
Abstract
Background. Functional neuroimaging is increasingly used in rehabilitation research to map the neural mechanisms subserving training targets. These data can inform intervention design and improve evaluation of treatment outcomes. Reliable neural markers may provide standard metrics of treatment impact and allow consideration of behavioral outcomes in the context of functional brain changes. Objective. To identify common patterns of functional brain changes associated with training across a diverse range of intervention protocols. Reliable brain changes could inform development of candidate neural markers to guide intervention research. Methods. Taking a quantitative meta-analytic approach, we review the functional neuroimaging studies of cognitive and motor skills training interventions in healthy young adults (N = 38). Results. Reliable decreases in functional brain activity from pretraining to posttraining were observed in brain regions commonly associated with cognitive control processes, including lateral prefrontal, left anterior inferior parietal lobule, and dorsal anterior cingulate cortex. Training-related increases were observed in the medial prefrontal cortex and posterior cingulate and angular gyrus, core regions of the default network. Activity within the subcortical striatum also showed reliable increases pretraining to posttraining. Conclusions. These data suggest that altered engagement of large-scale, spatially distributed cortical brain networks and subcortical striatal brain regions may serve as candidate neural markers of training interventions. The development of reliable metrics based on activity and functional connectivity among large-scale brain networks may prove fruitful in identifying interactions between domain-general and -specific changes in brain activity that affect behavioral outcomes.
Introduction
Functional brain data are increasingly used to inform rehabilitation research, with early evidence suggesting a role for neuroimaging in the design and evaluation of more targeted intervention protocols. 1 Studies of functional brain changes associated with training intervention vary widely with respect to training design (eg, repeated practice, guided skill learning, problem-solving techniques) and targeted behavior (eg, motor skills, cognition, and sensory-perceptual abilities). As a result, the neural basis of training-related performance gains is typically characterized as patterns of functional changes in localized brain regions known to be engaged by the training target. 2 Variability across studies and training methods pose a significant challenge for identifying broadly applicable and reliable neural markers of training. The purpose of the current meta-analysis is to investigate whether common patterns of functional brain changes may be associated with cognitive and motor skills training across a diverse range of intervention protocols. If reliable patterns emerge, this would provide preliminary evidence for overlapping neural mechanisms subserving training-related performance gains and inform the development of reliable metrics for measuring functional brain changes. Such domain-general markers would complement intervention- or domain-specific markers to better elucidate large-scale and localized functional brain changes associated with training across a wide array of training and rehabilitation interventions.
Although several qualitative reviews of studies examining functional brain changes associated with training have been published,2-8 we are aware of only 1 review 9 that has used quantitative meta-analytic methods. Previous reviews have revealed both increases and decreases in activity across multiple, spatially distributed brain regions; however, there is mounting evidence for more global or domain-general brain changes. Kelly and Garavan 2 reported that patterns of functional brain change differed for motor versus cognitive training interventions. They observed activation decreases in executive control and attention-related areas (prefrontal cortex [PFC], anterior cingulate cortex [ACC], and posterior parietal cortex) in the initial stages of motor-related training, whereas later stages of training (ie, well-learned motor behavior) were associated with increases or “expansions” in motor/sensory areas (eg, primary motor cortex, the supplementary motor area [SMA], and the basal ganglia). In contrast, cognitive training was associated with a general pattern of activation decreases across a more distributed network of brain regions. Indeed, posttraining decreases in activation have been consistently emphasized as a neural marker for well-automated or highly trained performance.2,10 The “scaffolding framework” put forth by Peterson et al 11 proposes that early learning is supported by a set of regions that come online to support novel task demands. After extensive practice, these scaffolding regions disengage, whereas a set of different regions are recruited to store representations of the trained skill. 11 Posttraining decreases have been reliably observed in prefrontal and other attention-related regions across both motor and cognitive domains, suggesting that these regions provide important control resources to support novel and effortful performance (see Chein and Schneider 9 ).
More recent qualitative reviews3,4,6 suggest that there may be less of a divergence between patterns of functional brain change across motor and cognitive training interventions. Within the motor domain, Doyon et al 4 posited that 2 pathways—a corticostriatal and corticocerebellar pathway—contribute to the early acquisition of motor skills. Together, these pathways engage the striatum, cerebellum, and motor cortical regions (eg, primary motor cortex, SMA, pre-SMA) as well as the PFC, parietal cortex, and the hippocampus during initial learning. As task performance reaches asymptotic levels, activity in the corticocerebellum pathway—typically associated with intentional motor skill acquisition—decreases, whereas activity in the striatum (caudate/putamen) remains steady, suggesting that the striatum may be critical for long-term retention of well-learned motor sequences. Similarly, Dahlin et al 3 reported that working memory training led to decreases in cortical areas typically related to working memory and attention (eg, frontal and parietal regions) and increases in the subcortical striatum. These data suggest that for both motor and cognitive training, there is lesser involvement of control-related cortical areas (eg, PFC) and an increase in the recruitment of subcortical striatum as training progresses.
Quantitative evidence for more domain-general or global patterns of neural change associated with training was also reported by Chein and Schneider, 9 who developed in-house quantitative methods to analyze brain imaging findings from 29 training studies. The authors contrasted activation maps showing training-related increases and decreases across both motor and cognitive domains. Training was associated with decreases across a broad network of regions implicated in executive and attentional control, whereas increases were limited primarily to posterior brain regions. Unlike these other reviews, training-related changes in the subcortical striatum were not reported. However, a majority of studies reviewed by Chen and Schneider contrasted “trained” versus “untrained” conditions during scanning; thus, training-related differences may have been at least partially obscured by differences in task characteristics.
Recently, evidence for training-related changes in large-scale functional brain networks has begun to emerge, providing further support for efforts to derive global markers of training efficacy. The default network is an interconnected set of brain regions, including medial PFC (mPFC), posterior cingulate cortex, superior and inferior frontal gyri, medial and lateral temporal lobes, and the posterior extent of the inferior parietal lobule. 12 These regions are typically suppressed during externally oriented, attention-demanding tasks. 12 Activation increases in the default network have been reported following problem-solving training in young adults 13 and visuospatial attention training following traumatic brain injury. 14 Changes following training have also been reported in frontoparietal control and dorsal attention networks, which are engaged during tasks requiring externally focused attention 15 and rapid adaptation of thoughts and behaviors to changing internal states and external environments.16,17 Regions of the frontoparietal control network include lateral PFC, precuneus, the anterior extent of the inferior parietal lobule, dorsal ACC, and the anterior insula.16-18 The dorsal attention encompasses the dorsolateral PFC, frontal eye fields, inferior precentral sulcus, superior occipital gyrus, middle temporal motion complex, and superior parietal lobule.15,19 Reduced activity in the frontoparietal control and dorsal attention networks has been observed following cognitive training.14,20,21 Dynamic changes in the connectivity profiles of large-scale functional brain networks was also shown during a motor learning task. 22 Moreover, these brain changes predicted future learning ability, highlighting the potential of functional brain data to inform intervention research.
Here we used quantitative meta-analytic methods, the activation likelihood estimation (ALE 23 ) technique to review all studies investigating functional brain changes associated with cognitive and motor skills training using a longitudinal, within-subject experimental design. Our aim was to identify common patterns of functional brain changes associated with training across a diverse range of intervention protocols within these broader training domains. If reliable patterns emerge, these could provide the basis for establishing global markers of functional brain changes that could be used, in combination with more localized, domain-specific training markers, to inform intervention design and provide brain-based outcome measures for evaluating treatment efficacy.
Methods
Study Selection
A systematic literature search was conducted to identify neuroimaging studies of cognitive and motor skills training for inclusion in the meta-analysis. Peer-reviewed articles, published in English up until June 2012, were selected from the search results of 2 separate databases: MEDLINE and PsycINFO. The literature search was conducted using the following search words: (1) keywords: “train* † ”<OR> “learn*” <OR> “rehab*” <OR> “practic*” >; AND (2) keywords: “fMRI” <OR> “neuroimaging”; AND (3) keywords: “cognit*” <OR> “motor” <OR> “memory” <OR> “attention” <OR> “executive function*” <OR> “language” <OR> “vision” <OR> “ percept.*”
Studies were selected based on the following inclusion criteria: (1) inclusion of a healthy, young adult participant group, (2) neuroimaging data acquired during the same task pretraining and posttraining, and (3) reporting of voxelwise, whole-brain data. Studies investigating functional brain changes during repeated task practice within a single scan session24-27 were excluded as were those comparing performance on a “trained” versus “control” task.28,29 Studies that reported region-of-interest analyses30-32 or did not report activation foci as 3D coordinates in stereotaxic space 33 were also excluded because these were incompatible with the ALE analysis methods.
For studies that reported results for multiple task contrasts using the same group of participants, 34 the contrast reporting the greatest number of foci was selected. This was done to limit the contribution of any 1 set of participants to the pool of foci. For studies containing more than 1 independent sample, peak activation foci from each sample were included. 35 Articles that reported results for pretraining and posttraining phases separately (ie, no direct pretraining vs posttraining comparison) were omitted because GingerALE v 2.0.4 does not compute between-group comparisons with random-effects analysis. As a final step to ensure the comprehensiveness of the review, reference lists of all included articles were searched for additional studies that fit these criteria. In total, 38 appropriate articles were identified for the ALE analysis (a complete list of references for all included studies can be found in the appendix).
ALE Method
ALE was performed using BrainMap Ginger ALE v 2.0.4, 23 a coordinate-based, random-effects meta-analysis for functional neuroimaging data. The ALE method uses a series of permutations to differentiate statistically significant patterns of brain activity from random clustering (ie, noise) of foci, across multiple independent experiments. 23 ALE maps are derived based on foci of interest, which comprise statistically significant peak activation locations from multiple studies. 36
We conducted 2 separate ALE analyses, each yielding an ALE map and corresponding cluster report: (A) posttraining < pretraining and (B) posttraining > pretraining. All studies included in the analysis are listed in Table 1.
Details of Included Studies in the Meta-analysis
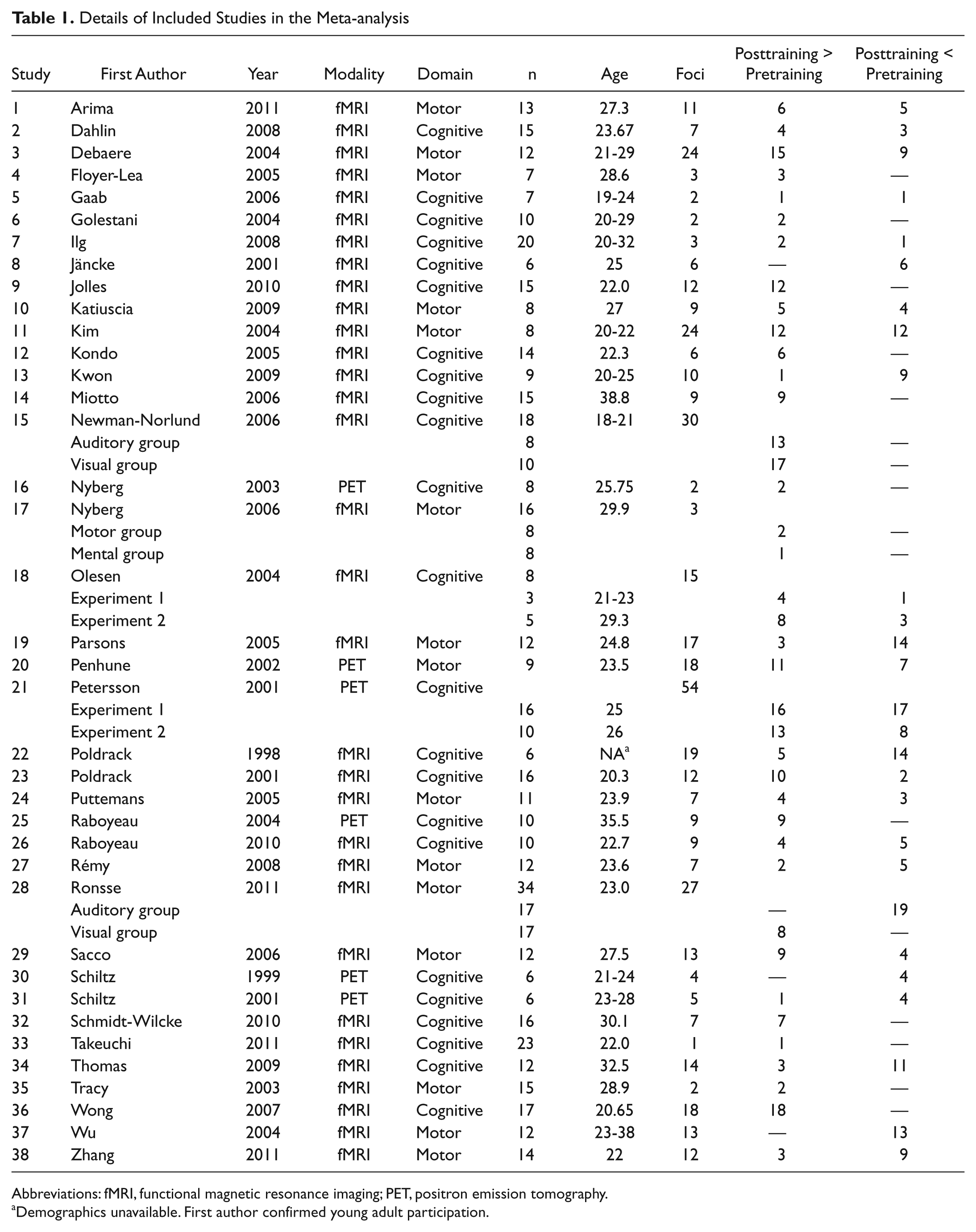
Abbreviations: fMRI, functional magnetic resonance imaging; PET, positron emission tomography.
Demographics unavailable. First author confirmed young adult participation.
All study coordinates were entered in GingerALE in the stereotaxic space of Talairach and Tourneaux. 37 Those studies that originally reported coordinates in Montreal Neurological Institute (MNI) space were transformed into Talairach space using icbm2tal 38 as implemented in GingerALE 2.0.4. Studies that originally reported coordinates in Talairach space derived from Statistical Parametric Mapping (SPM) were transformed back to MNI space using tal2mni and then retransformed into Talairach space using icbm2tal. Additional details regarding these transformations can be found on the BrainMap Web site (www.brainmap.org). Across all studies, activation likelihood estimates (ALE statistics) were computed for each voxel in the brain, reflecting the likelihood that a given voxel was activated across studies in each respective analysis. All reported data survived P < .05 significance, false discovery rate corrected for multiple comparisons, as determined by permutation testing, 23 with a spatial extent greater than 150 mm3. All the data in the figures are presented on a standard template in Talairach space. ALE results images were rendered to cortical surface maps using Caret 39 for presentation.
Results
Behavior
The majority of studies (29/38; 76%) reported statistically significant improvements in task performance following training. All remaining studies reported qualitative data indicating that performance improved with training. Variability in reporting of behavioral data across studies precluded further investigation of the impact of performance gains (eg, effect size estimates) on patterns of functional brain change.
Training-Related Decreases in Brain Activity
The contrast of posttraining < pretraining revealed a number of brain areas associated with reduced activation following training (see Table 2 and Figure 1).
Brain Regions Showing Reliable Decreases in Activity Across Studies Following Training (P < .05, FDR Corrected)
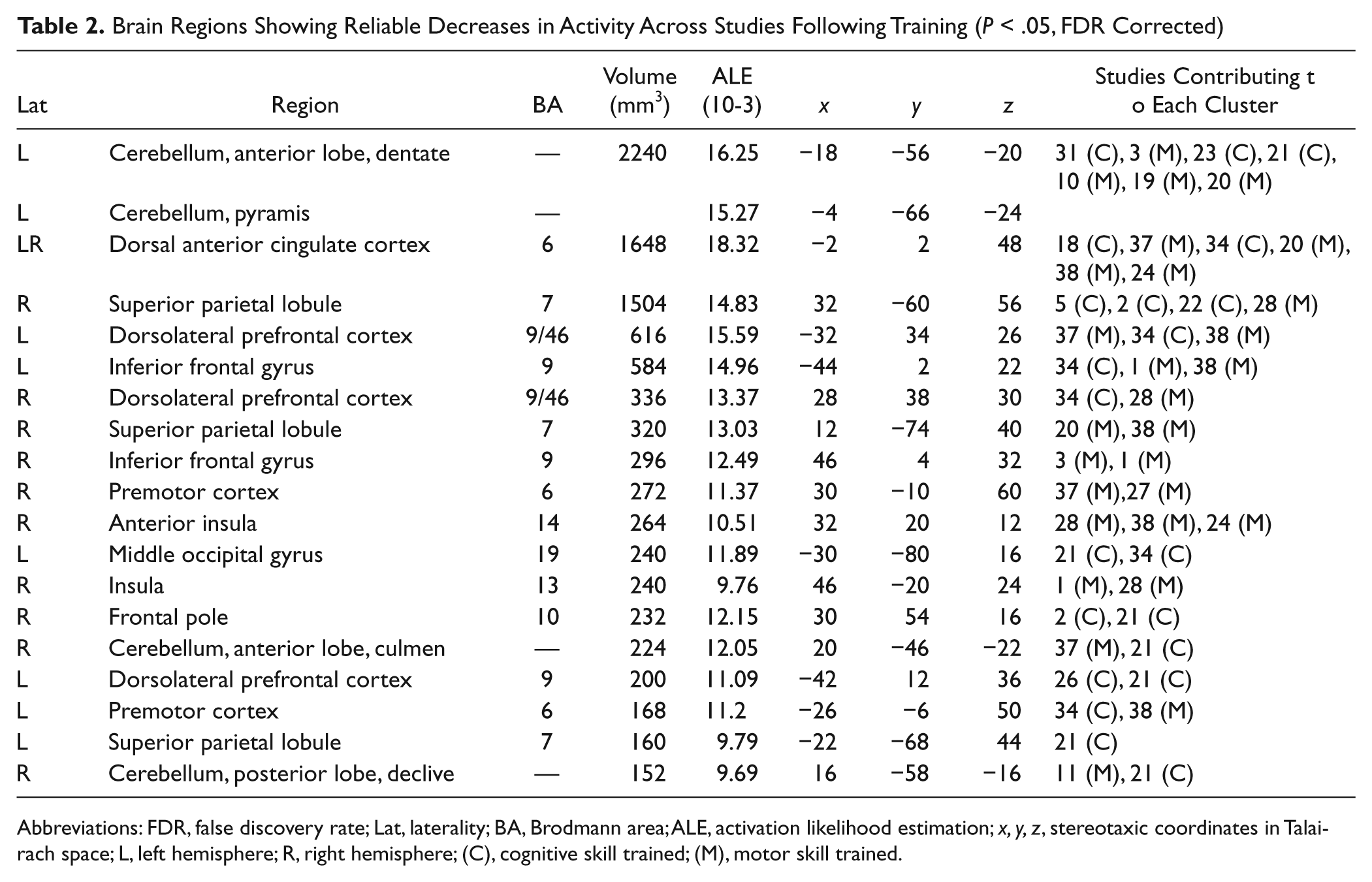
Abbreviations: FDR, false discovery rate; Lat, laterality; BA, Brodmann area; ALE, activation likelihood estimation; x, y, z, stereotaxic coordinates in Talairach space; L, left hemisphere; R, right hemisphere; (C), cognitive skill trained; (M), motor skill trained.
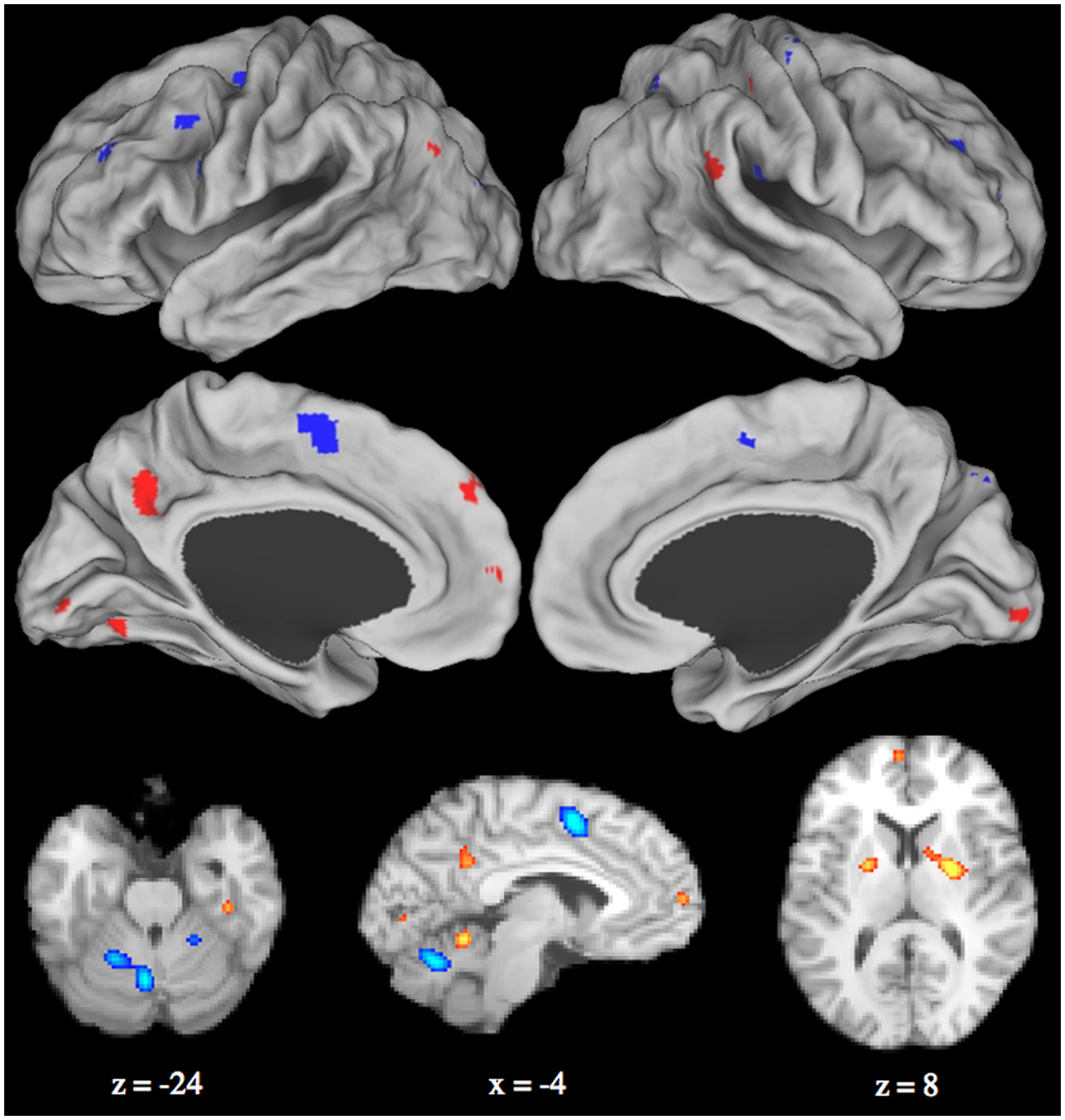
Combined activation likelihood estimation map showing significant activation clusters: red = posttraining > pretraining; blue = pretraining > posttraining. Surface maps of the activation likelihood clusters (false discovery rate P < .05) are shown on an inflated surface map in Caret (Van Essen, 2005, rows 1-2). Cerebellar and subcortical structures are presented in the volume in row 3.
These included bilateral areas of the frontal lobe, including the dorsolateral PFC (Brodmann area [BA] 9/46), inferior frontal gyrus (BA 9), dorsal ACC (BA 6), and the right lateral frontal pole (BA 10). Significant decreases were also observed in the premotor cortex (BA 6) and superior parietal lobule (BA 7) bilaterally. Reduced activation was also recorded in the left dentate subregion of the anterior cerebellum, with clusters of reduced activation extending into the right anterior and posterior lobes. Posttraining decreases were also identified in the right anterior insula (BA 14) and left middle occipital gyrus (BA 19).
Training-Related Increases in Brain Activity
The posttraining > pretraining ALE contrast revealed a number of brain areas associated with increased activation following training (see Table 3 and Figure 1).
Brain Regions Showing Reliable Increases in Activity Across Studies Following Training (P < .05, FDR Corrected)
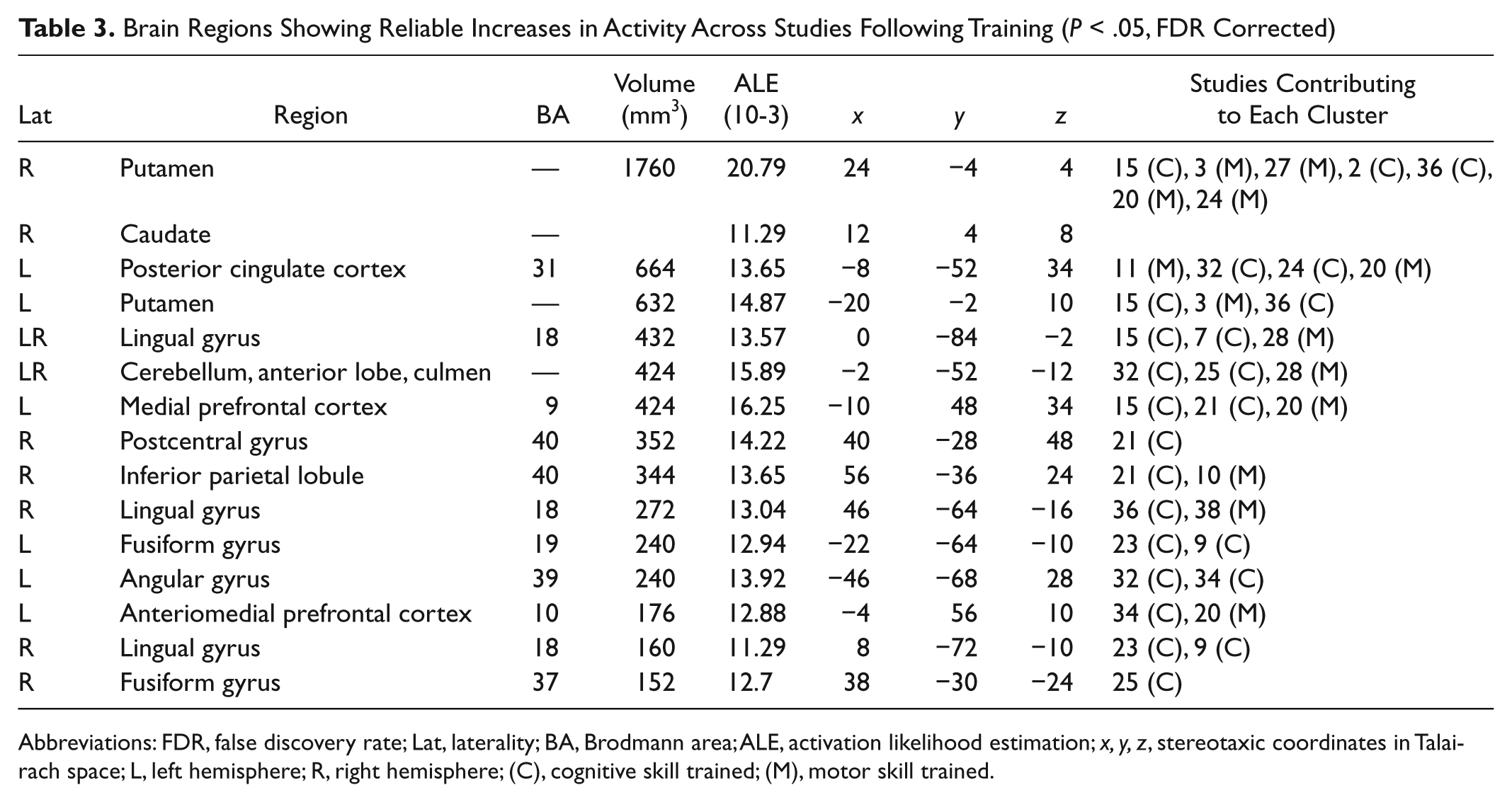
Abbreviations: FDR, false discovery rate; Lat, laterality; BA, Brodmann area; ALE, activation likelihood estimation; x, y, z, stereotaxic coordinates in Talairach space; L, left hemisphere; R, right hemisphere; (C), cognitive skill trained; (M), motor skill trained.
These included the left mPFC (BA 9), left posterior cingulate (BA 31), left inferior parietal lobule (angular gyrus; BA 39), and right postcentral gyrus (BA 40). Posttraining increases were also identified in both the lingual gyri (BA 18) and fusiform gyri (left: BA 19; right: BA 37), bilaterally, as well as the putamen, bilaterally, and right caudate and anterior cerebellum bilaterally.
Contributing Studies by Training Domain
To investigate whether the ALE clusters reflected domain-general training effects across cognitive and motor skills training interventions, we categorized each study as either cognitive (n = 23) or motor (n = 15) skills training (see Table 1) based. Studies were allocated based on the author’s description, the nature of the training task, and outcome measures reported in each study. We then examined the distribution of studies contributing to each ALE cluster (Tables 2 and 3). Many of the ALE clusters had both cognitive and motor training studies contributing (9 of 18 clusters for training-related decreases in brain activity; 9 of 14 clusters for training-related increases in brain activity). For those clusters with 3 or more contributing studies, 5 of 6 clusters for training-related decreases in brain activity were of mixed domain (ie, motor and cognitive), and all clusters were mixed domain for training-related increases in brain activity. For those clusters with only 2 contributing studies, 4 of 11 clusters for analysis training-related decreases in brain activity and 3 of 6 for training-related increases in brain activity had contributing studies from both cognitive and motor domains.
Repeat Scanning and Practice Effects
We were unable to quantitatively parse the contribution of repeat scanning or practice effects to the pattern of brain changes reported here. This limitation reflects an ongoing challenge in the literature examining the functional neural basis of training, where few studies incorporate nontraining control groups. In our sample, 10 of the 38 included studies implemented controlled experimental designs for their neuroimaging analyses. Of these, 5 studies reported a significant group by session interaction,34,40-43 and 2 studies did not observe significant brain changes across the 2 sessions in their control group.44,45 Although not conclusive, a significant majority of the studies in our sample using a controlled group design demonstrated significant brain changes in their treatment group relative to untreated control groups. Thus, the pattern of functional brain changes emerging from our review is unlikely to be significantly attributable to repeat scanning or practice. However, limitations in the literature preclude further investigation of these effects on the ALE results.
Discussion
Using quantitative ALE meta-analytic methods, we reviewed findings from functional neuroimaging studies investigating the neural correlates of training in young adults. Our results demonstrated that training, irrespective of targeted domain, is associated with reduced activation in regions closely overlapping with the frontoparietal control and dorsal attention networks. Specific reductions were observed in dorsal ACC (BA6), dorsolateral and ventrolateral PFC (BA 9/46), anterior insula, and premotor (BA 6) and frontal polar cortices (BA 10). We also observed reductions from pretraining to posttraining in parietal (superior parietal lobule—BA 7) and cerebellar (anterior aspect) brain regions. Training-related decreases in the frontoparietal and dorsal attention networks suggest that training interventions reduce demands on externally focused attention, a finding consistent with a previous quantitative review of the training literature. 9 Notably, these activation decreases were accompanied by increases in default network brain regions, areas typically suppressed during highly demanding tasks. 41 Specific reductions were observed in medial aspects of the PFC (BA 9, 10) as well as the posterior cingulate cortex (BA 31), inferior parietal lobule (BA 40), and angular (BA 39) and postcentral (BA 40) gyri. Our review also demonstrated reliable patterns of increased activation in the striatum (caudate, putamen), consistent with previous reports suggesting that training is associated with increased involvement of subcortical brain regions.3,4 These results implicate relative changes in large-scale functional brain networks as potential markers of training-related gains, irrespective of the training target.
Posttraining reductions in anterior, dorsal, and ventral aspects of the lateral PFC as well as the right dorsal ACC and the left anterior aspect of the inferior parietal lobule are in good agreement with previous reports demonstrating that both motor46,47 and cognitive20,34 skills training lead to reduced activity in frontoparietal control network brain regions. These areas highly overlap with those that comprise the frontoparietal control and dorsal attention networks. These sets of interconnected brain regions are known to be involved in controlled attention and goal-directed processing in response to shifting environmental contingencies. 16 In addition, we observed bilateral reductions in the superior parietal lobule, a region that is reliably engaged during attention-demanding tasks. 15 The distinct functional contributions of these attention-related regions to posttraining performance, however, remain less clear. For instance, the dorsal ACC is known to play a prominent role in performance monitoring by detecting error-prone states. 48 Neuroimaging studies have shown that activation of this region tracks error occurrence, increases with response conflict, 49 and positively correlates with task difficulty. 50 Thus, the dorsal ACC would be expected to be involved in the early stages of learning where effort, conflict monitoring, and flexibility are critical. 51 Reduced activity posttraining may thus reflect lower demand for performance monitoring as skilled performance emerges.
Reduced recruitment of brain regions implicated in attention control reflects a shift from more controlled to automatic forms of task performance following training.3,4,9,10,30,52 Indeed, previous findings have revealed that those classified as “good” learners on a motor sequence learning task exhibited a quicker decrease in activation of prefrontal regions as compared with “poor” performers. 28 Activation decreases across key nodes of the frontoparietal control network may therefore hold predictive value in identifying those individuals who would take longer to reach more automated (ie, less controlled) levels of performance. Training-related automaticity may also decrease the need to suppress task-irrelevant information. 2 That is, reduced engagement of frontoparietal control network regions may attenuate suppression of the default network, allowing for greater task-independent processing.
Reliable training-related increases in brain activation were observed in core regions of the default network, including the posterior cingulate cortex (BA 31), medial PFC (BA9 and BA10), and left posterior inferior parietal lobule (BA 39). The default network has been observed to increase in activation under conditions of low attention demand12,53 as well as decrease in activity as task difficulty increases.54-56 Higher levels of activity within the default network have been associated with greater task ease and mind wandering, 57 whereby participants are able to perform a task well while simultaneously recollecting past events or imagining personal future events, processes supported by the default network. 58 Greater activation of the default network and less demand for controlled attentional processing may free the brain to explore its dynamic repertoire of internal thoughts, memories, and future goals following training. Based on these data, we hypothesize that increased default network activity, combined with reduced frontoparietal control network activity, may provide a candidate neural marker of training-related performance gains.
Our findings provide evidence that relative shifts between large-scale brain network activity may reveal reliable functional brain markers that generalize across a wide range of cognitive and motor skills training interventions. However, the behavioral correlates of changes to the balance among distributed brain networks are just beginning to be understood. Although we were unable to directly evaluate the impact of functional brain changes on behavior in our data, we suggest that brain changes at the level of interacting functional networks may provide important insights into treatment efficacy in both training and neurorehabilitation contexts. Indeed, in 1 recent report, training-related changes in network flexibility—dynamic shifts in the allegiance of nodes to specific brain networks—was shown to be predictive of training-related gains in performance on a motor-learning task. 22
In addition to cortical brain changes, our analyses also revealed posttraining increases in the bilateral putamen and right caudate nucleus. This finding reflects the growing body of studies reporting increased striatal activity after both motor40,59,60 and cognitive34,61,62 skills training. Greater involvement of the basal ganglia after training is thought to reflect a shift in the balance of corticostriatal activity from untrained to trained task performance. 4 In the context of working memory, this may reflect greater engagement of gating functions mediated by basal ganglia to facilitate context-specific and flexible updating of skill representations in the frontal cortex as learning occurs. 63 After training, such flexible updating is no longer necessary, enabling representations of the trained skill to be maintained by the basal ganglia. 34 The idea that the basal ganglia may serve as a “memory store” for well-trained performance is also supported by motor skills training studies showing that the basal ganglia remain active, even when trained performance is tested after a delay. Albouy et al 64 reported that bilateral activation of the putamen was associated with improved performance on an oculomotor sequence learning task over the course of training and was sustained following a 24-hour delay, consistent with a hypothesized role for this region in maintaining representations of trained skills.
The basal ganglia also play an important role in reinforcement learning because dopaminergic neurons in the striatum respond to stimuli that predict award and adjust their firing rate according to the magnitude of the expected reward.65-67 Training may therefore serve to strengthen stimulus-response associations by promoting successful behaviors that are reinforced through reward signaling mediated by corticostriatal circuits. Indeed, recent animal findings demonstrated that blockage of the anterior forebrain pathway, a cortical-basal ganglia circuit, during training of a complex motor skill disrupted gradual skill improvement that normally occurs with training. 68 Unblocking these channels posttraining reinstantiated learning. This suggests a crucial role for corticostriatal reward circuits in supporting and maintaining representations of trained skills. Indeed, it is now well understood that aspects of the frontoparietal control network, particularly regions of the lateral PFC, are highly functionally connected with the basal ganglia69,70 and that these connections are altered with training. 34 Although we are unable to address this directly here, the results of this meta-analysis raise the intriguing possibility that alterations in cortical-subcortical functional connectivity may serve to modulate the dynamic interactions among cortical brain networks. Investigating how these large-scale, distributed brain networks interact and how these interactions are modulated by training, aging, brain disease, or injury represent important areas of future inquiry.
Our meta-analysis also demonstrated that training was associated with a large cluster of reduced activity in the dentate subregion of the anterior cerebellum. This parallels previous findings showing reduced activity in the dentate nucleus after extensive practice on a motor sequence task. 46 We also observed a smaller cluster of increased activity in the anterior region of the cerebellum, consistent with hypothesized involvement of this region in highly trained motoric performance. 71 However, activation increases in anterior cerebellar regions are often transient, potentially reflecting rapid learning and adaptation processes not fully captured in the extended training paradigms reviewed here. Moreover, both motor and cognitive studies contributed to these clusters. Training-induced functional cerebellar changes may reflect both the degree and speed with which automaticity is attained over the course of training, and this may vary across motor and cognitive tasks. Further work will be needed to determine how the composition of training tasks influence automaticity and how this is reflected in activation changes in cerebellar subregions.
Our findings that training is associated with changes in large-scale cortical and subcortical brain networks may have important implications for training and rehabilitation intervention design. In the context of neurorehabilitation, Chen and colleagues 1 recently identified a neural marker of goal-directed attention that was significantly altered by neurorehabilitation training in a sample of patients with acquired brain injury. They observed functional changes in distributed brain regions purported to mediate goal-directed behavior that were associated with significant performance gains on behavioral measures of attention regulation. The authors also reported that baseline PFC activation, a purported mediator of attentional regulation, predicted posttraining changes in this region, a potential marker of treatment readiness not accessible through behavioral data alone. Similarly, Strangman and colleagues 72 observed that moderate baseline prefrontal activity was associated with greater treatment gains than either high or low baseline PFC response. Although much work remains, these data suggest that neural markers obtained at baseline may predict response to neurorehabilitation interventions, thus informing treatment planning and potential triaging of patients to appropriate training protocols. Moreover, changes in neural response from pretraining to posttraining may also provide informative outcome data regarding mechanisms of treatment efficacy that are not accessible through standard behavioral measures.
Markers of functional brain response can also be used to interpret negative training outcomes. 60 This later contribution of functional brain data to interpreting negative training outcomes was recently highlighted in a study by Dahlin and colleagues. 34 The authors reported that older adults failed to show transfer of training from one working memory task to another. Both tasks had been shown to recruit overlapping areas in the subcortical striatum, and this overlap was hypothesized to be important for mediating training transfer between working memory tasks in younger adults. Older adults’ failure to engage these striatal brain regions posttraining was posited to underlie their inability to generalize training gains, suggesting a potential role for striatal brain regions in mediating training transfer among working memory tasks. Here functional brain data were used to identify a potentially important neural mechanism critical for training transfer in a specific cognitive domain. This mechanism could be targeted in future training and treatment protocols, again highlighting the importance of neural markers in treatment design and evaluation.
Our findings suggest that changes in the activation of large-scale functional brain networks may provide important neural markers of training-related performance gains irrespective of training domain or specific training task. Changes in distributed functional networks from pretraining to posttraining may thus inform the development of reliable, domain-general markers of treatment response in training and neurorehabilitation settings. Mapping the intrinsic neural architecture of the regional connections comprising the default, frontoparietal control and dorsal attention networks is under way. 73 Determining training-related changes in distributed network activation, in addition to interregional coupling of activity, may represent a critical step toward developing functional neural markers to elucidate the mechanisms and evaluate the efficacy of neurorehabilitation interventions.
Footnotes
Appendix
Acknowledgements
We thank Julia Spaniol and Lixia Yang for their helpful comments on earlier versions of this manuscript. We apologize to any authors whose work was mistakenly overlooked.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: GRT recognizes the Heart and Stroke Foundation of Canada, Centre for Stroke Recovery and the Faculty of Health, York University, ON, Canada, for their generous support.
†
The “wildcard” asterisk (*) was used to retrieve search words with multiple endings (train, training, trained, etc).