
Abstract
In response to several publicized failures of inactivation procedures, the Select Agent program in the United States has recently focused on validation or documentation of inactivation methods appropriate for biological select agents and toxins covered under the regulations. Although some data on inactivation of Brucella spp are available within the historical literature, reports are primarily limited to heat inactivation of milk, and there are discrepancies among studies in time required to eliminate all viable bacteria. In our experiment, complete elimination of viable Brucella bacteria (B abortus, B suis, and B melitensis) within 30 to 60 minutes required temperatures approaching boiling, whereas lower temperatures required much longer heating times (hours). Buffered neutral formalin (10% concentration) was highly effective in inactivating Brucella bacteria by 4 hours from tissue sections that had high levels of colonization. Methods based on methanol or methanol-acetone at 50% to 70% concentrations inactivated viable Brucella from liquid samples within 3 to 5 days. Phenolic disinfectants were also highly effective in quickly inactivating Brucella in solutions. After passage through a 0.22-μm filter, no Brucella were recovered from spiked serum samples or serum samples from experimentally infected animals. Despite demonstration of the effectiveness of these procedures in our laboratory, it is highly recommended that others validate any similar procedures used under their own in vitro conditions. It is also recommended that culture methods be used, as possible, on all inactivated samples to verify the effectiveness of the validated procedure and protect against inadvertent laboratory errors.
Partly due to historical efforts to weaponize Brucella, such as experiments by the US military with Brucella suis in the 1950s, bacteria in this genus have been considered to be potential bioweapons. Following congressional passage of the Antiterrorism and Effective Death Penalty Act of 1996 and publication in the Federal Register on October 24, 1996, regulatory oversights were put in place for the transfer of biological agents and toxins (BSATs; ie, Select Agents) that have the potential to pose a severe threat to public health and safety. Subsequently, the Bioterrorism Act of 2002 required consideration of the following for inclusion of an agent or toxin on the select agent list: the degree of pathogenicity (ability of an organism to cause disease), communicability (ability to spread from infected to susceptible hosts), ease of dissemination, route of exposure, environmental stability, ease of production, ability to genetically manipulate or alter, long-term health effects, acute morbidity (illness), acute mortality (death), available treatment, status of host immunity, vulnerability of special populations, and the burden or impact on the health care system. As a result of their potential for use in bioterrorism, B abortus, B suis, and B melitensis were included on the original select agent list and have remained listed as BSATs after biennial reviews as required by the Agricultural Bioterrorism Protection Act of 2002.
For various reasons, there is frequently a need to inactivate cultures or samples from animals infected with BSATs to allow movement outside approved select agent areas or containment facilities. In response to several publicized failures of inactivation procedures, most notably related to Bacillus anthracis samples, the Select Agent program has recently focused on validation or documentation of inactivation methods appropriate for BSATs. On March 31, 2017, published guidance on established regulatory requirements for inactivation processes was distributed. Although some data are available within the historical literature, reports are primarily limited to heat inactivation of milk, and there are discrepancies among studies in times required to inactivate Brucella. Therefore, this article is designed to provide data validating some methods for inactivating laboratory and sera samples containing high concentrations of B abortus, B suis, and B melitensis. Although publication of these data is not meant to be inclusive of all possible inactivation processes, this information should be beneficial in preventing duplicate validation studies by scientists working with Brucella spp at other registered select agent laboratories by providing baseline data to validate procedures used under their own in vitro conditions.
Methods
Bacterial Cultures
Cultures of B melitensis strain 16M, B abortus strain RB51, B suis strain 1330, and B abortus strain 2308 were obtained from the National Animal Disease culture collection. Cultures were first propagated on tryptose agar (Difco Laboratories, Detroit, Michigan) containing 5% bovine sera (tryptic soy agar [TSA]) for 72 hours at 37°C and 5% CO2. Bacteria were harvested from agar plates with 0.15M phosphate-buffered saline (PBS) and the concentration adjusted for each inactivation study with an optical density method and a spectrophotometer. Final concentrations of live bacteria in suspensions used for inactivation studies were determined by serial dilution and standard plate counts on TSA plates and reported as colony-forming units (CFU) per milliliter.
Filter Sterilization
Sterile PBS and bison sera were spiked with B abortus strain 2308 at concentrations of approximately 103 and 109 CFU/mL. Two milliliters of spiked sera or spiked PBS were filtered once through a 0.22-μm filter (Millex-GV; Millipore-Sigma, Billerica, Massachusetts). A portion of each filtrate (150 μL) was plated in duplicate onto Kuzda and Morse (KM) plates 1 and incubated at 37°C and 5% CO2 for 7 days. The remaining filtrate for each sample (approximately 1.7 mL) was added to 5 mL of tryptose broth. A similar volume of unfiltered PBS and bison sera that had been spiked with bacteria was added to 5 mL of tryptose broth. After 7 days of incubation at 37°C and 5% CO2, 150 μL from each tryptose broth sample was plated in quadruplicate on KM media and incubated for 7 days. Viable bacteria were quantified with standard plate counts for each sample. Any recovered isolates with colony morphology compatible with Brucella were tested with a Brucella-specific polymerase chain reaction (PCR) assay.1,2
Greater than 7000 sera samples of variable volumes (1 to >50 mL) obtained at various times after experimental infection of cattle, bison, and elk with B abortus, as well as cattle and swine infected with B suis and goats infected with B melitensis, were filtered through 0.22-μm filters. These samples included sera samples from animals obtained within 24 to 48 hours after brucellosis abortion. Data from our studies suggest that these blood samples are frequently culture positive for recovery of Brucella. After filtration, 100 to 200 μL of each sample was plated on KM media and incubated at 37°C and 5% CO2 for 7 days. Plates were examined at 7 days for Brucella colonies. As with filter sterilization, a Brucella-specific PCR assay was used to test any colonies that had morphology similar to Brucella spp.
Heat Inactivation
Cultures of 4 Brucella species (2308, RB51, 1330, 16M) were cultured as described and washed from plates with PBS. After adjustment to approximately 5 × 109 CFU/mL based on optical density, 5 mL of the bacterial solution was placed into glass tubes (13 × 100 mm). Glass tubes were placed into a dry heating block (Barntead Thermolyne, Dubuque, Iowa) that had been adjusted to temperatures of 65°C, 75°C, 85°C, or 95°C based on a thermometer located in the center of the block. Concentration of bacteria in the initial inoculum for each temperature was determined by serial dilution prior to heat treatment. At 60-minute intervals for heat block temperatures of 65°C and 75°C, 30-minute intervals for 85°C, and 15-minute intervals for 95°C, samples were obtained from each sample for microbiological evaluation. All samples were serially diluted, plated in duplicate onto KM plates, and incubated at 37°C in 5% CO2 for 7 days. Concentration of bacteria within each sample was determined by standard plate counts. Experiments were run in triplicate for each Brucella spp at each temperature. As with filter sterilization, a Brucella-specific PCR assay was used to confirm any isolates recovered after heat treatment that had colony morphology consistent with Brucella spp.
To model the time needed to heat tubes of bacteria to water bath temperature, a glass tube (13 × 100 mm) containing 5 mL of PBS at room temperature was placed into the heat block at each temperature. An alcohol thermometer was suspended in the center of the PBS without touching the bottom or sides of the glass tube. After placement into the heat block, temperature readings were obtained at approximately 2-minute intervals at each temperature until the temperature of the PBS remained stable for a 4-minute interval. Three replicates of this experiment were conducted.
Formalin Inactivation
To validate the inactivation properties of formalin, samples of cotyledon were removed at necropsy from 6 bison that aborted after experimental infection in midgestation with strain 2308. After being trimmed to approximately 1.5-cm thickness, fresh sections were placed into whirlpack sample bags, and additional sections were immersed in 10% buffered neutral formalin at approximately a 1:10 ratio (vol:vol, tissue:formalin). At approximately 2, 4, and 24 hours after being placed in formalin, 1 to 2 g of cotyledon was removed from formalin and washed once in 50 mL of sterile PBS. Approximately 1 g of formalin-fixed samples and fresh, unfixed cotyledon were obtained, weighed, and individually ground in 2 mL of PBS (pH = 7.2) with sterile glass grinders in a similar manner to methods used in other studies.3,4 Serial dilutions of each tissue suspension were plated onto KM plates and incubated at 37°C in 5% CO2 for 7 days. Standard plate counts were used to determine concentration of live bacteria (CFU/g) in fresh and formalin-fixed samples.
Methanol-Acetone and Methanol Inactivation
Cultures of 2308, 16M, and 1330 were grown as previously described and washed from plates with PBS, with bacteria ultimately suspended in 10-mL volumes of 0%, 40%, 50%, 60%, or 70% methanol-acetone (50:50 vol:vol) at approximately 1010 CFU/mL. All tubes were incubated at 4°C with 100-µL aliquots removed from samples after agitation at 3 and 6 days and plated on KM plates. In addition, 2 mL of each sample was obtained at each time point, centrifuged for 2 min at 16 000 × g, washed once in 1 mL of PBS, and suspended in 1 mL of PBS, with 200 µL plated on KM plates. Plates were incubated at 37°C in 5% CO2 for 7 days and standard plate counts completed. All isolates with colony morphology consistent with Brucella spp were confirmed with a Brucella-specific PCR assay.1,2 Two replicates of the study were conducted.
To assess inactivation with methanol, 2308, 16M, RB51, or 1330 bacteria were grown and harvested as previously described and then suspended in a total volume of 5 mL at methanol concentrations of 30%, 40%, 50%, 60%, or 70% with bacterial concentrations at approximately 1010 CFU/mL. Additional polystyrene tubes contained the bacterial stock solution prepared in 5-mL volumes without methanol (0%). Tubes were incubated at 4°C and 1-mL aliquots removed from each tube after 3, 5, and 7 days. After centrifugation and 1 PBS wash, the pellet was suspended in 100 µL of PBS and plated in duplicate onto TSA plates. Bacterial growth was evaluated after incubation at 37°C and 5% CO2 for 7 days and colonies with morphology consistent with Brucella confirmed by PCR. In a similar manner, a second replication evaluated 2308, 16M, and 1330 stocks in methanol suspensions of 40%, 50%, 60%, and 70% at bacterial concentrations of approximately 1010 CFU/mL.
Inactivation by Disinfectants
Cultures of 2308, 16M, and 1330 were prepared as previously described and adjusted to approximately 1010 CFU/mL with optical density measurements. After addition of 200 μL of bacterial stock to a polystyrene tube, an equal volume of saline or disinfectants prepared in accordance with manufacturer recommendations (Amphyl, 0.5% vol/vol; Wexcide, 1 oz per gallon) were added and the suspension vortexed. After incubation at room temperature for 10, 20, or 30 minutes, 1 mL of PBS was added to each tube. Tubes were then centrifuged at 16 000 × g for 2 min, washed once with PBS, and suspended in 1 mL of PBS. After inoculation of 100 µL onto TSA plates in duplicate, cultures were incubated at 37°C in 5% CO2 for 7 days. Colonies with morphology consistent with Brucella were confirmed by PCR
To evaluate the effect of dilution on bactericidal activities of disinfectants, 10-1 and 10-2 dilutions of working concentrations of Amphyl and Wexcide were also prepared. Equal volumes (200 μL) of 1010 CFU/mL of 2308, 16M, and 1330 suspensions and diluted disinfectants were combined and incubated at room temperature for 10, 20, or 30 minutes. After addition of 1 mL of PBS, tubes were centrifuged, washed once with PBS, and suspended in 1 mL of PBS, with 100 µL plated in duplicate on TSA plates. Bacterial growth was evaluated after incubation at 37°C and 5% CO2 for 7 days and any suspect colonies confirmed by PCR.
Statistics
Microbiologic data for heat and formalin inactivation experiments were converted to a logarithm for analysis, with samples in which no recovery was made converted to 1 prior to logarithmic conversion. Data were compared by a general linear model procedure (SAS Institute Inc, Cary, North Carolina) to evaluate the effects of strain and/or time. Means were separated by a least square means procedure (P < .05).
Results
Filter Sterilization
Methods used in the filter sterilization experiment should have detected live Brucella at concentrations >3 CFU/mL in spiked samples prior to enrichment. Spiked PBS and bison sera samples that were not passed through a 0.22-μm filter had a lawn of Brucella growth on media after 7 days of incubation at 37°C. Tryptose broth inoculated with unfiltered spiked samples and enriched at 37°C and 5% CO2 for 7 days also had a lawn of Brucella growth after inoculation on media. In contrast, no Brucella isolates were recovered from spiked serum samples plated immediately after filtration once with a 0.22-μm filter or after 7 days of enrichment in tryptose broth.
Our estimated level of detection was between 5 and 10 CFU/mL from sera from experimentally infected animals. No Brucella isolates were recovered after inoculation of media with filtered serum samples obtained from animals (cattle, bison, elk, swine, goats) at various times after experimental infection with Brucella spp (B abortus, B suis, or B melitensis), including samples obtained from culture-positive animals shortly after abortion.
Heat Inactivation
Our data indicated that time had a significant influence (P < .001) at all temperatures and that recovery of viable bacteria was influenced (P < .05) by Brucella strain during heat killing at 65°C, 75°C, and 95°C temperatures but not at 85°C (Figure 1). Internal temperatures within the tubes stabilized at 61°C, 69°C, 79°C, and 90°C in approximately 10 to 12 minutes for heat block temperatures of 65°C, 75°C, 85°C, and 95°C, respectively (Figure 1).
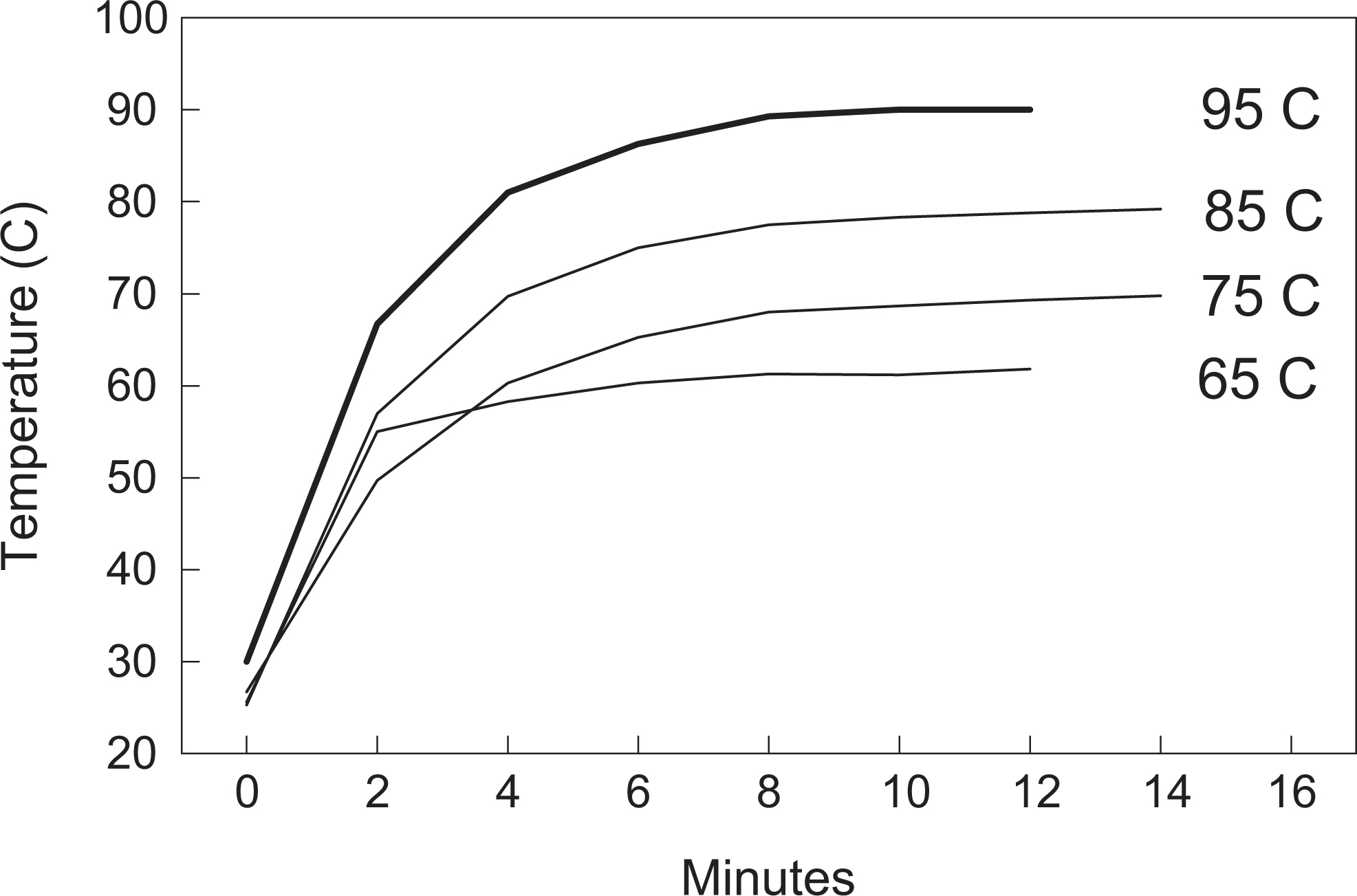
Temporal change in internal temperature within 5 mL of phosphate-buffered saline in glass tubes after placement on a heat block set at temperatures of 65°C, 75°C, 85°C, and 95°C.
When heated in a heat block set at 65°C, Brucella strains showed an average reduction in CFU/mL of 3.6 logs across Brucella strains by 60 minutes, but rate of inactivation plateaued such that a mean of 3.7 ± 0.18 logs of viable bacteria were still present after 5 hours of incubation at this temperature (Figure 2). In a similar manner, Brucella heated in a heat block set at 75°C or 85°C demonstrated average reductions of 5.2 and 6.7 logs after 60 minutes of heat killing. However, mean numbers of viable bacteria were 2.7 ± 0.22 logs after 4-hour treatment at 75°C and 0.82 ± 0.30 logs after heat treatment at 85°C for 2 hours. Heat treatment in a heat block set at 95°C demonstrated an average reduction of 7.9 logs by 30 minutes, with an average viability of 0.28 ± 0.13 logs at 60 minutes. No isolates of 16M or RB51 were recovered after 60 minutes of heat treatment at 95°C.
Concentrations of Brucella abortus strains 2308 and RB51, Brucella suis strain 1330, and Brucella melitensis strain 16M after heating at temperatures of 65°C, 75°C, 85°C, and 95°C. CFU, colony-forming unit.
Methanol-Acetone and Methanol Inactivation
Methods used in the methanol-acetone inactivation experiment should have detected live Brucella at concentrations >2.5 CFU/mL. Standard plate counts indicated that all samples contained >1010 CFU/mL of live bacteria at the onset of the experiment. All bacteria stocks suspended in the absence of methanol-acetone (0%) had a lawn of growth on plates at both sampling times, with standard plate counts indicating concentrations exceeding 1010 CFU/mL in these tubes at the end of the experiment. In both replicates, 2308 or 16M bacteria were recovered at low concentrations (reduction of 7 to 8 logs) at 40% methanol-acetone concentrations after either 3 or 6 days of incubation at 4°C. However, no live bacteria were recovered at methanol-acetone concentrations ≥50% at either sampling time. Our data suggest that initial concentrations of 1010 CFU/mL of Brucella are completely inactivated within 3 days in solutions of 50% or 70% methanol-acetone.
We calculated that our detection limit for the methanol inactivation experiment was 2.5 CFU/mL. Standard plate counts indicated that all tubes contained bacterial concentrations >8 × 1010 CFU/mL at the beginning of the experiment. A lawn of bacterial growth was observed at all sampling times from tubes without added methanol. At 3 days of incubation in the first replication, live bacteria were repeatedly recovered from most methanol concentrations, with the exception of 16M and 1330 in solutions containing 70% methanol. With the exception of recovery of live bacteria from tubes containing 30% methanol concentrations at 5 and 7 days in the first replication, no Brucella were recovered from remaining samples (40%-70% methanol) after 5 and 7 days of incubation. In the second replication, with the exception of 2308 in 50% methanol at 5 days, all samples in 50%, 60%, or 70% methanol were negative for recovery of live Brucella after 3, 5, or 7 days of incubation. With the exception of recovery of low concentrations (1.47 logs) of strain 2308 at 5 days in the first replication from a tube containing 50% methanol, all remaining tubes with methanol concentrations of 50% to 70% from both replications were negative for recovery of live Brucella at 5 and 7 days. Our data suggest that complete inactivation of live Brucella occurs within 5 days in solutions of 60% or 70% methanol when initial bacterial concentrations are 1010 CFU/mL.
Disinfectant Inactivation
Standard plate counts indicated that bacterial concentrations in this experiment exceeded 4 × 1010 CFU/tube (>1011 CFU/mL), and our detection limit was calculated as 5 CFU/mL. After incubation in Wexcide for at least 20 minutes, no colonies of strain 1330 or 2308 were recovered. Although no viable 16M bacteria were recovered after 30 minutes of incubation in Wexcide, approximately 1.5 logs of 16M were recovered at the 20-minute time point. When inactivated with Amphyl, no colonies of 16M or 1330 were recovered at 20 or 30 minutes. However, viable 2308 bacteria (approximately 1.5 logs) were recovered at 20 minutes after exposure to Amphyl, and a single colony recovered after 30 minutes of incubation. A lawn of growth was present on plates inoculated from tubes without disinfectants and from all tubes in which 0.1 or 0.01 concentrations of disinfectants were added.
Formalin Killing
After 2 hours in 10% buffered neutral formalin, 2 of 6 cotyledon samples obtained after abortion had recoverable B abortus but demonstrated a significant (P < .05) reduction in mean colonization of approximately 6.5 logs as compared with fresh (unfixed) samples (Table 1). After being immersed in formalin for 4 or 24 hours and rinsed in PBS before processing, Brucella were not recovered from any bison cotyledon samples examined.
Formalin Killing of Brucella abortus in Bison Cotyledon Tissue.

Abbreviation: CFU, colony-forming unit.
Means denoted with different superscripts are significantly different (P < 0.05).
Discussion
This article presents data validating selected inactivation methods for tissues, solutions, and sera that may contain virulent species in the Brucella genus. Although initial bacterial concentrations, sample volumes, or concentration of inactivation agents may differ among laboratories, these data provide baseline data that can be used to narrow parameters for inactivation validation in other facilities. Validation studies of this type are currently being required by the Select Agent program to ensure that live agents are not mistakenly being removed from containment areas/facilities registered for BSATs. The Brucella genus is very homologous, with an average >94% identity at the nucleotide level across B abortus, B suis, and B melitensis genomes. 5 For this reason, we chose to evaluate 1 representative strain from each of the 3 virulent Brucella species that are recognized as select agents. In some experiments, we included the attenuated B abortus RB51 vaccine strain, as its lipopolysaccharide structure differs from the 3 virulent strains. Overall, although we did find some minor differences in inactivation among strains, these differences have only minor significance on parameters for inactivating virulent Brucella strains.
Heat inactivation procedures, such as pasteurization, are commonly used to eliminate viability of Brucella in products such as milk. Early work in the 1920s reported that initial concentrations of B abortus at 5 × 109 CFU/mL were killed at 60°C in 15 minutes and similar populations of B suis in 20 minutes.6 –11 Data also indicated that heating at 55°C destroyed B abortus in 2 hours and temperatures of 61°C to 62°C killed all virulent B abortus strains in milk within 5 to 30 minutes. Many studies reported complete inactivation within 5 to 10 minutes.9,10,12,13 However, conclusions by authors in 1 report differed in that their data suggested that 40 minutes of heat treatment at 60°C was required to inactivate B suis in milk. 14 Differences among early reports could have been influenced by bacterial concentrations (107-1012 CFU/mL), the fluid medium (milk vs saline), isolation procedures (some inoculated inactivated cultures into guinea pigs), and incubation parameters, as some laboratories cultured Brucella under normal atmospheric conditions without the higher CO2 concentrations as required by most B abortus strains.1,12,13 However, the prolonged survival of Brucella after heat treatment was supported in a more recent report on preparation of heat-inactivated B abortus suspensions, where initial concentrations of 109 CFU/mL retained 95%, 55%, and 25% bacterial survival after heat treatment at 65°C, 75°C, or 80°C, respectively, for 120 minutes. 15 This report concluded that field strains of B abortus are more thermoduric than previously recognized, and it led to a subsequent study with milk from an infected dairy, which concluded that pasteurization procedures (63°C for 30 minutes or 72°C for 15 seconds) completely inactivated Brucella in milk. 16 Concentrations of Brucella shed in bovine milk are estimated to average 20 to 440 CFU/mL, with highest concentrations secreted in milk estimated at approximately 105 CFU/mL. 8 Therefore, concentrations in milk are probably 4 to 7 logs less than the concentrations used in most heat inactivation studies.
Although our data more closely concur with the more recent study, 15 our results are quite different from historical studies of Brucella reported in the 1920s and 1930s. However, our inactivation curves are similar to reported heat inactivation curves of other bacteria (Salmonella, Escherichia coli, Streptococcus) under in vitro conditions. 17 In the laboratory, heat treatment is attractive, as it does not require addition of chemicals to bacterial solutions and can be done relatively quickly as compared with other inactivation techniques. However, as Select Agent regulations require elimination of all viable bacteria before removal from registered spaces, our data suggest significant differences among temperatures in the time required to meet this criteria. In our experiment, complete elimination of viable Brucella bacteria within 30 to 60 minutes required temperatures approaching boiling, whereas lower temperatures required much longer heating times (hours). Although our data cannot eliminate the possibility that there may be differences among cultures in various growth phases (ie, exponential, stationary, or logarithmic decline) in susceptibility to inactivation, others have reported that 48- and 96-hour cultures of B abortus do not differ in susceptibility to heat inactivation. 10 Although our formalin inactivation study most likely evaluated bacteria in various stages of growth, the influence of stage of growth was not an parameter that we chose to evaluate in the current study. Our data did indicate minor differences in susceptibility to heat inactivation across Brucella species that may be consistent with other reports suggesting that porcine strains exhibit increased heat resistance.12 –14 However, as stated previously, these differences appear to have only minor effects on heat inactivation curves. It should also be emphasized that the current study did not use enrichment procedures after heat inactivation to try to recover viable Brucella in negative samples that might have had viable bacteria concentrations below our detection limit. However, as these concentrations would be well below the median infectious dose for highly susceptible host species (ie, guinea pigs), 18 if present, we believe the concentrations are not of biological or biosafety significance.
One influence on heat inactivation of live bacteria may be the time that it takes for a tube to reach the targeted temperature. This time is likely influenced by the target temperature, sample volume, surface area in contact with the heat source, thermal characteristics of the sample, and the properties of the container. The current experiment was not designed to evaluate all possible combinations but rather provide insight on the rate of Brucella inactivation across different temperatures under similar experimental conditions. As data from the current experiment suggest that internal temperatures generally reached equilibrium within 10 to 12 minutes without agitation, the short delay in reaching the targeted temperature does not explain the prolonged viability of Brucella within our samples during heat inactivation. As there will certainly be differences in procedures and equipment among laboratories that will influence inactivation of Brucella during heat treatment, our data strongly suggest that laboratories should validate any heat inactivation procedures currently being used for Brucella with equipment and samples appropriate to their conditions. Data reported here and in other studies raise concerns that in vitro heat treatments utilized for Brucella killing that are based on pasteurization procedures are likely insufficient for inactivation under laboratory conditions.
While treatment at or near boiling temperatures (95°C) can be effective, prolonged exposure to these temperatures may inhibit recovery of nucleic acids or proteins for downstream analysis. Therefore, use of methanol or methanol-acetone inactivation methods may be more appropriate for some downstream applications. Our results suggest that methanol and methanol-acetone allow complete elimination of viable Brucella but require fairly high concentrations (60% or 70%) and at least 3 to 5 days of incubation at refrigeration temperatures. Because we did have recovery of live Brucella at 3 days and at later time points when suspended in lower methanol or methanol-acetone concentrations, our data emphasize the importance of using bacteriologic methods to confirm that no Brucella bacteria remain viable in a sample before removal from biosafety level 3 containment. The disadvantage of these methanol and methanol-acetone inactivation procedures is the time required and the use of materials that may need to be stored in an explosion-proof refrigerator.
As 10% neutral formalin is a preferred fixative for pathologic studies, it was not unexpected that it would be active against Brucella. Even with placement of approximately 1.5-cm-diameter sections of infected cotyledon tissue containing high concentrations of bacteria, no viable Brucella were recovered by 4 hours after fixation. These results offer a high degree of certainty that formalin-fixed tissues can safely be handled and sectioned outside of biosafety level 3 containment if tissue sections of the appropriate size are initially obtained. In a similar manner, it was anticipated that the disinfectants evaluated would be effective even though label claims for Brucella were not made. By mixing disinfectants prepared in accordance with label directions with equal volumes of sample, our highest concentrations of disinfectants were actually at 50% of recommended concentrations. However, this was intentional because, when disinfectants are applied under working conditions, they are frequently diluted by surface conditions. It was somewhat surprising that even solutions containing only 10% of recommended disinfectant concentrations demonstrated significant reductions in bactericidal properties. In addition to interference by organic matter, our data suggest that ensuring adequate contact time and preventing dilution of the disinfectant solution are concerns for use of these disinfectants on contaminated surfaces or materials. Our data on the suitability of disinfectants are primarily related to biosafety and facility cleaning, as it is unlikely that Brucella inactivated with this method would be suitable for downstream laboratory applications due to the presence of residual and potentially inhibitory chemicals.
Last, our data suggest that filter sterilization with a 0.22-μm filter is highly effective in removal of high concentrations of Brucella from fluid samples. Extensive data from >7000 samples obtained from animals after experimental infection endorse this method of inactivation of serum samples.
Overall, our data suggest that even high concentrations of virulent species of Brucella can be readily inactivated with standard methods. Despite demonstration of the effectiveness of these procedures in our laboratory, it is highly recommended that others investigators validate similar procedures used under their own in vitro conditions. Heat inactivation appears to be highly influenced by temperature with heating to near 100°C required for rapid killing of all bacteria within samples. Although chemical methods were highly successful for inactivating Brucella, they were influenced by concentrations of the inactivating agent within the sample. Because even a single failure could result in significant consequences from violation of Select Agent regulations, it is highly recommended that culture methods be used, as possible, on all inactivated samples to verify the effectiveness of the validated procedure and protect against inadvertent laboratory errors.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.