 days of training, which were averaged to compute a baseline score for comparison with postoperative rehabilitative training, there was no significant difference between the age-groups, t(47) = 0.46, P = .44. These results agree with previously published work from our laboratory, showing that aged mice are able to learn a new skilled reaching task to a similar extent as young mice, albeit more slowly. 8
Infarct Analysis
Endothelin-1 infusion resulted in significantly more tissue damage in aged mice compared with young mice (Figure 2), despite a smaller ET-1 dose in aged mice (3 vs 4 µL in young mice). There was no difference in the volume of the contralesional hemisphere between young and aged mice, t(47) = 0.29, P = .77. However, the ipsilesional hemisphere was significantly smaller, t(47) = −3.92, P < .001, and the interhemispheric volume differences were greater, t(47) = −2.87, P = .006, for aged mice than for young mice (Figure 2A). There was no significant difference in infarct size between the training conditions in young—F(2, 24) = 0.69, P = .51—or aged animals—F(2, 23) = 0.74, P = .49). Representative coronal sections from young and aged animals are shown in Figure 2B.
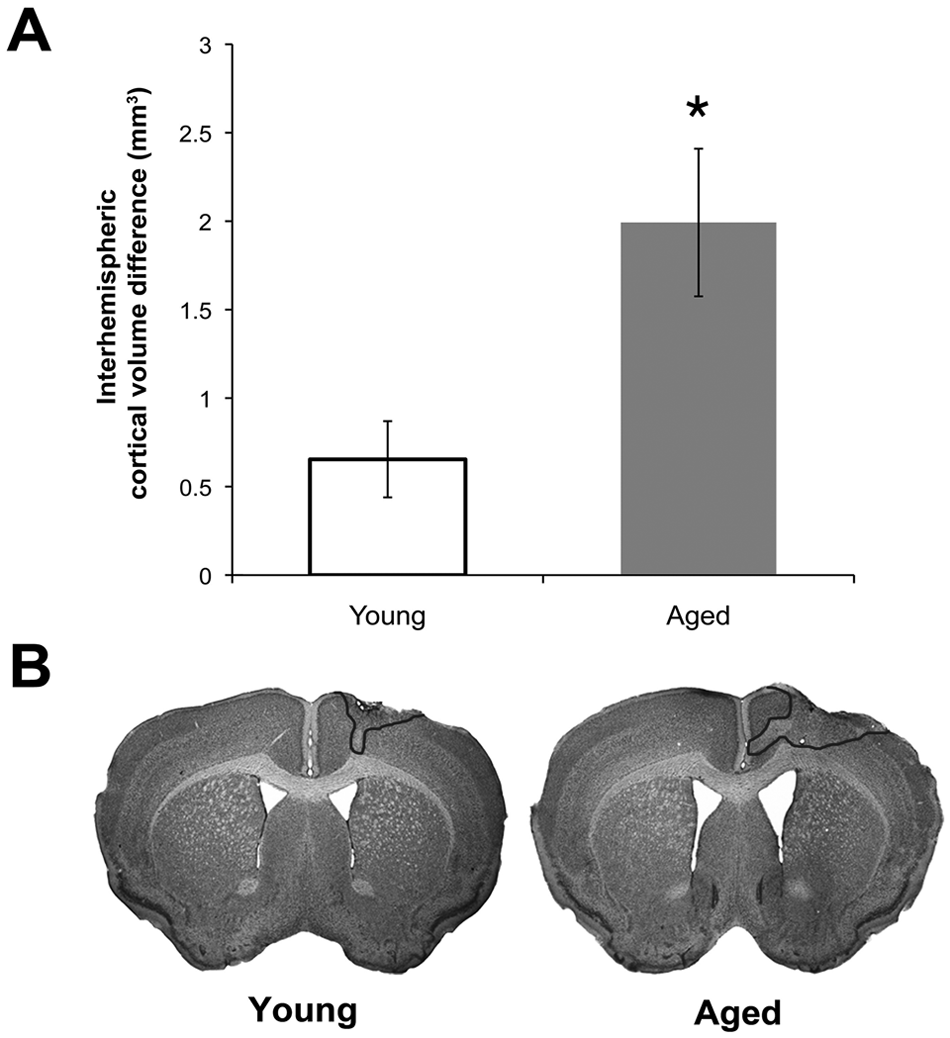
Endothelin-1-induced infarcts were larger in the aged mouse brain. (A) Interhemispheric volume differences were used to estimate infarct size. (B) Representative Nissl-stained coronal sections from young (left) and aged (right) animals. The infarct is outlined in the right hemisphere in both cases. *P < .05, young versus aged. Data are means ± standard error.
Rehabilitative Training
Young and aged mice were equally as successful during rehabilitative training sessions on the PMRT, in which they “relearned” the previously acquired reaching task (Figure 1C). There was a significant effect of rehabilitative training day, F(44, 660) = 9.14, P < .001, but no significant effect of age, F(1, 15) = 0.58, P = .46. A significant day × age interaction, F(44, 660) = 4.57, P < .001, suggested that the pattern of relearning differed between young and aged mice. Although aged mice were initially more impaired on the PMRT after stroke, they quickly relearned the task and performed at a similar success level to young mice.
When mice received rehabilitative training on the untrained reaching task, Tray Reaching, we found a significant effect of rehabilitative training day, F(44, 616) = 3.65, P < .001, and a significant day × age interaction, F(44, 616) = 3.28, P < .001. There was a strong trend toward an effect of age, but this did not reach statistical significance, F(1, 14) = 4.53, P = .052. The 2 groups had similar performance for the first month after stroke (Figure 1D). Only after this time did young mice begin to surpass aged mice in successful performance on this task. Thus, while aged mice were able to successfully learn this new task, they were ultimately not as skilled on this task as young mice.
Pasta Matrix Reaching Task Probe Trials: Young Mice
Initially, the 3 groups of young mice had similar postoperative deficits (Figure 3A). Rehabilitative training on either reaching task began to improve success levels in the first training week. The greatest postoperative performance levels on the PMRT were achieved in mice that received rehabilitative training on this task. When the results of postoperative weekly probe trials were compared, there were significant overall improvements in reaching performance as a result of training on both the PMRT—F(1, 15) = 4.13, P = .04, PM- versus No-Rehab—and Tray Reaching Task—F(1, 14) = 4.70, P = .048, Tray- versus No-Rehab—compared with no-training controls. Neither analysis indicated significant day × group interactions (Ps = .16 and .31, respectively). There was no significant group effect, F(1, 15) = 1.01, P = .33, PM- versus Tray-Rehab, or day × group interaction, F(1, 150) = 1.40, P = .21, when the 2 types of training were directly compared. These results indicate that both reaching tasks were effective as rehabilitative training in young mice.
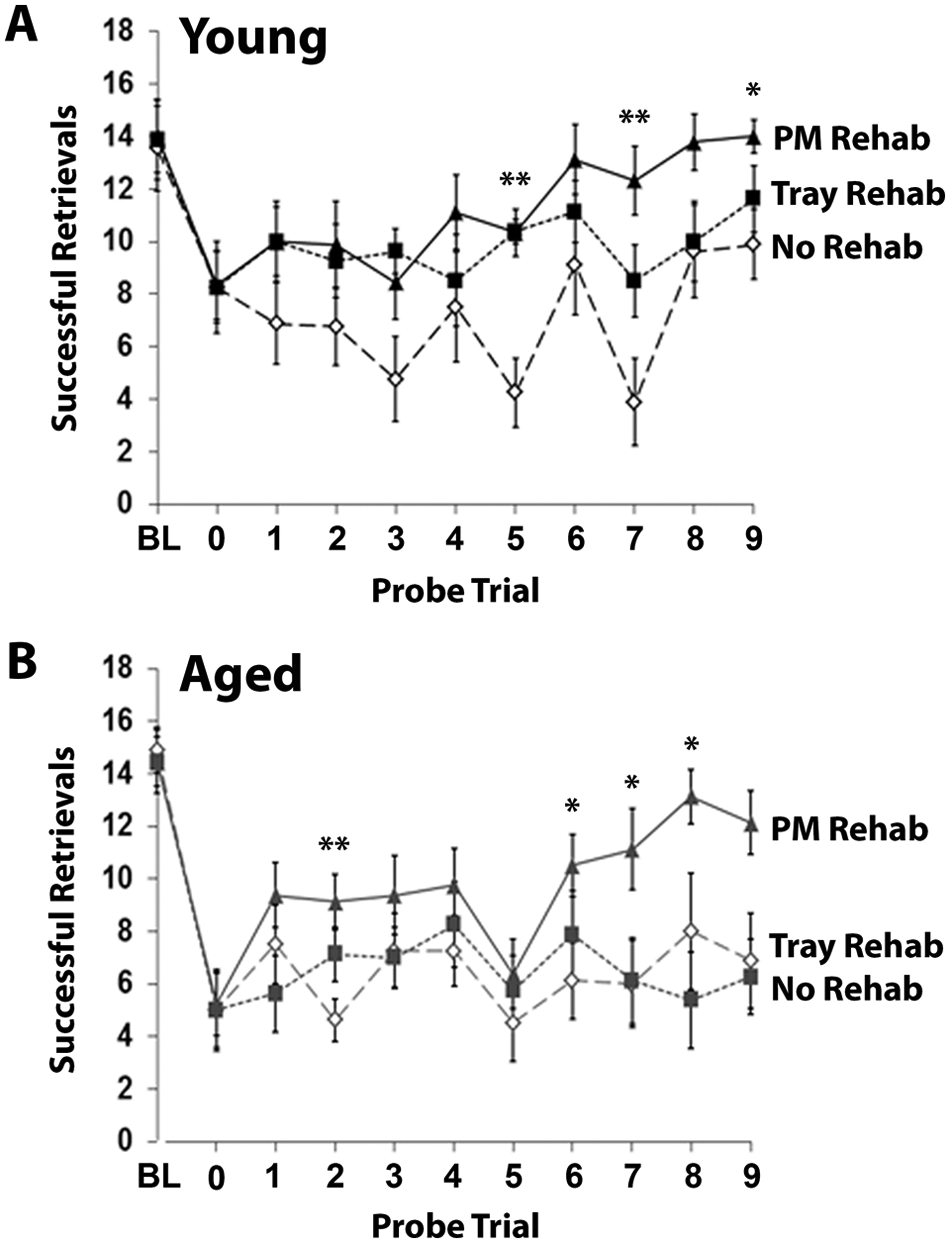
Relearning a complex motor skill after ischemia is dependent on the type of rehabilitative training provided. (A) Results of Pasta Matrix Reaching Task (PMRT) probe trials throughout the rehabilitative training period in young (A) and aged (B) mice. Successful retrievals indicate the number of pasta pieces broken and eaten per training session. BL = baseline. Data are means ± standard error.
Pasta Matrix Reaching Task Probe Trials: Aged Mice
Initially, all groups of aged mice had similar postoperative deficits (Figure 3B). Like young mice, training on the PMRT in aged mice improved reaching success compared no training. While PMRT training significantly improved overall performance compared with no-training controls—F(1, 14) = 4.63, P = .049, PM- versus No-Rehab—Tray Reaching did not, F(1, 14) < 0.001, P = .99, Tray- versus No-Rehab. When the 2 types of training were compared, there was a significant group effect—F(1, 14) = 5.27, P = .04, PM- versus Tray-Rehab. None of the analyses revealed significant group × day interactions (P values >.05). These results indicate that only the PMRT was effective as rehabilitative training in aged mice.
Comparison of Young and Aged Mice
Preoperative performance on the PMRT was similar in young and aged mice, t(47) = −0.78, P = .44. Aged mice, which had larger lesions than young mice, were more impaired in the first postoperative probe session—t(47) = 2.85, P = .007—and at the end of the 9 weeks of rehabilitative training compared with young mice—t(47) = 2.91, P = .005. To assess the effect of age on behavioral improvement in each rehabilitative training group, we directly compared young and aged mice for their change from baseline performance (Figure 4). This revealed that there were no effects of age on reaching performance in the No-Rehab (Figure 4A)—F(1, 14) = 2.14, P = .17—and PM-Rehab conditions (Figure 4C)—F(1, 15) = 2.12, P = .17. However, there was a significant age difference in the effectiveness of Tray-Rehab (Figure 4B) F(1, 14) = 7.64, P = .02. There were no day × age interactions within any of the 3 rehabilitative training condition (P values = .16, .71, and .29, respectively). These results indicate that even though aged mice were initially more impaired and had more cortical damage than young mice, rehabilitative training on the PMRT was similarly effective across ages in promoting behavioral improvement. However, rehabilitative training on the Tray Reaching Task failed to induce sufficient behavioral improvement in aged mice, despite being relatively effective in younger mice.
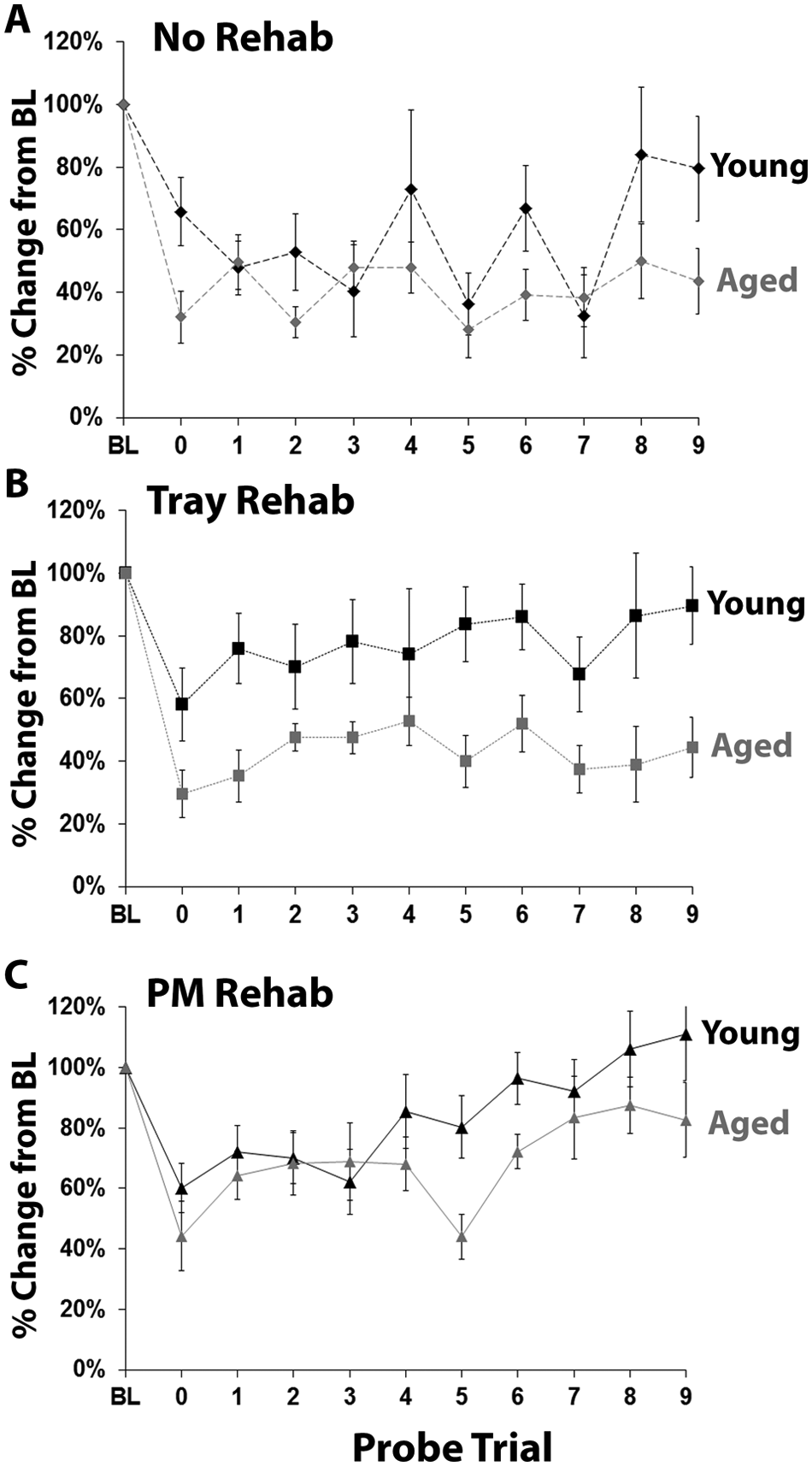
Rehabilitative training on the Pasta Matrix Reaching Task (PMRT), but not the Tray Reaching Task, was similarly effective across ages in promoting behavioral improvement. (A-C) Results of PMRT probe trials comparing young and aged animals that received (A) no rehabilitation, (B) Tray Reaching rehabilitation, or (C) PMRT rehabilitation. There was a significant effect of age only in the Tray Rehab condition.
Intracortical Microstimulation
All infarcts were within the boundaries of the CFA in the motor cortex. Notably, although aged mice had larger infarcts than young mice, both age groups retained a large amount of cortical map area (Table 1). As previously found, 11 only a subset of mice had a discernible RFA, located just anterior to the CFA. The number of aged animals with an evident RFA (n = 10 out of 24) was not different from that of young animals (n = 12 out of 25); χ2(49) = 0.20, P = .90. However, all animals, including those without a discernible RFA (ie, RFA area = 0 mm2), were included in the analyses of ICMS data, except for the remaining cortical representations areas presented in Table 1.
Estimated Remaining Forelimb Motor Cortical Representation Areas (mm2) Relative to Intact Brains. a
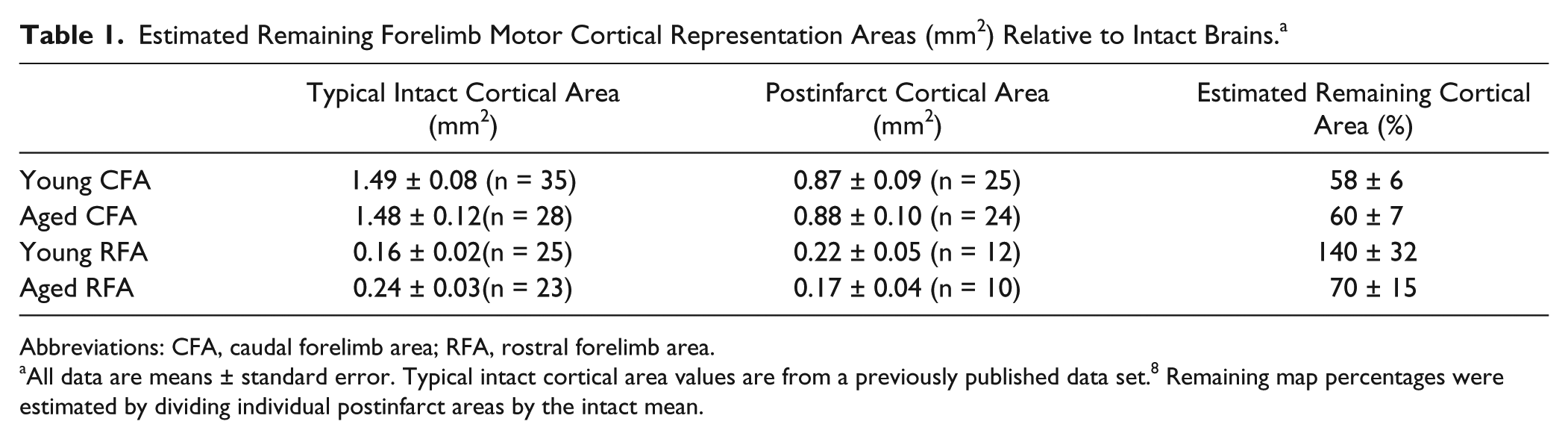
Abbreviations: CFA, caudal forelimb area; RFA, rostral forelimb area.
All data are means ± standard error. Typical intact cortical area values are from a previously published data set. 8 Remaining map percentages were estimated by dividing individual postinfarct areas by the intact mean.
Neither rehabilitative training strategy had a significant effect on the overall size of the remaining peri-infarct CFA in young mice (Figure 5A)—t(14) = 1.40, P = .18, PM- versus No-Rehab; t(9) = −0.06, P = .95, Tray- versus No-Rehab. However, young mice that received rehabilitative training on the PMRT had a significantly larger RFA compared with mice that received no training (Figure 5B) t(9) = −2.34, P = .04. Tray reaching seemed to have an intermediate effect on RFA size, as this group was not significantly different from either the PM-Rehab—t(11) = −1.85, P = .09—or No-Rehab—t(10) = 0.67, P = .52—groups. There were no group differences in the areas of digit, wrist, elbow, or shoulder representations within either the CFA (t-statistics = −1.71 to 1.09, P values = 0.11 to 0.88) or RFA (t-statistics = −1.75 to 1.00, P values = 0.11 to 0.88), nor in the sum or ratio of proximal and distal movement representations (CFA, t-statistics = −1.13 to 1.56, P values = 0.13 to 0.71; RFA, t-statistics = −2.10 to 1.44, P values = 0.07 to 0.88).
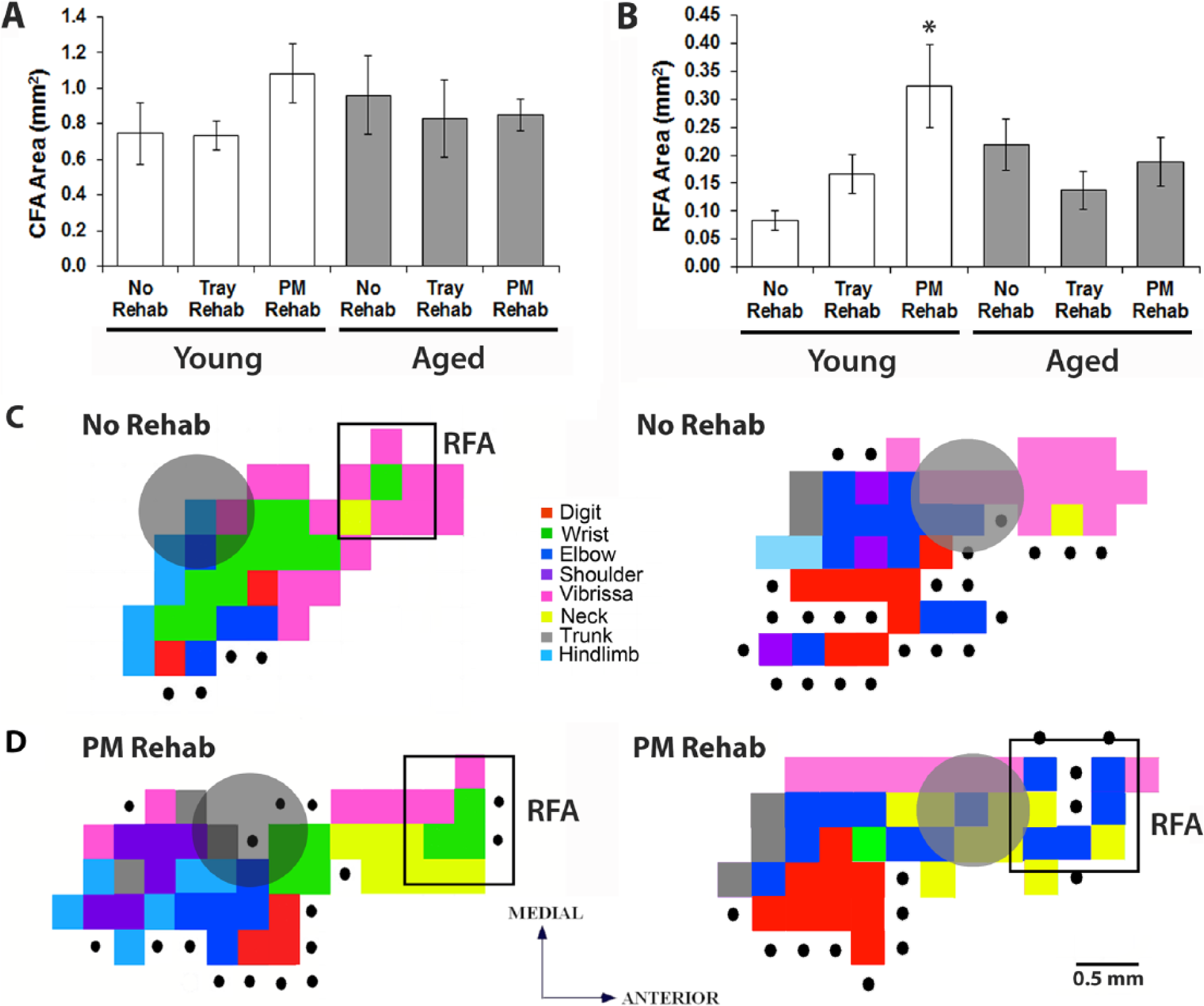
Rehabilitative training on the Pasta Matrix Reaching Task (PMRT) resulted in a larger rostral forelimb area (RFA) in young mice. (A) Caudal forelimb areas (CFAs) across age and training conditions. (B) RFAs across age and training conditions. (C) Representative intracortical microstimulation (ICMS)-evoked forelimb representations from young mice that received no rehabilitative training (C) and rehabilitative training on the PMRT (D). Grey circles indicate the infarct location. *P < .05 Pasta Matrix versus No Rehab. Data are means ± standard error.
Despite remarkable behavioral improvements following rehabilitative training, we failed to see reorganization of aged forelimb motor representations (Figure 5A and B). There were no rehabilitative training effects on the overall area of the CFA—t(9) = −0.48, P = .65, PM- versus No-Rehab; t(13) = −0.42, P = .68, Tray versus No-Rehab—or RFA—t(8) = −1.43, P = .19, PM- versus No-Rehab; t(11) = 0.41, P = .69, Tray- versus No-Rehab. There were no group differences in digit, wrist, elbow, or shoulder representations within either the CFA (t-statistics = −1.03 to 2.05, P values = 0.07 to 0.98) or RFA (t-statistics = −1.44 to 1.45, P values = 0.18 to 1.00) nor in the sum or ratio of proximal and distal representations (CFA, t-statistics = −0.81 to 1.65, P values = 0.13 to 0.85; RFA, t-statistics = −1.43 to 1.43, P values = 0.19 to 1.00). The direct comparison of young and aged motor maps is complicated by baseline differences in the size of RFA (Table 1). 8 Thus, even though there was a significant enlargement of RFA in young mice following rehabilitative training on the PMRT compared with young No-Rehab controls, there was no difference in CFA or RFA size between young and aged mice when comparing within treatment groups (t values = 0.43 to 1.12, p values = 0.27 to 0.67). Our conclusion that reorganization is age dependent is based on the evidence that rehabilitation resulted in map reorganization in young, but not aged, mice compared with age-matched controls.
Discussion
Following focal ischemic lesions, aged mice exhibit deficits similar to those seen in human stroke survivors, including impairments in coordinated and dexterous use of the contralesional upper extremity. Task-specific rehabilitation has been shown to improve motor function (ie, dexterity, range of motion) of the affected arm and hand15-18 and prevent compensation with the less-impaired side of the body.19,20 Our results suggest that task-specific rehabilitative training may be an especially effective way to induce behavioral improvement of the upper extremities in older stroke survivors and drive beneficial plasticity in premotor areas of the younger brain.
Relearning skilled movements through rehabilitative training results in the maintenance and reorganization of surviving motor representations in young rats and primates.7,21,22 In squirrel monkeys, lesions of the MI hand area result in enlargement of the hand representation within the ventral premotor cortex (PMv). 23 Following lesions of both the MI and PMv, there is an enlargement of the hand representation in the supplementary motor area (SMA). 24 Recent evidence implicates that similar plasticity is occurring in areas of the rodent brain that are analogous to premotor or supplementary motor areas of the primate brain. Gharbawie et al 25 showed that rats with complete lesions of the motor area, inclusive of CFA and RFA, were more impaired on a skilled reaching task than animals that only had damage to the CFA, leaving RFA intact. Zeiler et al 26 found a reduction in inhibitory markers in the medial agranular cortex (AGm) following lesions of the CFA and postoperative retraining on a skilled reaching task. The deficits were reinstated by damaging the AGm after relearning had occurred, suggesting that other areas of the mouse brain, such as the RFA, also play a role in stroke recovery. Our finding that ICMS-evoked motor representations in the RFA expanded and reorganized in young mice following successful rehabilitative training further supports the idea that plasticity in nonprimary motor areas is important for poststroke behavioral improvement with rehabilitative training, at least in the young brain.
Although aged animals regained reaching ability following rehabilitative training on the PMRT, we failed to find any changes in ICMS-evoked motor maps. This could be explained by the larger lesions seen in aged brains resulting in greater damage to the areas that would normally compensate for lost functions. Interestingly, although aged mice had larger infarcts than young mice, they retained a similar amount of CFA representation area compared to typical intact representations areas. 11 This either suggests that much of the CFA was spared after ischemia or that the forelimb movement representations extended into peri-infarct tissue. If this second point is true, then cortical plasticity may not be as constrained in the aged brain as the lack of enlargement of RFA representations would imply. Alternatively, because reorganization of movement representations occurs late in the process of skill learning, 27 it is feasible that plasticity in the aged RFA has not yet happened, but could occur following a longer duration of rehabilitative training. We have previously reported that, in the absence of brain damage, there is a dissociation between behavioral performance and cortical reorganization in aged animals. 8 It is not unheard of for a cortical area to undergo reorganization after brain injury and rehabilitative training, but not after motor skill learning in the absence of brain damage. In rats, reorganization of the RFA, which was not seen due to motor learning alone, 12 could be resolved using long-duration ICMS after ischemic lesions of the CFA. 28 However, this does not seem to be the case in the aged brain. It is likely that motor learning or relearning in the aged brain is simply not reflected in the reorganization of movement representations as detected using the ICMS methods of this study. This hardly rules out the possibility of an age-related recruitment of other brain areas for acquisition or relearning of motor skills. In the human motor system, the aged brain is able to compensate for the slowed processing capabilities of primary motor areas by recruiting additional areas of the brain, including contralateral motor areas and ipsilateral sensory and cognitive areas, during performance of a motor task. 29 Future studies from our laboratory will investigate learning- and stroke-induced plasticity occurring in other cortical and subcortical areas (such as the somatosensory cortex and thalamus) that are strongly interconnected to the motor cortex. Thus, although we did not see clear evidence of plasticity of the aged premotor cortex in the current study, it does not rule out the possibility that the aged brain retains plasticity in response to ischemia and rehabilitative training.
Overall, the results of this study show that while the aged brain may suffer more damage in response to ischemia, aged animals are able to regain a remarkable amount of motor function if provided with task-specific rehabilitative training. In contrast to young animals, in which rehabilitative training on a novel task, the Tray Reaching Task, produced some behavioral improvement, aged mice benefited less from postoperative training on the Tray Reaching Task. This suggests that the ability to master new tasks after stroke or to generalize between motor tasks is lost or impaired during aging. Thus, in order for rehabilitative therapy to be effective in older stroke survivors, clinicians must take care to target therapy so that it is specific to the task being relearned.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute of Neurological Disorders and Stroke (NS056839 to TAJ) and the National Institute on Aging (F31AG034032 to KAT).