
Abstract
Sudden unexpected death in epilepsy (SUDEP) is the most catastrophic complication associated with epileptic seizures. Despite its devastating impact, the mechanistic underpinnings of SUDEP remain poorly understood, limiting the development of effective preventive strategies. Animal models have proven indispensable in understanding the complex pathophysiology of SUDEP, offering controlled systems to dissect potential causes of these fatal events. This review provides a comprehensive overview of the current landscape of SUDEP animal models, with an emphasis on key experimental variables that impact SUDEP outcome in animal studies. In particular, this review focuses on how the animals’ genetic background, age, sex, vigilance state, and time of day influence their phenotypic variability. We also report the major discrepancies in terminal seizure characteristics between mice and humans and highlight the necessity to segregate “anti-SUDEP” effects from potential “antiseizure” effects. From a regulatory and animal welfare perspective, we discuss the challenges of studying SUDEP as an endpoint and outline the critical steps to implement the preclinical SUDEP common data element, thereby promoting consistency, reproducibility, and interoperability across preclinical studies. By synthesizing insights across critical experimental variables and remaining challenges in SUDEP preclinical studies, this review aims to promote rigorous experimental design in animal SUDEP studies, with the long-term goal of facilitating translational insights into human SUDEP prevention.
Introduction
Sudden unexpected death in epilepsy (SUDEP) is the primary cause of death in epileptic individuals. Animal models of SUDEP offer a complementary tool with rich resources for testing and validation that is not possible in humans. Death occurring immediately after a generalized convulsive seizure in an otherwise healthy animal is considered clinically relevant to the study of SUDEP. SUDEP-like events have been documented in a wide range of model organisms (Figure 1). Albeit evidence of SUDEP across diverse species, the house mouse (Mus musculus) has been the primary model for studying SUDEP, accounting for >70% of the published literature that studies SUDEP using animals (Figure 1). This review examines key experimental variables in mouse models of SUDEP that impact the outcome measures. It also addresses the phenotypic differences in terminal seizure characteristics between animal models and human SUDEP cases. We emphasize the critical need to distinguish “anti-SUDEP” from potential “antiseizure” effects. The goal of this review is to promote rigorous experimental design and enhance reproducibility in SUDEP research using animal models.

Common model organisms used in SUDEP studies. Proportions were estimated by querying PubMed for “SUDEP” and each species name during 2015 and 2025. SUDEP: sudden unexpected death in epilepsy.
Seizure Induction Paradigms and SUDEP Rates
Experimentally confirmed seizures, either spontaneous or induced, are a prerequisite for the study of SUDEP. Spontaneous seizures are typically infrequent, making sequential SUDEP events extremely rare in mice, except in the presence of specific pathogenic mutations. Moreover, reliable detection of spontaneous seizures requires continuous long-term video-electroencephalogram (EEG) recordings, which are highly time- and resource-intensive. Although spontaneous seizures are a preferred metric, 1 researchers more commonly use induced seizure models in SUDEP studies because they allow greater feasibility and more precise control of the timing of seizures. SUDEP has been reported following seizures induced through diverse mechanisms, including chemoconvulsants, acoustic stimuli, hyperthermia, and maximal electroshock (MES). However, caution is warranted when interpreting mortality in these models, as there is the possibility of competing causes of death unrelated to epileptic activities, particularly heatstroke or dehydration in hyperthermia-induced seizure, or direct cardiorespiratory arrest following MES when high alternating currents are applied. It is also important to note that mortality following status epilepticus does not meet the definition of SUDEP. Therefore, prolonged seizures induced by pilocarpine, kainic acid, or other approaches are not appropriate models of SUDEP.
SUDEP is commonly assessed using either the mortality rate as a binary readout or the survival curve over time as right-censored data. A high mortality rate in response to the first seizure stimulus (eg, ∼100% SUDEP in audiogenic seizure in primed DBA/2J mice) is a desirable feature for efficient testing of potential rescue strategies in SUDEP animal models. However, models in which animals experience multiple nonfatal seizures prior to a fatal episode offer distinct advantages. These within-subject designs allow for critical comparisons that help elucidate why certain seizures are survivable while others become fatal. Such models enable investigation into the “secondary threshold” that must be crossed for a seizure to result in SUDEP in a given individual. Additionally, these models enhance statistical power, reduce animal usage, and provide a more robust framework for testing SUDEP prediction algorithms and mechanistic hypotheses.
Critical Experimental Variables for Studying SUDEP in Animals
When designing animal studies of SUDEP, critical experimental variables, including strain genetic background, sex, and age of mice, as well as time of testing, should be carefully considered for the expected SUDEP phenotype and also for experimental repeatability and reproducibility.
Strain Genetic Background
Each human individual, including monozygotic twins, is genetically unique. This different genetic makeup largely determines our susceptibility to disease and response to medication. Likewise, in preclinical epilepsy and SUDEP studies, the genetic background of inbred mouse and rat strains is a pivotal experimental variable, as seizure susceptibility and SUDEP phenotypes are strongly influenced by strain-specific genetic factors.
Mouse models, such as audiogenic seizures in DBA/1J and DBA/2J mice, have been widely used in SUDEP research due to their high mortality rates, particularly after priming at a young age. However, mortality in response to acoustic stimulation varies significantly across strains, emphasizing the importance of genetic background-dependent phenotypes. Martin et al found that a 129/SvTer mouse is a more versatile model of SUDEP due to its wider age range of susceptibility compared to DBA/1J and DBA/2J mice without priming. 2 The strain-dependent SUDEP phenotype was also observed following seizures induced by MES, 3 cerebral malaria infection, 4 and flurothyl exposures. 5 In our screening of 31 strains of recombinant inbred Collaborative Cross mice, we observed striking variability in SUDEP sensitivity across mouse strains following flurothyl-induced seizures. 6 Notably, these differences were independent of their seizure susceptibility, underscoring how naturally occurring genetic variation can influence SUDEP risks. Strain genetic background-dependent SUDEP susceptibility is also frequently observed in SUDEP mouse models carrying particular pathogenic mutations. The best example is the Dravet syndrome model mice. Scn1a+/− mice on the C57BL/6J background develop spontaneous seizures and die within weeks. In contrast, Scn1a+/− mice on the 129S6/SvEvTac background do not exhibit an overt phenotype. 7 Of note, even the choice of different substrains can significantly alter SUDEP phenotypes in Scn1a+/−,8,9 Scn8a gain-of-function mutant, 10 and Kcna1 knockout11,12 mice. These findings underscore the importance of considering and explicitly reporting mouse strain and substrain backgrounds in SUDEP animal studies.
While often viewed as a confounding variable, the naturally occurring genetic diversity of different mouse strains can be leveraged as a powerful tool to identify loci/genes and modifiers that control SUDEP susceptibility/resistance through targeted breeding and quantitative trait locus mapping (Figure 2). Among other candidates, genetic mappings revealed Gabra2 as a common genetic modifier of SUDEP phenotype for both Scn1a 13 and Scn8a 10 mutations given that C57BL/6J, one of the shared parental strains for both studies, carries a splice site mutation that reduces the expression of Gabra2. 14
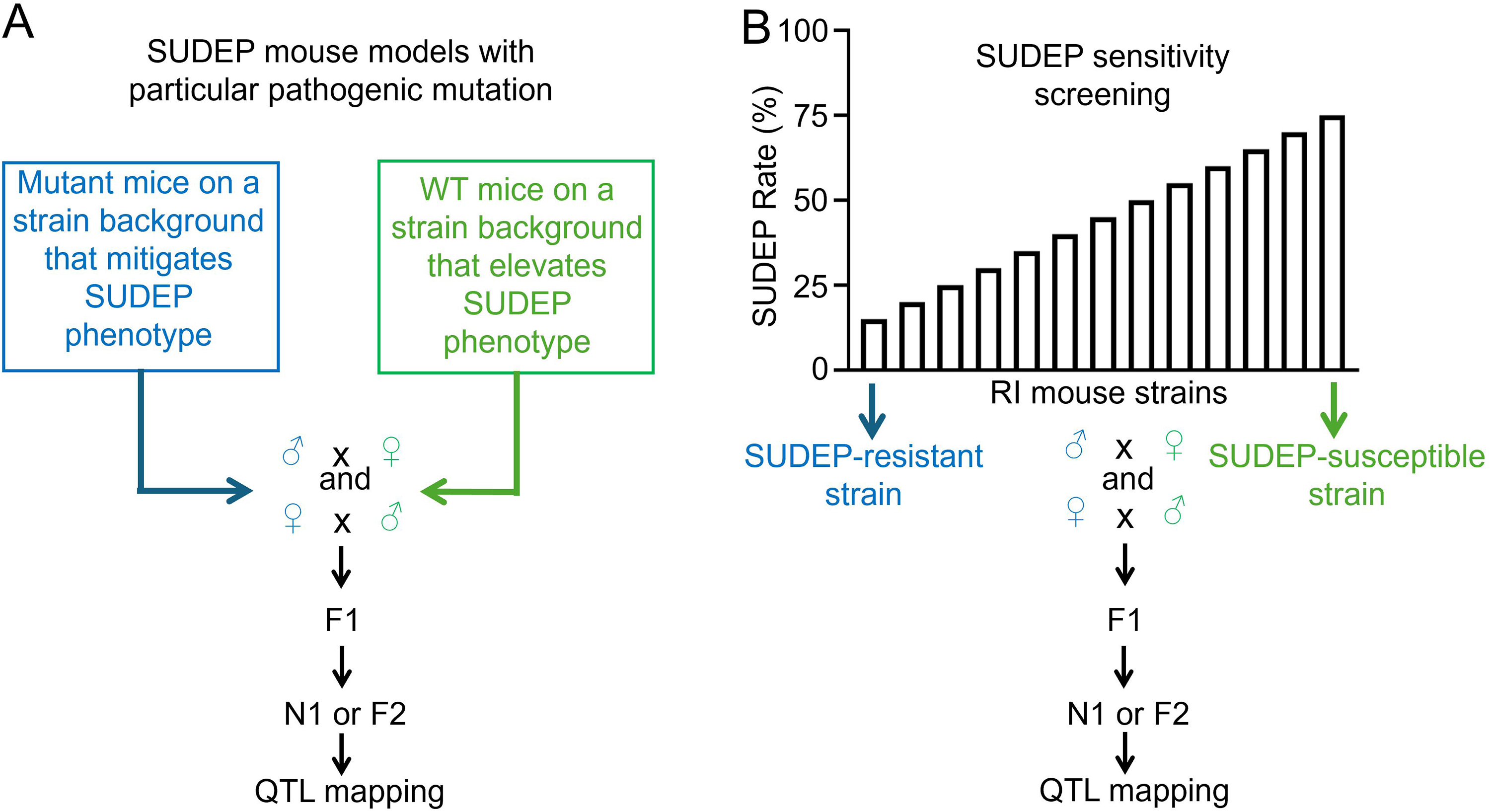
Genetic mapping strategies using (A) SUDEP model mice with specific pathogenic mutations and (B) a population of recombinant inbred (RI) mice to identify genetic modifiers and candidate genes that control the trait of SUDEP. Depending on the QTL effects based on F1 phenotyping, N1 (backcross) or F2 (intercross) population can be generated for QTL mapping. SUDEP: sudden unexpected death in epilepsy; QTL: quantitative trait locus.
Age and Sex
Age plays a significant role in deciding an individual's susceptibility to SUDEP. Unlike the inverted “bell-shaped” curve of the epilepsy prevalence with age in the general population, which peaks in childhood and late adulthood, SUDEP risk is highest among young adults aged 20 to 45 years 15 (Figure 3). The reason for this inverse relationship between the prevalence of epilepsy and SUDEP remains unclear, but the observed SUDEP rates in young adults are significantly higher than expected. One possible explanation is that young adults are more likely to live alone, whereas children and the elderly are more often under the care of others, potentially leaving young adults more vulnerable to fatal outcomes following a seizure due to the lack of supervision or immediate intervention. On the other hand, animal models employing induced seizures at various ages recapitulate the overall age dependency of SUDEP in humans, arguing against an effect driven by caregiver presence, assuming lab animals receive consistent veterinary care throughout their lifespan. One study induced audiogenic seizures in mice that carry the pathogenic Scn8AN1768D/+ mutation at various postnatal ages (P15, P20-P21, P32, and P49-P69). Remarkably, 11 out of 13 mice at P20 to P21 succumbed to SUDEP after audiogenic seizure. In contrast, no SUDEP-related mortality was observed in the younger (P15) or older (P32 and P49-P69) age groups. 16 Other audiogenic SUDEP mouse models, like DBA/2J, showed similar trends where the SUDEP rate was highest at P21 and declined as the mice aged. 2 Longitudinal monitoring of survival curves in multiple mouse models carrying SUDEP-related pathogenic mutations further corroborates the age-dependent nature of the SUDEP phenotype. SUDEP most frequently occurs in juvenile mice carrying pathogenic mutations of SUDEP-associated genes, including Scn1a+/− (∼P23-P27), 17 Scn2aR1882Q/+ (∼P12-P29), 18 Scn1b homozygous KO (∼P12-P24), 19 Kcna1 homozygous KO (∼P21-P35), 12 and Depdc5 dorsal cortical progenitor (Emx1-Cre) KO (∼P20-P25) 20 mice. While the peak SUDEP age is delayed until young adulthood in Scn8AN1768D/+ (∼P60-P120), 21 Depdc5 pan-neuronal (Syn1-Cre) KO (∼P90-P140) 22 and forebrain excitatory neuronal (Rbp4-Cre) KO (∼P50-P120) 23 mice. Notably, the mice carrying the identical pathogenic gene exhibit distinct survival curves, depending on specific mutations and/or affected cell populations.24,25 Overall, the animal studies support the observation that SUDEP occurs at significantly higher rates during juvenile to young adulthood, generally P21 to P60 in mice, compared to earlier or later life stages. This pattern is consistent with SUDEP incidence trends seen in humans.

Estimated prevalence trajectory of epilepsy in the general population and prevalence trajectory of SUDEP in epilepsy patients across ages. SUDEP: sudden unexpected death in epilepsy.
Sex, like age, influences an individual's risk of SUDEP. In humans, males are associated with a 1.4-fold increase in SUDEP risk and generally constitute more than 60% of all SUDEP-related mortalities in a given population. 26 The underlying mechanism for this sex difference is unclear, but studies show that the occurrence of epileptic seizures is generally higher in men than in women and hormonal milieu contributes to variations in epilepsy presentation between men and women. Consistent with clinical observations, male Scn8AN1768D/+ mice and Kcna1 homozygous KO mice have a higher SUDEP-associated mortality rate during early development compared to female littermates.27,28 In contrast, female mice carrying Scn1a+/− exhibit a significantly higher mortality rate compared to their male counterparts, though male mice have more frequent and severe seizures. 29 In a separate study using the same line of Scn1a+/− mice, however, no sex differences (P = 0.933) were observed in the SUDEP occurrences, despite the increased seizure frequency and severity in female mice. 30 Overall, these findings highlight substantial disagreement in the literature—not only regarding which sex is more susceptible to SUDEP, but also concerning the underlying mechanisms driving that susceptibility.
Vigilance State and Time of the Day
A significant majority (∼58%-88%) of human SUDEP cases occur during the night and early morning while the patients are presumably asleep. 31 However, this elevated SUDEP risk can be attributed to multiple factors, such as time of day, vigilance state, sleep homeostasis, and circadian clock. Gordon Buchanan's team conducted a series of animal studies to unravel the causal elements of SUDEP nocturnality. Their first study examined how a mouse's vigilance state impacts its susceptibility to SUDEP. Seizures induced via MES during rapid eye movement (REM) sleep were associated with more profound respiratory suppression and were more likely to result in death compared to equivalent seizures induced when the mice were awake or under non-rapid eye movement sleep. 32 Indeed, REM sleep represents a small portion of total sleep and is generally resistant to spontaneous seizures; however, when seizures were experimentally induced during REM sleep in this model, they consistently resulted in death. The impact of diurnality on SUDEP sensitivity was further studied under a 12/12 light-dark cycle by challenging different cohorts of mice at various times of the day using audiogenic seizure in DBA/1J mice. The SUDEP rate was highest during dusk and dawn, with nearly 90% mortality around early morning (∼ZT 22 h) versus zero mortality at noon (∼ZT 6 h). The same trend persisted even when the mice were raised in constant darkness. 33 A similar diurnal SUDEP pattern was also observed following spontaneous seizures in Scn1aR1407X/+ transgenic mouse model of Dravet syndrome. 34 The elevated mortality during the nighttime did not result from increased seizure frequency, as the number of seizures were comparable between subjective nighttime and daytime. 34 Altogether, these findings suggest that nighttime represents a risk factor for SUDEP in both humans (diurnal) and mice (nocturnal), possibly through different mechanisms, despite their opposite activity patterns.
Major Discrepancies in Terminal Seizure Characteristics Between Mice and Humans
While mouse models of SUDEP are extremely valuable for elucidating underlying mechanisms and developing therapeutic interventions, major discrepancies exist between terminal seizure characteristics in mice and humans.
First, the semiology of terminal seizures differs dramatically across species. In humans, generalized tonic-clonic seizures (GTCSs) are a major risk factor for SUDEP and precede the majority of the limited number (n = 11) of monitored cases. 35 Patients experiencing GTCSs typically first show sudden muscle stiffness followed by rhythmic jerking of the extremities. Conversely, in mice, terminal seizures—except those induced by MES—typically begin with a prominent clonic phase, followed by a transient tonic extension of both forelimbs and hindlimbs before SUDEP 36 (Figure 4). This distinct terminal seizure semiology may reflect differences in how terminal seizures originate and propagate in the human brain versus the mouse brain. For example, studies support the hypothesis that propagation of seizures to the brainstem, where stimulation can directly result in tonic muscular activities, 37 is heavily involved in terminal seizures induced by electroshock, 38 audiogenic, 39 and flurothyl 40 in rodents. Moreover, the tonic contraction of the diaphragm is sufficient to produce the concomitant apnea independent of forebrain epileptiform activity, which may lead to SUDEP in these animals. 41

Schematic of major discrepancies in terminal seizure characteristics between humans (based on limited observations) and mice.
Second, the temporal dynamics of the postictal period are highly variable across species. Data from the MORTEMUS study, which monitored a limited number of patients preceding SUDEP, indicate that the postictal phase is heterogeneous and often prolonged, ranging from 3 to 18 min. 35 Remarkably, the postictal period in these patients often features a progressive breakdown in central autonomic control, characterized by impaired respiratory and cardiac function, generalized EEG suppression, and ultimately, terminal asystole. 35 In contrast, mice typically exhibit ictal cardiorespiratory impairment and a brief and uniform postictal phase with tonic limb extension rapidly transitions to relaxation of muscle tone and cessation of breathing and brain activity, usually lasting less than 10 s (Figure 4). Interestingly, some studies have reported residual bradycardia persisting (for 10-20 min) even after brain and respiratory activity have ceased following spontaneous, audiogenic, and pentylenetetrazole-induced seizures in mice carrying gain-of-function Scn8a mutations, 36 MES-induced seizure in Lmx1bf/f/p mice, 42 spontaneous seizure in Kcnh2 knock-in rabbits, 43 as well as in our own studies following flurothyl-induced seizures in pro-SUDEP Collaborative Cross mice. 5 These findings raise the possibility of decoupled postictal cardiac-autonomic responses and suggest additional complexity in SUDEP pathophysiology that may not be fully captured by the standard mouse seizure-to-death trajectory.
Anti-SUDEP Versus Antiseizure
Theoretically, seizure control remains the primary strategy for SUDEP prevention. However, SUDEP commonly occurs in patients with drug-refractory epilepsy, where the incidence rises significantly, from 1.5 to 4.2 per 1000 patient-years in this population. 44 Given the persistent challenges in achieving optimal seizure control for these patients, anti-SUDEP therapy that works independently of seizure control is urgently needed. This presents a major challenge for preclinical studies, as most existing SUDEP rescue studies delivered genetic, pharmacologic, and/or neuromodulation (eg, electrical, chemogenetic, and optogenetic) therapeutic interventions before seizure onset, thereby inevitably affecting seizure activity itself. Consequently, any observed potential anti-SUDEP effects may be confounded by unintended antiseizure actions of the intervention, making it difficult to discern whether reduced mortality results from true SUDEP mitigation or merely improved seizure control.
To address these concerns, many SUDEP studies attempt to characterize the frequency, threshold, severity, and duration of the “terminal” seizures to demonstrate that the intervention prevents SUDEP without directly altering seizure per se. Nevertheless, we need to take extra caution when interpreting these mortality results. For example, enhancing serotonergic (5-HT) tone before seizure induction has been shown to reduce postictal apnea and SUDEP. 45 However, growing evidence indeed suggests that 5-HT augmentation also has antiseizure effects. One study demonstrated that photo-stimulation of dorsal raphe nucleus 5-HT neurons before seizure induction suppressed audiogenic seizures in the DBA/1J mouse model of SUDEP, raising the possibility that a decrease in frequency of seizure-induced respiratory arrest may be secondary to a lower seizure severity. 46 An optimal experimental design to isolate true anti-SUDEP effects would require more precise temporal control with life-saving interventions initiated after seizure termination. Such designs would avoid direct modulation of seizure activity and more clearly delineate the specific effects on SUDEP prevention. Nonetheless, all SUDEP prevention strategies, whether their mechanism of action is antiseizure or not, are essential to saving lives from this tragic consequence.
SUDEP as an Endpoint for Preclinical Studies and Implementation of Common Data Elements
Preclinical SUDEP studies inevitably involve animal models in which death is an anticipated outcome, requiring more rigorous ethical and regulatory oversight. Unlike chronic disease models—such as those for cancer, where body weight loss is commonly used as a humane endpoint—SUDEP occurs suddenly and unexpectedly in otherwise healthy epileptic animals. Growing evidence indicates that seizure-induced respiratory arrest is the primary cause of SUDEP in various animal models. Interventions such as mechanical ventilation 36 and diaphragmatic pacing 47 have been shown to resuscitate affected animals. As such, ictal and periictal respiratory arrest may serve as a potential humane endpoint in SUDEP research. However, there is still a lack of institutional guidelines regarding humane endpoints specific to SUDEP studies. As a result, basic scientists often find SUDEP-focused protocols particularly challenging to design and justify, especially when seeking approval for experiments that anticipate death without preceding signs. These regulatory complexities may discourage investigators from pursuing SUDEP research. A recent report from the Epilepsy Research Institute in the United Kingdom noted that some researchers may redirect their focus away from SUDEP to other areas of epilepsy research with fewer regulatory obstacles. 48
Preclinical common data elements (CDEs) are standardized data definitions designed to promote consistency, reproducibility, and interoperability across preclinical studies. Their use enhances data sharing and integration, which are critical for advancing big data analytics. Citizens United for Research in Epilepsy assembled a working group to develop SUDEP-specific preclinical CDEs. These CDEs span several topic areas, including neurological and physiologic variables, therapeutics and pharmacology, imaging, electrophysiology, and comorbidities.49,50 Moving forward, successful implementation will depend on raising awareness, encouraging adoption, and ensuring that SUDEP research findings are routinely reported using these CDEs. Accumulation of standardized preclinical data will ultimately enable the aggregation of high-quality datasets, paving the way for advanced big data approaches and facilitating artificial intelligence-aided SUDEP analysis and prediction.
Concluding Remarks
While animal studies have significantly advanced our understanding of SUDEP, knowledge gaps remain, particularly regarding experimental variables such as genetic background, sex, age, and vigilance state, which can impact SUDEP phenotypes in animal models. These variables complicate data interpretation and also provide opportunities to identify risk factors through appropriately designed experiments. Special attention also needs to be paid when extrapolating findings from animal studies to humans, given the distinct characteristics of terminal seizures between species and the need to separate “anti-SUDEP” from “antiseizure” effects. Adoption of alternative endpoints, such as seizure-induced respiratory arrest, can improve animal welfare and mitigate the regulatory obstacles. A final consideration is that implementing SUDEP preclinical CDEs can enhance experimental repeatability and reproducibility.
Footnotes
Acknowledgment
The authors thank Tyler Shannon for his participation in conceptualizing and discussing this article.
Author Contributions
MN and YS contributed equally to this article. MN, YS, and BG participated in the literature search, drafting, and revising the manuscript. BG supervised the project. All authors reviewed and approved the final manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Institute of Neurological Disorders and Stroke (Grant No. 1R01NS140338-01A1).