
Abstract
Background. Functional outcomes are generally poor following peripheral nerve injury (PNI). The reason is multifactorial but includes the misdirection of regenerating axons to inappropriate end organs. It has been shown that brief electrical stimulation (Estim) of nerves has the potential to improve the accuracy and rate of peripheral axon regeneration. Objective. The present study explores a novel percutaneous transvertebral approach to Estim, which was tested in the mouse femoral nerve model. Methods. Inspired by the protocol of Gordon and colleagues (ie, 20 Hz, for 1 hour), we applied Estim to the cervicothoracic spinal cord (SC-Estim) to remotely activate lumbar motor neurons following transection and repair of the femoral nerve. Fluorescent dyes were applied to the distal nerve to label reinnervating cells. Sections of nerve were taken to quantify the numbers of reinnervating axons as well as to stain for a known femoral axon guidance molecule—polysialylated neural cell adhesion molecule (PSA-NCAM). Results. In comparison to sham treatment, SC-Estim led to significantly greater expression of PSA-NCAM as well as improved the specificity of motor reinnervation. Interestingly, although SC-Estim did not alter the number of early reinnervating (ie, pioneer) axons, there was a reduction in the number of retrogradely labeled neurons at 2 weeks postrepair. However, by 6 weeks postrepair, there was no difference in the number of neurons that had reinnervated the femoral nerve. Conclusions. The present findings support the development of SC-Estim as a novel approach to enhance the specificity of reinnervation and potentially improve functional outcomes following PNI.
Keywords
Introduction
Peripheral nerve injuries (PNIs) are remarkably common and can result from a variety of causes ranging from penetrating or closed trauma to compression by mass lesions and drug toxicity. Unlike axons in the brain and spinal cord, peripheral axons retain some capacity to regenerate after injury. This has led to the misconception that patients who suffer a PNI should expect a favorable outcome. In fact, the opposite is probably much closer to the truth. For example, the majority of patients who suffer traumatic PNI wait months to years for a recovery of function that is generally incomplete. 1 In many cases, regenerating axons fail to reestablish specific connections with their appropriate end organs, which is a critical step required for the restoration of normal sensory and motor performance. 2 Therefore, the development of strategies aimed at increasing the specificity of axon regeneration is an important step toward achieving better clinical outcomes after PNI.
For many decades, developmental biologists have dissected the cellular and molecular mechanisms of axon guidance through the application of simple, yet elegant model systems. In contrast, well-described models for the study of axon guidance in the mature mammalian nervous system are limited. To this end, Brushart3,4 developed the rodent femoral nerve model, in which regenerating motor axons reproducibly demonstrate selective targeting behavior. This phenomenon is commonly referred to as preferential motor reinnervation (PMR). In brief, the femoral nerve is sectioned and repaired before it bifurcates into similarly sized distal branches to the quadriceps muscles and purely sensory saphenous nerve. Following a period of recovery, a second surgery is performed to apply fluorescent dyes to the distal branches in order to retrogradely label the motor neurons (MNs) that either (1) correctly reinnervated the motor pathway or (2) incorrectly reinnervated the sensory pathway. Because these pathways are originally supplied by a comparable number of myelinated axons, 3 simple statistical analysis can be applied to this binary targeting choice. Permutations of the PMR paradigm have been described in rats,3-6 mice,7-9 and primates. 10
Brief electrical stimulation applied to the proximal end of transected peripheral nerves (ie, nerve-Estim) has been shown to augment PMR,5,11,12 improve the specificity of sensory reinnervation,12,13 and accelerate the reinnervation of distal target tissues.5,12,14-32 These effects appear to be dependent on the depolarization of the neuronal soma itself and not just its axon.5,26 Several axon guidance factors have been implicated, including polysialylated neural cell adhesion molecule9,11 (PSA-NCAM), the L2/HNK-1 carbohydrate,33-35 and brain-derived neurotrophic factor.12,26,33,36
In the present study, we explored whether activation of central pathways that input onto lower MNs could replicate the effects of nerve-Estim on peripheral axon regeneration. Specifically, we used the mouse femoral nerve model to describe how a novel percutaneous, transvertebral approach for brief Estim of the rostral spinal cord (ie, SC-Estim) improves the specificity of motor axon reinnervation.
Parts of this article have been published previously in abstract form. 25
Methods
Mice
A total of 69 young adult (8-12 weeks old) male C57Bl/6 mice were obtained from Charles River (Wilmington, Massachusetts) for use in these experiments. Mice were maintained in a 12/12 light/dark cycle with ad libitum access to food and water. All protocols involving animals were approved by the University of Calgary Animal Care Committee in accordance with the policies of the Canadian Council of Animal Care (CCAC).
Surgery
Mice were anesthetized with 2% isoflurane (Baxter, Toronto, Canada), and surgery was performed under aseptic conditions. The femoral nerve was carefully exposed with blunt dissection after a small incision was made in the overlying skin of the inguinal area. The transection of the femoral nerve was made approximately 2 mm proximal to its bifurcation into the quadriceps muscle branch and saphenous (cutaneous) nerve as previously described in mice by Franz et al. 9 The proximal and distal stumps were then microsurgically repaired with a single 11-0 nylon suture (FST, North Vancouver, Canada).
Electrical Stimulation
Immediately following repair of the femoral nerve, mice were put into a prone position and needle electrodes were positioned on either side of the spine, 10 mm caudal to the occiput, with the neck hyperextended, at the level of the cervicothoracic junction. Estim was applied transvertebrally using percutaneous needle electrodes connected to a SD9 stimulator (Grass Technologies, West Warwick, Rhode Island) set at 20 Hz, with a 100-µs pulse duration, for 1 hour, as previously described by Gordon and colleagues for direct Nerve-Estim. 5 The stimulus was adjusted to 1.5 times the threshold required to evoke visible quadriceps muscle contraction in the contralateral limb, which ranged from 3 to 5 mV. The STIM group underwent the SC-Estim treatment, but the SHAM group underwent only placement of the percutaneous electrodes for 1 hour.
Immunohistochemistry
For analysis of c-Fos expression, the L2-4 SC was isolated and dissected free from the spine of mice that were transcardially perfused with 4% paraformaldehyde (PFA) 1 hour following SC-Estim (n = 3) or sham treatments (n = 3). The SC segments were cryoprotected overnight in 30% sucrose in phosphate-buffered saline (PBS), immersed in OCT compound (Sakura Finetechnical Co, Torrance, California), flash frozen in dry-ice-cooled isopentane, and cryostat sectioned in the transverse plane at 20 µm. Sections from STIM and SHAM were collected in adjacent rows on the same Superfrost plus slide (Fisher Scientific, Nepean, Canada) and air dried overnight. Slide-mounted SC cross sections were incubated in 0.1% Triton X-100, rinsed, and preblocked in 2% bovine serum albumin (BSA) in PBS for 60 minutes at room temperature (RT). Sections were incubated in rabbit anti-c-Fos antibody (1:500; Abcam, Cambridge, Massachusetts) diluted in PBS containing 2% BSA at 4°C for 12 hours. Next, the slides were washed 3 times in PBS (10 min/wash) and then incubated in Alexa 555 conjugated goat antirabbit IgG antibody (1:1000; Invitrogen) diluted in 2% BSA for 1 hour at RT. The slides with SC sections were washed again with PBS and mounted with FluorSave reagent (Calbiochem, San Diego, California) for fluorescence microscopy. Stained SC sections were visualized under an upright fluorescent microscope (Olympus BX51, Center Valley, Pennsylvania), and images were captured with a Cool Snap Pro digital camera operated by Image Pro Plus software (Media Cybernetics, Silver Spring, Maryland).
To examine the levels of PSA-NCAM, a known axon guidance molecule in both the development and regeneration of the peripheral nervous system, 37 we transcardially perfused mice with 4% PFA 5 days after nerve transection and repair. A 3-mm segment of femoral nerve immediately proximal to the repair site was dissected free, cryoprotected in 30% sucrose at 4°C and then embedded in OCT compound (Sakura Finetechnical Co). Nerves were sectioned in the horizontal plane at 14 µm using a cryostat with sections from control unoperated (CON; n = 7), SHAM (n = 7), and STIM (n = 7) treated mice collected in adjacent rows on the same Superfrost plus slide (Fisher Scientific).
Slide-mounted nerve cross sections were incubated in 0.1% Triton X-100, rinsed, and preblocked in 2% BSA in PBS for 60 minutes at RT. Sections were incubated in the monoclonal anti-PSA IgM antibody (1:105, 5A5 ascites fluid; Developmental Hybridoma Bank, Iowa) plus mouse anti-Tuj1 antibody (1:500, Covance, Berkley, California) and mouse anti-NF200 antibody (1:500, Sigma, Oakville, Canada) in PBS containing 2% BSA at 4°C for 24 hours. Next, the slide with nerve sections was washed 3 times in PBS (10 min/wash) and then incubated in Alexa 594 conjugated goat antimouse IgM antibody (1:1000; Invitrogen) and Alexa 488 conjugated goat antimouse IgG antibody (1:1000; Invitrogen) in 2% BSA for 1 hour at RT. The slide with nerve sections was washed again with PBS and mounted with FluorSave reagent (Calbiochem) for fluorescence microscopy.
Stained nerve sections were visualized under an upright fluorescent microscope (Olympus BX51), and images were captured with a Cool Snap Pro digital camera operated by Image Pro Plus software (Media Cybernetics). For these double-labeling experiments, controls included omission of one or both primary antibodies. To compare changes in the expression of PSA-NCAM, 2 channels of fluorescence were captured and aligned per the procedures of Hanson and Landmesser. 38 Anti-Tuj1 + NF200 were acquired in the first channel and acted as control for potential variations in section thickness, nerve orientation, and the number of axons within each section. The second channel imaged was anti-PSA-NCAM. Pixel intensity (arbitrary units) was measured within, but not including, the outer epineural layer that defined each nerve cross section using Image J (National Institute of Mental Health, Bethesda, Maryland). The normalized pixel intensity was calculated by dividing the mean values of PSA-NCAM by Tuj1 + NF200 for each condition. Images of representative nerve sections were acquired and analyzed in a blinded fashion.
MN and Dorsal Root Ganglion Quantification
After either 2 (n = 7/group) or 6 (n = 14/group) weeks, a second operation was performed to assess the accuracy of motor axon regeneration in SHAM- and STIM-treated mice. The distal muscle (Mu) and cutaneous (Cu) branches of the femoral nerve were transected 5 mm distal to their bifurcation point, and 10% fluororuby or fluoroemerald (Invitrogen, Burlington, Canada) was randomly applied to either branch to allow quantification of the number of reinnervating cells, as previously described. 9 The mice were given 7 days to recover for maximal neuron labeling, 39 then deeply anesthetized and transcardially perfused with 4% PFA prepared in PBS. The lumbar SC along with the L2-4 dorsal root ganglia (DRG) were dissected free, postfixed in 4% PFA overnight, cryoprotected in 30% sucrose in PBS, flash frozen, and then sectioned with a cryostat. SCs were cut longitudinally at 60 µm, whereas DRG were cut at 10 µm. Labeled MNs were counted as described previously 9 and raw counts were corrected by the method described by Abercrombie. 40 Labeled DRG were counted in every second section to minimize the likelihood for double counting large cells 13 (note that DRG were only collected at the 2-week time point; n = 5/group). All cell counts were performed by a blinded observer who was unaware of the treatment group and which dye had been applied to either branch.
Myelinated Axon Analysis
The 2-mm segment of femoral nerve immediately distal to the repair site was dissected from the thigh following transcardial perfusion with 4% PFA at the 2-week time point only (n = 5/group). Nerve specimens were postfixed in 2.5% glutaraldehyde buffered in 0.025 M cacodylate overnight, washed, and stored in 0.15 M cacodylate buffer, fixed in 2% osmium tetroxide, washed in graded alcohols, and then embedded in Epon. 41 Cross sections of thickness 1 µm were made through the proximal 2 mm of the reinnervated nerve stump, collected on to a slide, stained with toluidine blue, and mounted with Permount (SP15-100, Fisher Scientific). Stained sections of nerve were sampled in the center of each quadrant as nonoverlapping photomicrographs were captured at 100× magnification under oil immersion microscopy (Olympus BX51). In each field, the density of myelinated axons was determined using Image J (National Institute of Mental Health, Bethesda, Maryland), and the average axon density was then multiplied by the total cross-sectional area of the nerve to estimate the total number of myelinated axons distal to the nerve repair.
Statistical Analysis
Mean values (±standard error of the mean; SEM) are reported throughout. The Student t test (unpaired) was used to make comparisons between MN and axon counts (P critical = .05). A 1-way analysis of variance (ANOVA) was used to make comparisons between normalized PSA-NCAM levels. Tukey honestly significant difference post hoc test was used to determine where the significant differences occurred only if the F value exceeded F critical.
Results
SC-Estim Activates Lumbar MNs
Single pulses of supramaximal Estim (3-5 mV, 100 µs) were applied with percutaneous electrodes placed at the junction of cervical and thoracic spinal cord of mice anesthetized with isoflurane (Figure 1A). Application of SC-Estim at this remote site was able to activate femoral MNs in the lumbar spinal cord as detected via compound muscle action potentials recorded from the vastus lateralis muscle (Figure 1B). Sensory nerve action potentials could also be recorded in hind paws with single pulses of SC-Estim, which presumably represented antidromic activation of dorsal column sensory pathways (data not shown). As expected, the complete ablation of the compound muscle action potential response was achieved by transecting the femoral nerve (Figure 1C). Furthermore, when SC-Estim was applied repetitively at 20 Hz for 1 hour (ie, STIM), there was widespread upregulation of the c-Fos immediate early gene, predominantly in the lumbar SC neurons, when compared with SHAM-treated controls (n = 3/group; Figures 1D and 1E).
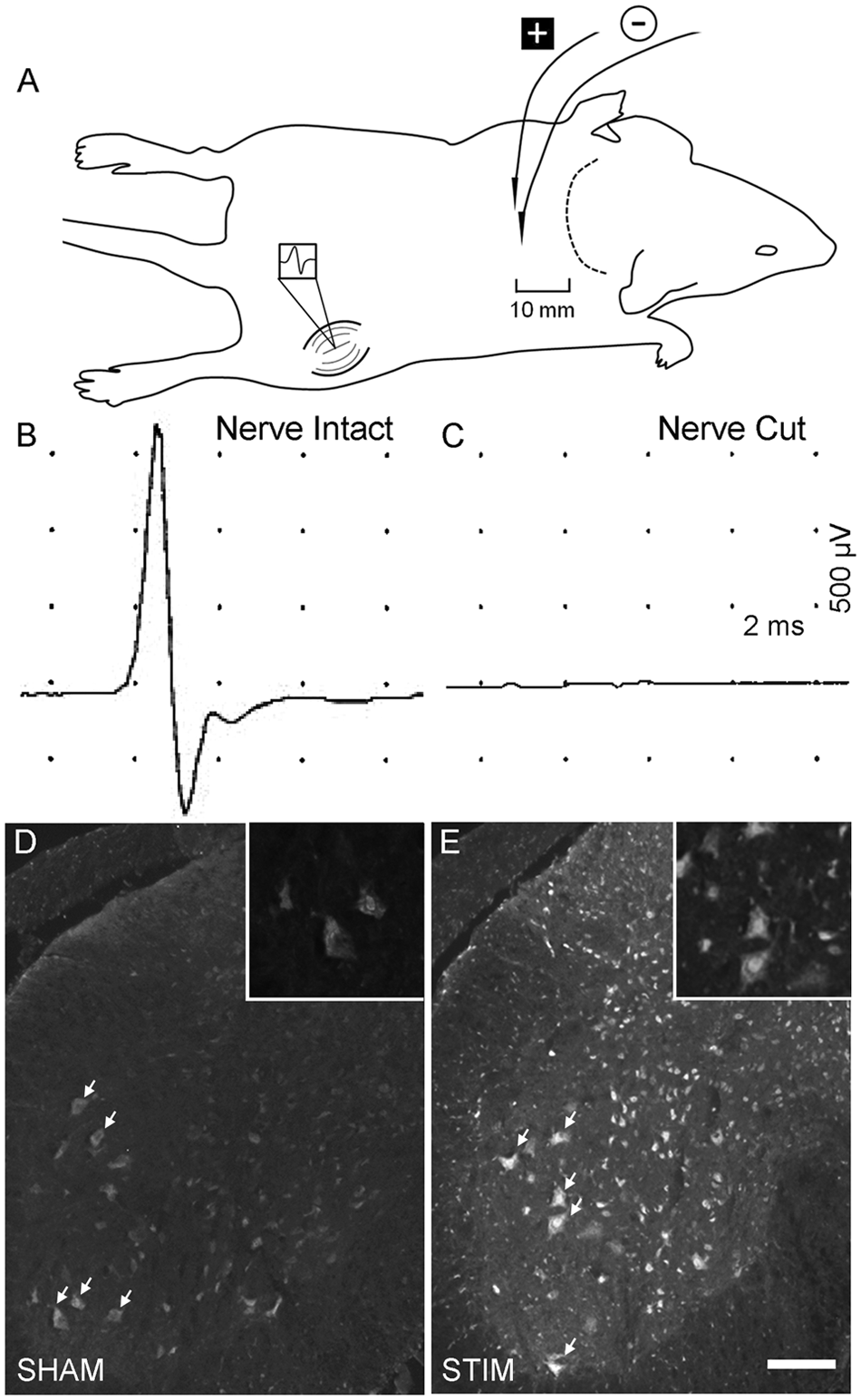
Rationale and procedure for brief SC-Estim. A. Sketch depicting the placement of stimulating electrodes flanking either side of the spine near the cervicothoracic junction. EMG recordings were made from the vastus lateralis muscle, which is supplied by the femoral nerve. B. Compound muscle action potential is shown following a single pulse (5 mV, 100 µs) delivered as depicted in A. C. Following the femoral nerve cut (in the same mouse), the EMG response was completely abolished. D and E. Representative photomicrographs of c-Fos staining in the lumbar spinal cord from a SHAM-treated mouse (D) as well as a mouse that received STIM treatment, with high-power insets. Qualitative assessment revealed a more widespread, cellular elevation of c-Fos in STIM vis-à-vis SHAM stimulation (n = 3/group). Arrows identify motor neurons. Scale bar, 250 µm and 100 µm (for insets).
SC-Estim Increases PSA-NCAM Levels
PSA-NCAM is an axon guidance molecule involved in neuromuscular development42-46 as well as femoral nerve regeneration.9,11 Because of its highly localized expression on the distal portion of regenerating axons, we applied a semiquantitative immunofluorescent approach to compare its expression. 11 As shown in Figure 2, compared with uncut CON femoral nerve (1.02 ± 0.02, n = 7), both the SHAM (2.39 ± 0.30, n = 7) and STIM (3.57 ± 0.45, n = 7) demonstrated significant upregulation of PSA-NCAM (P < .05 and P < .01, respectively). Furthermore, there were significantly higher levels of PSA-NCAM expressed on STIM as compared with the SHAM nerve (P < .05).
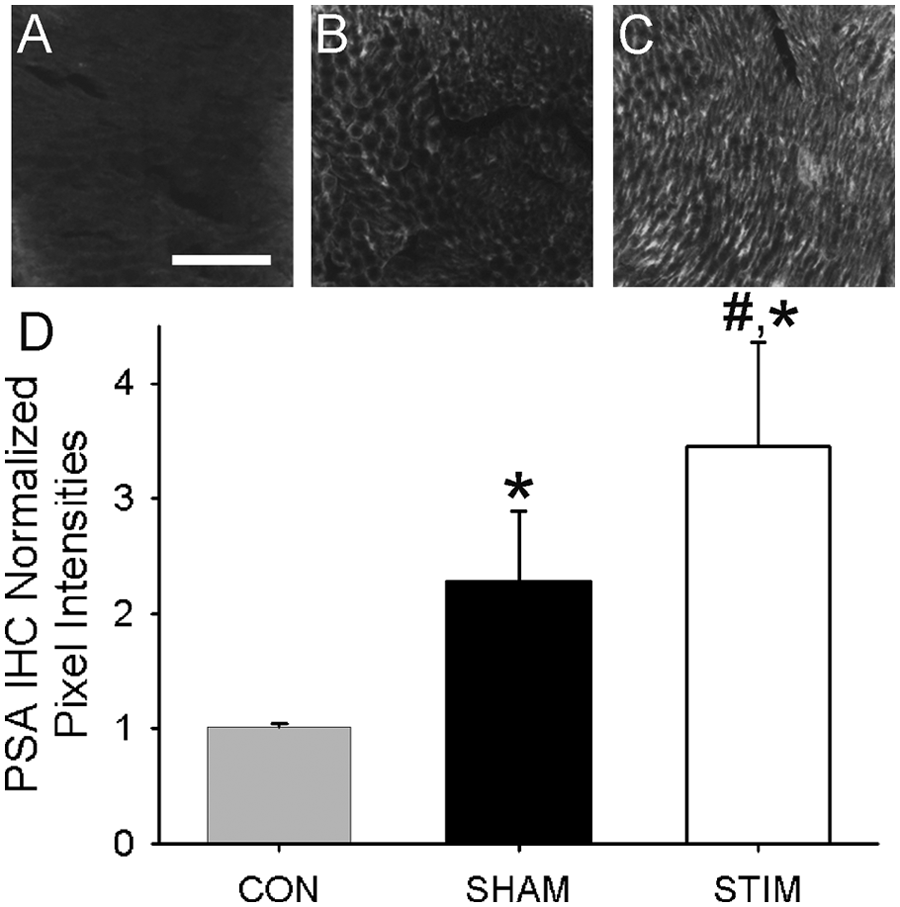
SC-Estim increased the levels of PSA-NCAM in transected femoral nerves. (A-C) Representative photo-micrographs of PSA-NCAM staining in the femoral nerve cross- section immediately proximal to repair level from a CON (unoperated; A), SHAM (B), and STIM (C) mouse. Qualitative assessment revealed upregulation of PSA-NCAM 5 days post–nerve repair relative to CON with the greatest staining seen in the STIM condition. (D) Semiquantitative analysis of normalized fluorescent pixel intensity was compared between CON, SHAM, and STIM groups. The SHAM group demonstrated significantly greater PSA-NCAM levels than CON (P < .05). The STIM group had significantly greater PSA-NCAM levels as compared with both CON (P < .001) and SHAM (P < .01). Scale bar, 100 µm.
SC-Estim Delays Reinnervation
To compare the rate of distal nerve reinnervation, the femoral nerve was cut and repaired. Following 2 or 6 weeks of recovery, neuron labeling was performed and ultimately nerve, DRG, and SC specimens were obtained for analysis (see Methods section). The number of labeled MNs in the SHAM (94 ± 12, n = 7) as compared with the STIM group (57 ± 10, n = 7) was significantly higher at 2 weeks (P < .05; Figure 3A). By 6 weeks of recovery, when the number of regenerating MNs had plateaued, 9 there was no difference in the number of labeled MNs between SHAM (142 ± 13, n = 14) and SC-Estim (144 ± 12, n = 14). The number of labeled DRG neurons in the SHAM (534 ± 60, n = 5) as compared with the STIM (320 ± 62, n = 5) was also significantly greater (P < .05; Figure 3B). However, there was no difference between SHAM (1326 ± 205, n = 5) and STIM (1096 ± 230, n = 5) in the number of myelinated axon profiles counted 1 to 2 mm distal to the site of nerve repair (P = .43; Figure 3C).
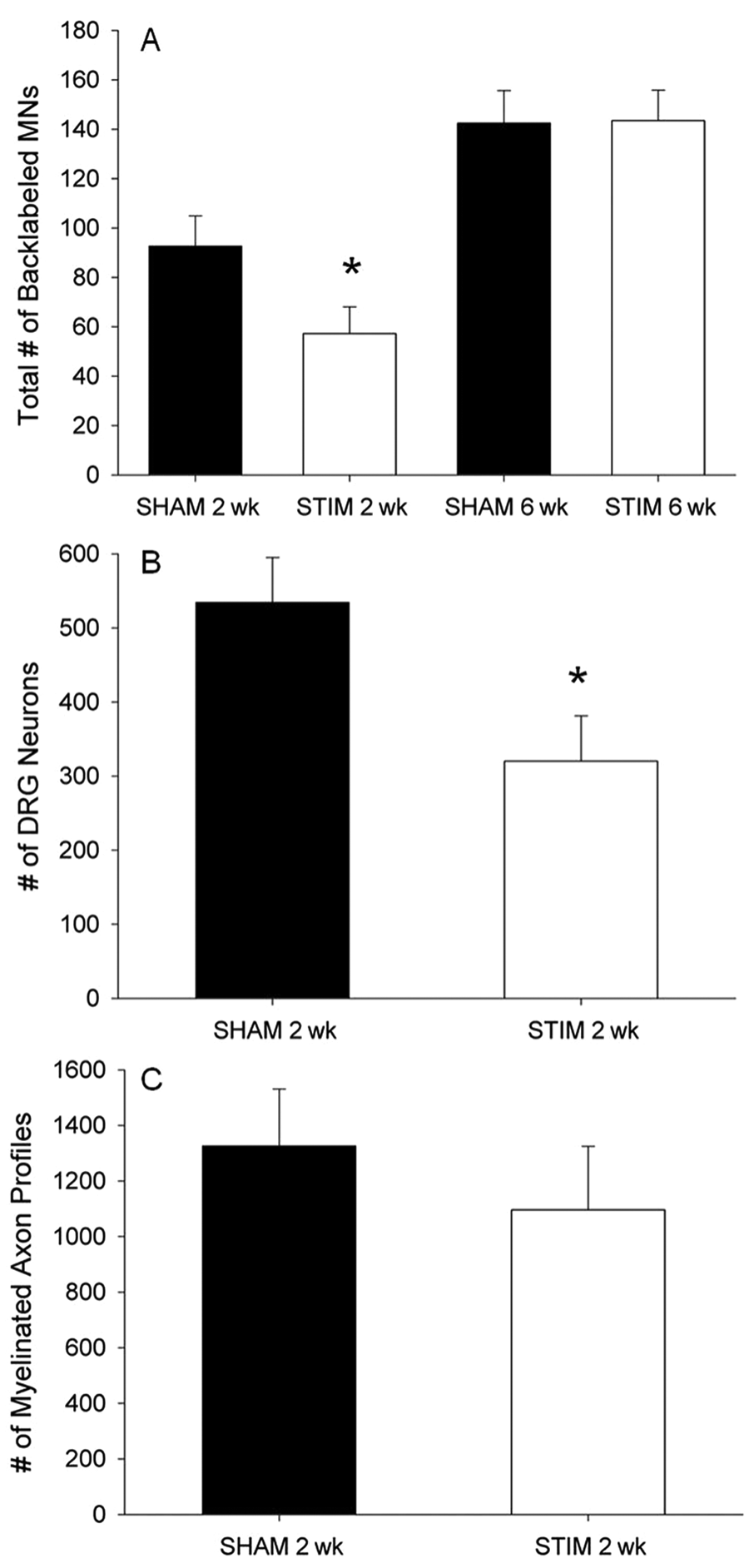
SC-Estim delayed the reinnervation of the distal femoral nerve. (A) The total number of backlabeled MNs that picked up tracer from either the distal muscle or cutaneous pathway in SHAM versus STIM groups at 2 weeks and 6 weeks post–nerve transection and repair. There were significantly fewer labeled MNs in STIM as compared with SHAM at 2 weeks (P < .05), but by 6 weeks, regeneration was complete, and there was no longer a difference between the groups. (B) The total number of backlabeled DRG that picked up tracer from either the distal muscle or cutaneous pathway in SHAM versus STIM groups at 2 weeks post–nerve transection and repair. There were significantly fewer labeled DRG in the STIM as compared with SHAM at 2 weeks (P < .05). (C) The total number of myelinated axons 1 to 2 mm distal to the repair site at 2 weeks post–nerve transection and repair. There was no difference in the number of myelinated axons distal to repair between the groups. *P < .05.
SC-Estim Improves Regeneration Accuracy
The accuracy of regeneration into the distal femoral nerve by axotomized MNs was assessed with a retrograde tracing technique adapted from Brushart. 4 As expected, at 2 weeks of recovery, neither the SHAM nor STIM groups reinnervated the distal nerve pathways with any preference for the muscle pathway (Figure 4A). In the SHAM group, the number of labeled MNs that reinnervated the muscle branch was 36 ± 11, the cutaneous branch was 50 ± 12, or both branches was 6 ± 2 (n = 7). In the STIM group, the number of labeled MNs that reinnervated the muscle branch was 22 ± 4, the cutaneous branch was 32 ± 8, or both branches was 3 ± 1 (n = 7).
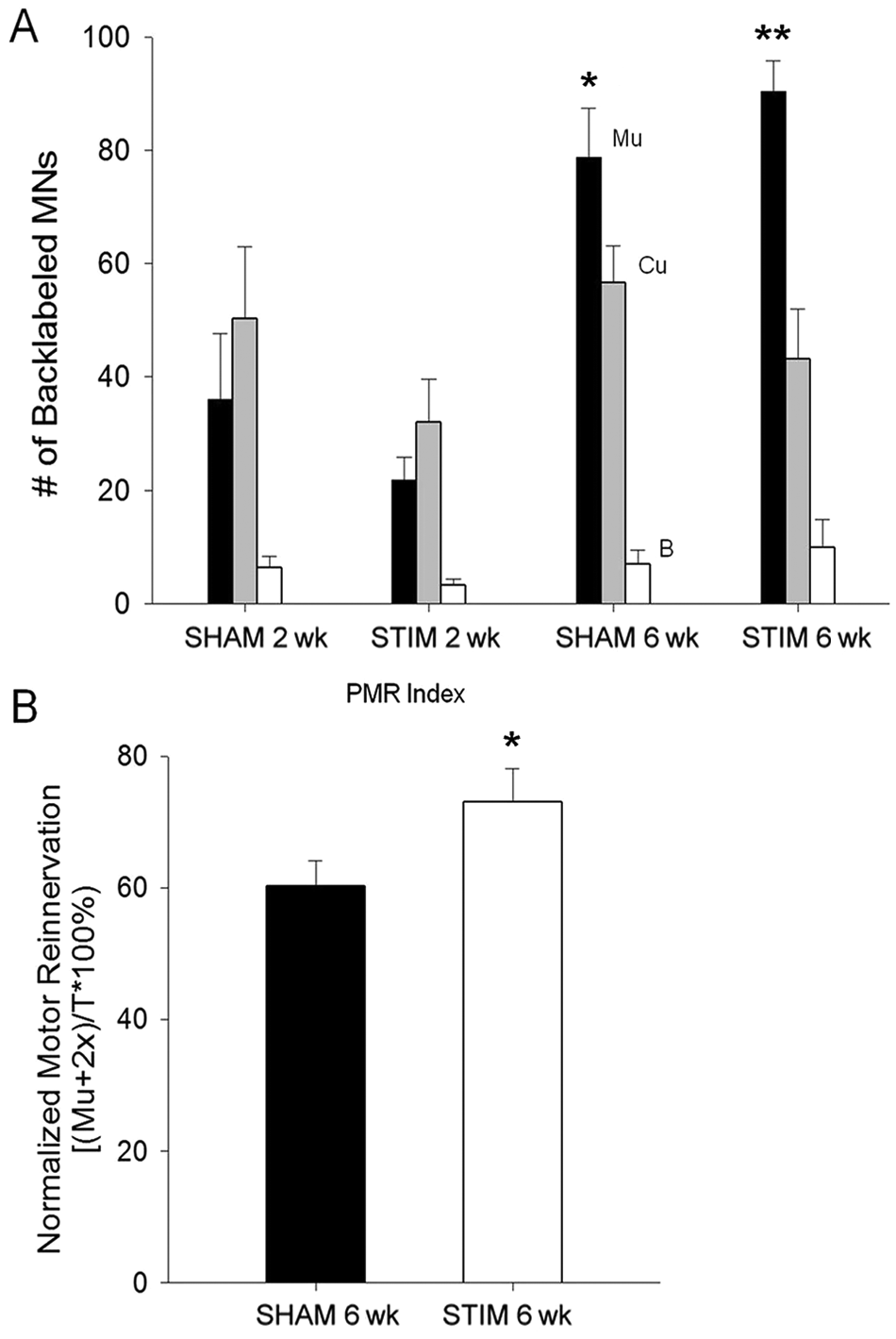
Motor axon regeneration accuracy was improved by SC-Estim. (A) The number of backlabeled MNs that picked up tracer from muscle pathway (Mu), cutaneous pathway (Cu), or both (B) at 2 weeks and 6 weeks post–nerve transection and repair. Neither group demonstrated a preference to reinnervate either the Mu or Cu pathway at 2 weeks, but by 6 weeks both the SHAM (P < .05) and STIM (P < .001) demonstrated preferential reinnervation of the Mu pathway. (B) Normalization of all MNs that reinnervated the Mu pathway (Mu+B), to the total number of MNs, revealed a significantly greater extent of Mu pathway reinnervation in the STIM versus SHAM group (P < .05). *P < .05; **P < .001. Abbreviations: SC-Estim, spinal cord electrical stimulation; MN, motor neuron; SHAM, sham group; PMR, preferential motor reinnervation.
By 6 weeks of recovery, both groups demonstrated PMR (Figure 4A). In other words, the number of MNs that correctly reinnervated the muscle branch was significantly greater than the cutaneous branch for both the SHAM (P < .05) and STIM (P < .001) groups. In the SHAM group, the number of labeled MNs that reinnervated the muscle branch was 79 ± 9, the cutaneous branch was 57 ± 7, or both branches was 7 ± 2 (n = 14). In the STIM group, the number of labeled MNs that reinnervated the muscle branch was 90 ± 5, the cutaneous branch was 43 ± 9, or both branches was 10 ± 5 (n = 14). When the number of MNs labeled with the muscle pathway tracer was normalized to the total number of labeled MNs (Figure 4B), a significantly greater percentage was seen in the STIM (73 ± 5) as compared with the SHAM (60 ± 4) condition (P < .05).
Discussion
There is strong evidence that brief Estim applied to regenerating peripheral nerves (ie, nerve-Estim) in rodents accelerates the process of reinnervation.5,23,24 This acceleration appears to be a consequence of more rapid axonal growth across the site of surgical anastomosis rather than an increase in the rate of axonal elongation per se.26,32 The finding of accelerated reinnervation after nerve-Estim has been reproduced in numerous studies from multiple research groups that have applied several different protocols12,14-32; however, the most widely studied protocol was developed by Al-Majed et al. 5 A recent clinical trial has even supported the idea that this protocol, combined with surgery, augments muscle reinnervation in patients with moderate to severe carpal tunnel syndrome. 47 However, despite finding an increased number of motor units after nerve-Estim, Gordon and colleagues did not detect any measurable improvement in hand function.
Although the factors underlying recovery of function after nerve injury are certain to be multifactorial, one of the major determining factors is widely accepted to be the specificity of reinnervation. The debate on what outcome measure(s) best predict functional recovery after nerve injury is far from settled, but Brushart recently reviewed the issue and found the specificity of reinnervation was a better predictor of functional outcomes than either the reinnervation of the distal nerve or the end organs. 48 That said, there are conflicting results on whether nerve-Estim augments or impairs the specificity of reinnervation. When applied directly to the nerve5,11-13 or to the SC (Figure 4), Estim improves the accuracy of femoral nerve reinnervation. When applied to the facial or sciatic nerve, nerve-Estim at best had no effect 25 or, worse, impaired the accuracy of reinnervation.23,49 The possible reasons for this discrepancy and mechanisms mediating SC-Estim will be addressed below.
One major difference between the commonly studied rodent femoral and sciatic nerve models of axon regeneration are the target choices. In the femoral nerve, regenerating motor axons are faced with the choice to either regenerate to muscle or cutaneous targets (ie, distal branches and end organs). If a motor axon sprouts exclusively into the cutaneous branch, it has no opportunity to make a functional connection with an end organ target, whereas if it sprouts into both muscle and cutaneous branches, the selective pruning of collaterals that do not reform functional connections appears to be a major mechanism underlying specific reinnervation.4,9,50 In support of this view, both nerve-Estim11-13,36 and conditioning nerve lesions 51 have been demonstrated to simultaneously increase axon sprouting and femoral reinnervation specificity. Furthermore, the extent of reexpression of PSA-NCAM, a known axon sprouting factor, 37 has been demonstrated to play an important role in both robust axon sprouting and specific femoral reinnervation9,11 (Figure 2).
In contrast, regenerating motor axons in the sciatic nerve are faced with the targeting choice of 2 mixed nerve branches, common peroneal (CP) and tibial (TIB), which each supply a mixture of muscle and cutaneous end organs in the lower limb (note: the smaller sural branch has been ignored in most targeting studies). The muscles and corresponding nerve branches originally supplied by CP or TIB MNs appear to be equally supportive of reinnervation regardless of MN origin. 52 Thus, unlike the femoral model, it is unlikely that selective pruning is possible with this type of axon targeting choice.
The assessment of axon targeting in the sciatic nerve model is complicated by the substantial overlap of MN pool topographies between CP and TIB. Using retrograde labeling methods, only a subset of axon targeting errors can be reliably detected in which the topography of reinnervation clearly deviates from the original MN pool. 23 The extent of topographical reinnervation error provides insight into the degree of reinnervation specificity. Using this approach in the sciatic model, treatments that enhance regeneration rate and presumably axonal sprouting such as nerve-ES or chondroitinase ABC have been shown to increase the number of topographical reinnervation errors.23,49 Interestingly, nerve-ES applied to the facial nerve model, which differs from both the femoral and sciatic nerves, in that it is a cranial nerve and has a pure motor distal branch point, also did not show any detectable benefit in topographical reinnervation. 25
Treadmill training regimen, either interval or continuous for 2 weeks, are activity-based interventions that have been shown to augment sciatic reinnervation without compromising its specificity.53-55 Although not yet tested in the femoral model, the treadmill has been hypothesized to augment sciatic nerve reinnervation without decreasing specificity because of its natural pattern of MN activation via the spinal cord locomotor circuitry. 54 In that respect, the MN activation that occurs via our SC-Estim protocol may provide a more natural type of activation paradigm in comparison to nerve-Estim because it depends on intrinsic spinal circuits. If true, then we hypothesized that SC-Estim may have broader applicability than nerve-Estim for enhancing the specificity of reinnervation after peripheral nerve repair because it would not be limited to augmentation of just muscle versus cutaneous targeting choices.
The delay in reinnervation seen 2 weeks after SC-Estim in terms of retrogradely labeled MNs and DRG was an unexpected result (Figures 3A and 3B), especially given that other methods of Estim have generally shown the opposite effect.5,23,24,32 The reason for this discrepancy is not clear. It is highly probable that our SC-Estim protocol has much wider effects on other SC neuronal and nonneuronal cell populations than previous Estim approaches. This is supported by the widespread upregulation of the c-Fos immediate early gene in the lumbar SC following SC-Estim treatment (Figure 1E). So then, why the reduction in the number of MNs whose axons have regenerated into the distal nerve by 2 weeks (Figure 3)? We speculate that the underlying mechanism may be related to some type of alteration in the MN’s central milieu such that the normal posttaxotomy transition from a “transmitting” to a “regenerating” phenotype is delayed. Potential mechanisms to be explored in future studies of SC-Estim should include, but not be limited to, the expression patterns of known regeneration-associated molecule families and their receptors such as the neurotrophins, cytokines, and neurotransmitters and neuropeptides. 56
Despite the smaller number of backlabeled MNs, numbers of myelinated axons were comparable between STIM and SHAM paradigms (Figure 3C). Taken together with the aforementioned reductions in MN and DRG numbers at 2 weeks (Figures 3A and 3B), this implies that SC-Estim increased the number of collaterals produced by each parent axon that crossed the repair site within the first 2 weeks posttransection, which may in itself play a critical role in the enhancement of reinnervation specificity. This is consistent with the relatively greater upregulation of PSA-NCAM after SC-Estim (Figure 2), which is a molecule shown to facilitate axon sprouting and targeting in both neural development42-45 and regeneration.9,11
Femoral nerve axons regenerate in discrete waves in a process known as staggered reinnervation. The initial phase involves a subset of “pioneer” axons that reinnervate the nerve early but without specificity. 4 However, the subsequent phase of axons appear to reinnervate with remarkable precision through some combination of selective pruning (detailed above) and specific axon outgrowth, which are likely governed by the distinct neurotrophic factor profiles of each distal pathway and its respective end organs upregulated after denervation. 57 We hypothesize that by having fewer neurons but no reduction in the number of pioneer axons participating in the nonspecific early phase of reinnervation, SC-Estim improves specificity by increasing the proportion of neurons participating in the highly specific late phase of reinnervation during which they likely have access to a more elaborate set of guidance factors to direct their outgrowth and/or pruning.
In summary, our results support the continued development of SC-Estim as a potential therapy for improving functional outcomes after PNI. The improvement in reinnervation specificity with SC-Estim appears to involve at least 1 established guidance factor for regenerating axons—PSA-NCAM; however, there is little doubt that molecular mechanisms underlying this phenomenon are much more complex. A greater understanding of these mechanisms should yield increasingly more refined treatment options to manage the tremendous burden posed by PNI. An intriguing feature and potential advantage of SC-Estim over nerve-Estim is that it uses intrinsic spinal circuitry to activate MNs, which is a less specific, but perhaps a more natural mechanism of activation. Future studies aimed at correlating functional outcomes with these results as well as testing the applicability of SC-Estim to other nerve injury models will be of great interest. In addition, the present data provide proof of principle for the exploration of other central nervous system–directed approaches to lower MN activation, such as transcranial magnetic stimulation, which would increase the potential for clinical application of this phenomenon.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant to DWZ and RM from the Canadian Institute for Health Research (Regenerative Medicine and Nanomedicine Team Grant #163322). Postdoctoral fellowship support to CKF was provided by Alberta Innovates-Health Solutions (AI-HS), whereas BS was supported by a graduate student award from AI-HS.