
Abstract
The metastasis of lung cancer is the most prevalent cause of patient death. Various treatment strategies have targeted the prevention of the occurrence of metastasis. The epithelial–mesenchymal transition (EMT) in lung cancer cells is considered a prerequisite to acquire the invasive/migratory phenotype and to subsequently achieve metastasis. However, the effects of Rubus idaeus on cancer invasion and the EMT of the human lung carcinoma remain unclear. In this article, we test the hypothesis that R idaeus ethyl acetate (RIAE) possesses an antimetastatic effect and reverses the EMT potential of human lung A549 cells. We extract the raspberry R idaeus with methanol (RIME), chloroform (RICE), ethyl acetate (RIAE), n-butanol (RIBE), and water (RIWE). The RIAE treatment obviously inhibits the invasive (P < .001), motility (P < .001), spreading, and migratory potential (P < .001) of highly metastatic human lung cancer A549 cells. The zymography and promoter luciferase analysis reveals that RIAE decreases the proteinase and transcription activities of MMP-2 and u-PA. Molecular analyses show that RIAE increases the E-cadherin level that is mainly localized at the cellular membrane. This result was also verified through confocal analyses. RIAE also induces the upregulation of an epithelial marker, such as α-catenin, and decreases mesenchymal markers, such as snail-1 and N-cadherin, that promote cell invasion and metastasis. RIAE inhibits MMP-2 and u-PA by attenuating the NF-κB and p-Akt expression. The inhibition of RIAE on the growth of A549 cells in vivo was also verified using a cancer cell xenograft nude mice model. Our results show the anti-invasive/antitumor effects of RIAE and associated mechanisms, which suggest that RIAE should be further tested in clinically relevant models to exploit its potential benefits against metastatic lung cancer cells.
Introduction
Cancer chemoprevention is classically defined as the use of natural or synthetic agents, such as polyphenols, flavonoids, and alkanoids, alone or in combination, to prevent, arrest, delay, or reverse either the initiation phase of carcinogenesis or the progression of neoplastic cells to malignant cancer, and it is a promising area in current cancer therapy.1,2 Natural compounds with antioxidant properties that protect the human body against the development of tumors can be found in a variety of plants, vegetables, and fruits.3,4 Epidemiological evidence also indicate that increasing the intake of fruits and vegetables reduces the incidence of cancer and is responsible for their protective properties. 5 Raspberry or Rubus idaeus L. is a plant that belongs to the Rosaceae family. The plant contains significant amounts of polyphenol antioxidants and is native to numerous parts of Europe and North America. Black raspberry wines contain rutin, epicatechin, catechin, malvidin 3-glucoside, 3,4-dihydroxybenzoic acid, myricetin, gallic acid, p-coumaric acid, and quercetin. 6 Rubus idaeus has an impressive prophylactic effect on renal stones formation in a mouse model of hyperoxaluria. 7 Earlier reports indicated that black raspberry extracts could reduce the growth rates of colon cancer HT-29 cells in vitro. 8 The ethanol extract of black raspberry inhibits cell proliferation and regulates apoptosis in human cervical cancer cells (SiHa, C33-A, and HeLa cell lines) in a dose- and time-dependent manner. 9 However, the effect of raspberry on cancer invasion and the metastasis of human lung cancer cells remains unclear.
Lung carcinoma, which is derived from epithelial cells, is the main cause of cancer-related deaths in Western and Asian populations with a 5-year survival rate of only approximately 15% and is the most commonly diagnosed malignancy. 10 Non–small cell lung cancer (NSCLC) is the most common type of lung cancer. About 85% of lung cancers are NSCLC. Adenocarcinoma, squamous cell carcinoma, and large cell carcinoma are all subtypes of NSCLC. Highly invasive malignance lung carcinoma cells exhibit an increase in matrix metalloproteinase (MMPs) or urokinase-type plasminogen activator (u-PA) activities, which results in an increase in proteolytic activities in specific sites, such as areas of cell-to-the extracellular matrix (ECM) or cell-to-cell contacts, to promote cell invasion and metastasis. 11 Most patient deaths from lung cancer are related to metastasis, which is a complicated and currently uncontrolled process. 11 Therefore, reducing the metastasis of lung tumor cells is considered to be one of the most important research areas in medicine and in the development of effective strategies in treating or preventing the metastatic stage of lung carcinoma.
Metastasis is one of the hallmarks of cancer cells and is an extremely complex biological event, where the cancer cell invasion of new tissues involves the coordination of several signaling pathways that allow the detachment of tumor cells, degradation of the ECM, attachment of circulating malignant cells to endothelial cells, extravasation or invasion out of the blood vessel, and reestablishment of growth at a distant site.12,13 Several proteases may be involved in proteolytic events, including serine proteinase, plasminogen activator, MMPs, and cathepsins, to degrade ECM and allow cancer metastasis. MMPs, including MMP-2 and MMP-9, and u-PA can directly degrade most of the components of ECM and are deeply involved in tumor growth, angiogenesis, invasiveness, and metastasis. 14 Therefore, the attenuation of the migration or invasion mediated by MMPs or u-PA may be a key feature in preventing cancer development and metastasis.13-15 Recent studies have demonstrated that the mitogen-activated protein kinases (MAPKs) family and the phosphatidylinositol 3-kinase (PI3K)-Akt pathway are critical points in regulating cell proliferation, survival, invasion, and the expression of MMPs and u-PA. 16
Epithelial–mesenchymal transition (EMT), a dedifferentiation program that converts adherent epithelial cells into individual migratory cells, is considered a key factor in the induction of oncogenic progression, tumor malignancy, and cancer metastasis. 17 EMT is a complicated multistep process because it is not only a simple morphological change but is also associated with the transcriptional or expressional alterations of several epithelial and mesenchymal genes. 18 For example, keratinocytes at wound tissues display a migratory action that precipitates wound healing. Epithelial cancer cells from in situ tumor or metastatic carcinoma lose contact with their neighbors because of the reduced expression of E-cadherin or α-catenin and the elevated expression of vimentin through the EMT program. 19 The epithelial molecule E-cadherin, which is known for regulating cell-to-cell contact, cell polarity, and cell shape, 20 connects adjacent cells through homophilic interactions and is linked to the cytoskeleton though the multi-catenin complex, including α-catenin and β-catenin, which are attached to their cytoplasmic tails. In this complex, β-catenin is directly associated with E-cadherin, whereas α-catenin is the link between β-catenin and the actin microfilament network of the cytoskeleton. 21 The loss of E-cadherin expression is positively correlated with tumor grade and stage, 22 and the aberrant or decreased expression of E-cadherin is considered to be one of the biomarkers for a poor prognosis in lung cancer. 23 Epithelial cells during EMT downregulate epithelial markers, such as E-cadherin and α-catenin, and acquire mesenchymal markers, such as N-cadherin, Vimentin, and Fibronectin.19,24 Several transcriptional factors, including Snail-1, Slug, Zeb-1, and Twist, were also identified to negatively regulate E-cadherin expression and to have an important function in EMT induction and in the maintenance of invasive and migratory phenotype in cancer cells.25-27 Akt reportedly upregulates Snail-l and CD44 expression, which promotes EMT. 28 Earlier reports have indicated that EMT is involved in drug resistance and the metastasis of lung cancers. 29 Raspberries may clearly inhibit the growth of various cancers by inducing cancer cells toward apoptosis 9 and scavenging reactive oxygen species, 30 but studies concerning the potential of raspberry extracts in retarding cancer cell invasion and reversing EMT transition remain limited. These studies suggested that raspberries may have enormous potential in cancer therapy. However, the effect of R idaeus extracts on cancer migration, the EMT of human lung carcinoma, and the underlying mechanisms of such effect remain unclear. In this study, we test the hypothesis that R idaeus ethyl acetate (RIAE) possesses an antimetastatic effect and reverses the EMT potential of human lung A549 cells.
Materials and Methods
Materials and Chemicals
Dulbecco’s modified Eagle medium (DMEM) was obtained from Sigma Chemical Co (St Louis, MO). Matrigel was purchased from BD Biosciences (Bedford, MA). Rabbit polyclonal antibodies against vimentin, E-cadherin, α-catenin, N-cadherin, p-Akt, Akt, p-GSK-3β, GSK-3β, and snail-1 were purchased from Cell Signaling Technology (Dancers, MA). A goat polyclonal antibody against β-actin was purchased from Santa Cruz Biotechnology Inc (Santa Cruz, CA). The Immobilon Western Chemiluminescent HRP substrate kit was obtained from Millipore (Burlington, MA).
Preparation of Rubus idaeus Extracts
The raspberry (R idaeus) purchased from a store (Taichung, Taiwan) was imported from Driscoll Strawberry Associates Inc (Watsonville, CA, USA). Their authenticity was confirmed by herb doctor Shang-Pang Huang. The air-dried raspberry (100 g) was extracted 3 times with boiling water (500 mL) for 30 minutes. The filtrate was partitioned with chloroform (RICE), ethyl acetate (RIAE), and n-butanol (RIBE). The residue was then extracted with methanol (RIME). The solvent was removed, and the filtrate was then lyophilized and stored at −20°C. The chemical profile of RIAE was also analyzed using a high-pressure liquid chromatogram (HPLC)–mass spectrometer. RIAE was analyzed using an HPLC–mass spectrometer (Waters 600 with a 2998 Photodiode Array detector). The samples (10 µL) were injected into a Merck LiChrospher 100 RP-18 column (4 mm × 250 mm). The analyses were performed using the mobile phase composed of 2 solvents: solvent A (0.05% acetic acid/water) and solvent B (acetonitrile). The flow rate was 1 mL/min. Elution was performed in a programmed gradient elution as follows: 0 to 30 minutes, with 0% to 60% B; 30 to 35 minutes, isocratic with 100% B; and 35 to 40 minutes, isocratic with 100% A. The absorbance was monitored at 280 nm. The molecular masses of the peaks were determined from the electrospray ionization mass spectra by using a multiple-charged ion profile based on the modified method of Wong et al. 31
Cell Culture
The A549 (NSCLC; human lung adenocarcinoma) cell line obtained from ATCC (Manassas, VA) was cultured in DMEM supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 U/mL penicillin, and 100 µg/mL streptomycin. All cell cultures were maintained at 37°C in a humidified atmosphere of 5% CO2.
Transwell Cell Invasion and Motility Assays
The A549 cells were pretreated with RIAE at an indicated concentration for 24 hours. The cells were then harvested and seeded to Millipore cell culture inserts at 5 × 104 cells/well in a serum-free medium and were incubated for 12 hours at 37°C. For the invasion assay, 100 µL of Matrigel (1 mg/1 mL) was applied to 8-µm-pore-size polycarbonate membrane filters with a standard medium at the bottom chamber of the apparatus. After incubation, the filters were air-dried for 5 hours in a laminar flow hood. The invaded cells were fixed with methanol and stained with Giemsa. The cell numbers were counted using a light microscope. Motility assay was performed as described for the invasion assay with no coating of Matrigel. 32
Wound Healing Migration Assay
The A549 cells were seeded into a 12-well culture dish and grown in a culture media containing 10% fetal bovine serum (FBS) to a nearly confluent cell monolayer (>95% confluence) for the cell migration assay. In each case, the cells were made quiescent through pretreatment with 5 μg/mL mitomycin C for 2 hours to ensure that the wounds are filled because of cell migration and not through cell proliferation. The wounds were introduced to the confluent monolayer of cells with a sterile 200-µL plastic pipette tip to create a denuded area. The floating cells were removed by washing twice with phosphate-buffered saline and were replaced with a culture media containing 1% FBS. RIAE was then added. The cells were incubated at 37°C, and the cell movement into the wound area was photographed at 0 hours and 48 hours by using a microscope. The cell motility into a wound was expressed as the percentage of cell migration. 33
Cell Spreading Assay
After the treatment with RIAE, the cells were plated in type I collagen (10 µg/mL) coated 24-well plates and were cultured in culture media with or without RIAE for 1, 6, and 12 hours. The cell morphology was photographed for morphological studies. 34
Determination of MMPs and u-PA by Zymography
The collected media were subjected to 0.1% gelatin–8% SDS polyacrylamide gel electrophoresis (SDS-PAGE) to determine the MMPs for the gelatin zymography. The gels were then washed with 2.5% Triton X-100 and incubated in a reaction buffer for 16 hours at 37°C. The gel was then stained with Coomassie brilliant blue R-250. u-PA activity was visualized through casein zymography, where 2% w/v casein and 20 µg/mL plasminogen were added to 8% SDS-PAGE gel. Casein zymography was performed as described in gelatin zymography. 35
Measurement of MMP-2 and u-PA Promoter Activity
A 460 bp (−218 to +243) segment from the 5′-promoter region of the MMP-2 gene and a 644 bp (−562 to +83) segment from the 5′-promoter region of the u-PA gene were cloned. pGL3-MMP-2 and -u-PA plasmid were transfected into A549 cells by using the PolyJet reagent (SignaGen Laboratories, Gaithersburg, MD) based on the manufacturer’s instructions. The cells were collected and disrupted by the Luciferase Assay System (Promega, San Diego, CA) after incubation with RIAE. After centrifugation, the aliquots of the supernatants were tested for luciferase activity by using the Luciferase Assay System. The firefly luciferase activities were standardized for β-galactosidase activity. 36
Immunofluorescence Staining
The cells were cultured on sterile glass coverslips in 6-well plates. The slides were incubated overnight at 4°C with an E-cadherin antibody and were then incubated with an FITC-conjugated anti-rabbit Ig at room temperature for 1 hour. The slides were counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and were analyzed through microscopy. 34
Western Blot
The cell lysate samples were separated in a 12.5% polyacrylamide gel and were transferred onto a nitrocellulose membrane as previously described. 35 The blot was subsequently operated with standard procedures and was probed with primary and secondary antibodies. The protein expression was detected through chemiluminescence by using an Immoblon Western Chemiluminescent HRP Substrate kit.
Snail-1 siRNA
Snail-1-specific siRNA were obtained from Invitrogen (Carlsbad, CA). Forward transfections were performed using Lipofectamine RNAiMax (Invitrogen), following the guidelines indicated by the manufacturer. After the treatment, the cell lysate and unclear extracts were extracted and analyzed by Western blot.
Measurement of Tumor Growth in Nude Mice
This study was performed in strict accordance to the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Institutional Animal Care and Use Committee (IACUC) of the institutional animal welfare guidelines of the Chung Shan Medical University (IACUC Approval Number: 822). For the nude mice xenograft model, 5- to 6-week-old immunodeficient nude mice (BALB/c AnN.CgFoxnnu/Crl Narl mice; from National Taiwan University Animal Center, Taiwan) weighing 18 to 22 g were used. The mice were housed with a regular 12-hour light/12-hour dark cycle and ad libitum access to a standard rodent chow diet (Laboratory Rodent Diet 5001, LabDiet, St Louis, MO) and were kept in a pathogen-free environment at the Laboratory Animal Unit. A549 cells (1 × 107 cells/0.1 mL/mouse) were injected subcutaneously into the right front axilla. The mice at 8-day postimplantation were randomly divided into 3 groups (n = 5 for each group) and were fed by oral gavage with water (control group) and RIAE (50 mg/day/kg and 100 mg/day/kg) suspended in water. The day of the cell implantation was designated as day 0. The average tumor volume at the start of the treatment was approximately 208 mm3, and the tumors of the treated and control groups were measured daily by using Vernier calipers to measure the long and short dimensions after 8 days of cell injections. The body weight was assessed daily after cell injection. On day 41, the tumor-bearing mice were euthanized with CO2 with minimal suffering. The primary tumors were separated from the surrounding muscles and dermis and were then weighed.
Immunohistochemistry Analysis
The paraffin-embedded slides were deparaffinized. Antigen unmasking was performed by microwave heating in citrate buffer for 20 minutes. The slides were incubated with 3% H2O2 and then with goat serum for 30 minutes at room temperature. The slides were washed with phosphate-buffered saline and then incubated with primary anti-Ki67 antibodies at 4°C overnight. Biotinylated secondary anti-mouse antibodies were added and incubated at room temperature for 1 hour. The sections underwent 30 minutes of streptavidin-HRP and were stained with a DAB substrate.
Statistical Analysis
The statistical significances were analyzed through a one-way analysis of variance (ANOVA) with post hoc Dunnett’s test. A P value of <.05 was considered statistically significant (Sigma-Stat 2.0, Jandel Scientific, San Rafael, CA).
Results
Effects of Rubus idaeus Extracts on the Invasion and Motility of A549 Cells
We successively extracted R idaeus with methanol (RIME), chloroform (RICE), ethyl acetate (RIAE), n-butanol (RIBE), and water (RIWE) to evaluate the bioactive compound of R idaeus (Figure 1A). The A549 cells were treated with different fractions (each concentration is 30 µg/mL). A decrease of invasion was detected by the Transwell invasion and motility assay. RIAE was the most efficient R idaeus ingredient in decreasing the invasion (Figure 1B and D) and motility (Figure 1C and E) activities in our assay.

Effects of Rubus idaeus extracts on the invasion and motility of A549 cells. (A) Procedure for the fractionation of the extracts from R idaeus. A549 cells were treated with these fractions by the Transwell invasion (B and D) and motility assays (C and E). The statistical significance of the results was analyzed using one-way ANOVA with post hoc Dunnett’s test (*P < .05, **P < .01, ***P < .001).
RIAE Exerts Strong Anti-Motility, Anti-Invasion, and Anti-Migration Efficacy Against A549 Cells
The RIAE treatment (10 to 50 µg/mL) inhibited the invasion (Figure 2A) and motility (Figure 2B) potential of A549 cells by 26% to 91% and 22% to 94%, respectively, in a dose-dependent manner. A wound healing migration assay was performed to assess if the RIAE affects cell migration. The incubation of A549 with 1% FBS produced a marked cell migration in the wound area 48 hours after wounding, whereas the wounds treated with RIAE showed pronounced delays in wound healing in the same conditions. The results showed that RIAE inhibited cell migration in a dose-dependent manner (Figure 2C).

Effects of RIAE on cell invasion, motility, and spreading. The A549 cells were analyzed for cell (A) invasion, (B) motility, (C) migration, (D) spreading, and (E) adhesion as described in the Materials and Methods section. The determined migration ability of A549 was subsequently quantified with the control group as 100% (without RIAE for 48 hours). The results were statistically evaluated using one-way ANOVA with post hoc Dunnett’s test (**P < .01 and ***P < .001). The results from 3 repeated and separated experiments were similar.
Inhibition on Cell Spread and Adhesion of A549 Cells by RIAE
Cancer cells that invade the host tissue break from their cell–cell contacts and make new contacts with the ECM. Therefore, we examined the effect of RIAE on the spreading potential of the A549 cells. Most of the control (without RIAE) cells exhibited spread morphology within 12 hours of plating. However, the cells treated with RIAE (50 µg/mL) remained round at the same time point (Figure 2D). The RIAE was then tested to determine its effects on the cell–matrix adhesion. The results showed that RIAE significantly reduced the cell–collagen interactions of A549 cells (Figure 2E).
RIAE Declines MMP-2 and u-PA
The effects of RIAE on MMP-2 and u-PA were investigated through gelatin and casein zymographies, respectively, in conditions of serum starvation to clarify if MMPs and u-PA were involved in inhibiting the invasion and migration of A549 cells by RIAE. RIAE reduced the activities of MMP-2 (P < .001; Figure 3A) and u-PA (P < .001) in A549 cells (Figure 3B).

Effects of RIAE on the protein and transcription activities of MMP-2 and u-PA. The cells were treated with RIAE for 24 hours and were then subjected to gelatin and casein zymographies to analyze the activities of (A) MMP-2 and (B) u-PA, respectively, as described in the Materials and Methods section. The luciferase activity was measured in transiently transfected A549 cells by using (C) pGL3-MMP-2 and (D) pGL3-u-PA. The data are expressed as mean ± SD with the control group as 100%. The statistical significance of the results was analyzed using one-way ANOVA with post hoc Dunnett’s test (*P < .05, **P < .01, ***P < .001).
Inhibitory Effect of RIAE on the Transcriptional Activity of MMP-2 and u-PA Promoter in A549 Cells
We performed a transient transfection with the pGL3-MMP-2 and pGL3-u-PA promoter and analyzed the luciferase activities to evaluate the effects of RIAE on the MMP-2 and u-PA promoter. The MMP-2 (P < .01; Figure 3C) and u-PA (P < .001; Figure 3D) luciferase activities of the transfectants treated with RIAE were reduced in a dose-dependent manner.
Attenuation of Akt Phosphorylation by RIAE
A treatment of RIAE to A549 cells inhibited the cell invasion and activities of MMP-2 and u-PA. We further investigated the underlying mechanisms. RIAE obviously reduced the activation of Akt and elevated the phosphorylated-GSK-3β, a critical downstream element of the PI3K/Akt pathway whose activity can be inhibited by the Akt-mediated phosphorylation of GSK-3β, in A549 cells in a dose-dependent manner (Figure 4A).

Effects of RIAE on the cytoskeleton related protein. Western blot of (A) p-Akt (Ser473) and p-GSK-3β (Ser9) in A549 cells after 24 hours of treatment with RIAE. (B) Western blot of E-cadherin, N-cadherin, α-catenin, and vimentin with β-actin as an internal control sample. (C) Immunofluorescence analysis of E-cadherin in A549 cells with or without a 50 µg/mL RIAE addition. (D) The nuclear extracts were subjected to SDS-PAGE followed by Western blot with anti-NF-κB and anti-snail-1 antibodies with the anti-C23 as an internal control sample. Similar results were obtained from 3 repeated and independent experiments.
RIAE Targets Signaling Molecules Regulating EMT in A549 Cells
We examined the RIAE effect on major regulators and markers of EMT. RIAE significantly elicited the upregulation of epithelial markers, such as E-cadherin and α-catenin, and decreased the mesenchymal markers, such as N-cadherin, but the RIAE treatment did not affect the vimentin expression (Figure 4B). The confocal analysis also confirmed that 50 µg/mL RIAE promotes E-cadherin expression in A549 cells (Figure 4C). RIAE significantly decreased the transcription factor nuclear protein expression of snail-1 and NF-κB with C23 as the internal control sample in A549 cells (Figure 4E).
Inhibitory Effect of RIAE on A549 Cell Invasion and Protease Secretion via the PI3K/Akt Signaling Pathway
We investigated the effects of the specific inhibitor of the PI3K pathway (LY294002) on A549 cells to determine further if the inhibition of MMP-2 and u-PA secretions by RIAE were mainly done through the inhibition of the PI3K/Akt signaling pathway. The results showed that a sole treatment with LY294002 led to the attenuation of cell invasion (P < .001) similar to that of RIAE (P < .001). The combined treatment of the inhibitor with RIAE could further decrease the invasion ability (Figure 5A). A similar trend for the reduction of the MMP-2 (Figure 5B) and u-PA (Figure 5C) of A549 cells by sole and combination treatments were also observed. Therefore, the decline of the PI3K/Akt signaling pathway may reduce the activities of MMP-2 and u-PA and tumor cell invasion. The effects of LY294002 on NF-κB were investigated by Western blot to determine the role of Akt in the regulation of the NF-κB expression. Sole LY294002 treatment decreased the NF-κB expression, which suggests that the PI3K/Akt may be upstream of the NF-κB pathway (Figure 5D).

Effects of RIAE, LY294002, and snail-1 siRNA on A549 cells. (A) The cells with the indicated pretreatment were subjected to in vitro invasion assay. The condition media were subjected to gelatin and casein zymographies to analyze the activities of (B) MMP-2 and (C) u-PA, respectively. (D) Western blot of NF-κB with C23 as an internal control sample. (E) The cells were treated with snail-1 siRNA, and/or RIAE. The cell lysates were then subjected to Western blot with anti-snail-1 and anti-E-cadherin antibodies. Similar results were obtained from 3 repeated and independent experiments. The results were statistically evaluated using one-way ANOVA with post hoc Dunnett’s test (**P < .01 and ***P < .001).
Increase in the E-Cadherin Level by RIAE Is Partly Caused by the Downregulation of Snail-1 Expression in A549 Cells
We used snail-1 siRNA to examine further the role of snail-1 in the regulation of the E-cadherin expression. Figure 5E shows that snail-1 siRNA strongly decreased the level of snail-1 after 48 hours of treatment. The knockdown of snail-1 expression by snail-1 siRNA was accompanied by an increase in the E-cadherin level in the A549 cells. We silenced the snail-1 expression by snail-1 siRNA and RIAE treatment to determine if RIAE increases the E-cadherin level by other pathways independent of snail-1. The Western blot results showed that the combined treatment of RIAE slightly increases the E-cadherin level when the snail-1 level is selectively inhibited, which suggests that a part of the function of snail-1 in the inhibition by RIAE leads to an increase in the E-cadherin level (Figure 5E).
Antitumor Effects of RIAE In Vivo
A549-bearing nude mice were treated with water or RIAE to verify the in vivo antitumor effects of RIAE. Small solid tumors were observed 8 days after cell inoculation, and a 2.5- and 5.1-fold reduction in the RIAE-treated (50 mg/kg and 100 mg/kg) animals were observed on day 41 compared with that of the control animals (Figure 6A and B). RIAE (100 mg/kg) feeding also induced a 5.6-fold reduction in tumor weight by day 41 (Figure 6C) without any apparent signs of toxicity as proven by the body weight monitoring (Figure 6D) throughout the experiment. A significant increase in the proliferation determined by Ki-67 stain in tumors was observed, which is consistent with the profound effect on tumor size (Figure 6E).

In vivo antitumor effects of RIAE. After the subcutaneous implantation of the A549 cells, the BALB/c nude mice were treated with saline or RIAE and were analyzed for tumor growth. (A) Control group (water), 50 mg/kg/day RIAE, and 100 mg/kg/day RIAE, (B) average tumor volume, (C) tumor weight, and (D) body weight. (E) Immunohistochemistry for Ki-67 (cell proliferation marker) in A549 tumors (200×). The results were statistically evaluated using one-way ANOVA with post hoc Dunnett’s test (**P < .01).
Identification of RIAE Fractions
HPLC was performed to analyze the detailed compositions of RIAE. The chromatographic patterns from the HPLC analysis of RIAE showed peaks that correspond to the retention times (Figure 7C). The absorbance was monitored at 280 nm. Compared with 16 kinds of standard compound (Figure 7A and B) and RIAE (Figure 7C), Table 1 show that gallic acid, 3,4-dihydroxybenzoic acid (3,4-DBA), epigallocatechin (EGC), epicathchin (EC), rutin, and epicatechin-3-gallate (ECG) are contained in the composition of RIAE.

Chemical profile of RIAE analyzed via HPLC–mass spectrometry. (A) HPLC chromatogram of 13 kinds of standard compound. Peaks: 1, 5 µg of gallic acid; 2, 10 µg of GC; 3, 5 µg of protocatechuic acid; 4, 5 µg of EGC; 5, 5 µg of catechin; 6, 5 µg of methyl gallate; 7, 5 µg of EC; 8, 5 µg of EGCG; 9, 1 µg of GCG; 10, 5 µg of rutin; 11, 5 µg of ECG; 12, 10 µg of naringin; 13, 5 µg of quercetin. (B) Standard compound. Peaks: 14, 25 µg of 3,4-dihydroxybenzoic acid (3,4-DBA); 15, 25 µg of ρ-coumaric acid; 16, 8 µg of myricetin. (C) The chromatographic patterns from the HPLC analysis of the RIAE extracts showed peaks that correspond to the retention times (minutes).
Characterization of Phenolic Compounds of RIAE.
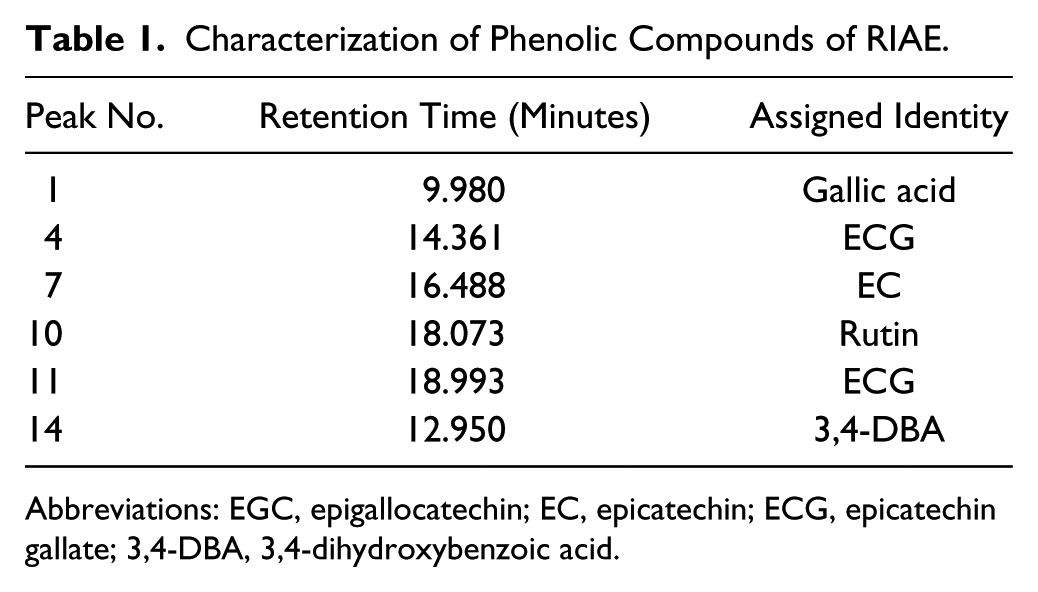
Abbreviations: EGC, epigallocatechin; EC, epicatechin; ECG, epicatechin gallate; 3,4-DBA, 3,4-dihydroxybenzoic acid.
In summary, these findings suggest that RIAE can transcriptionally regulate MMP-2 and u-PA expressions via the downregulation of the Akt pathway and can reverse EMT by modulating the E-cadherin expression through the repression of the snail-1 expression (Figure 8).
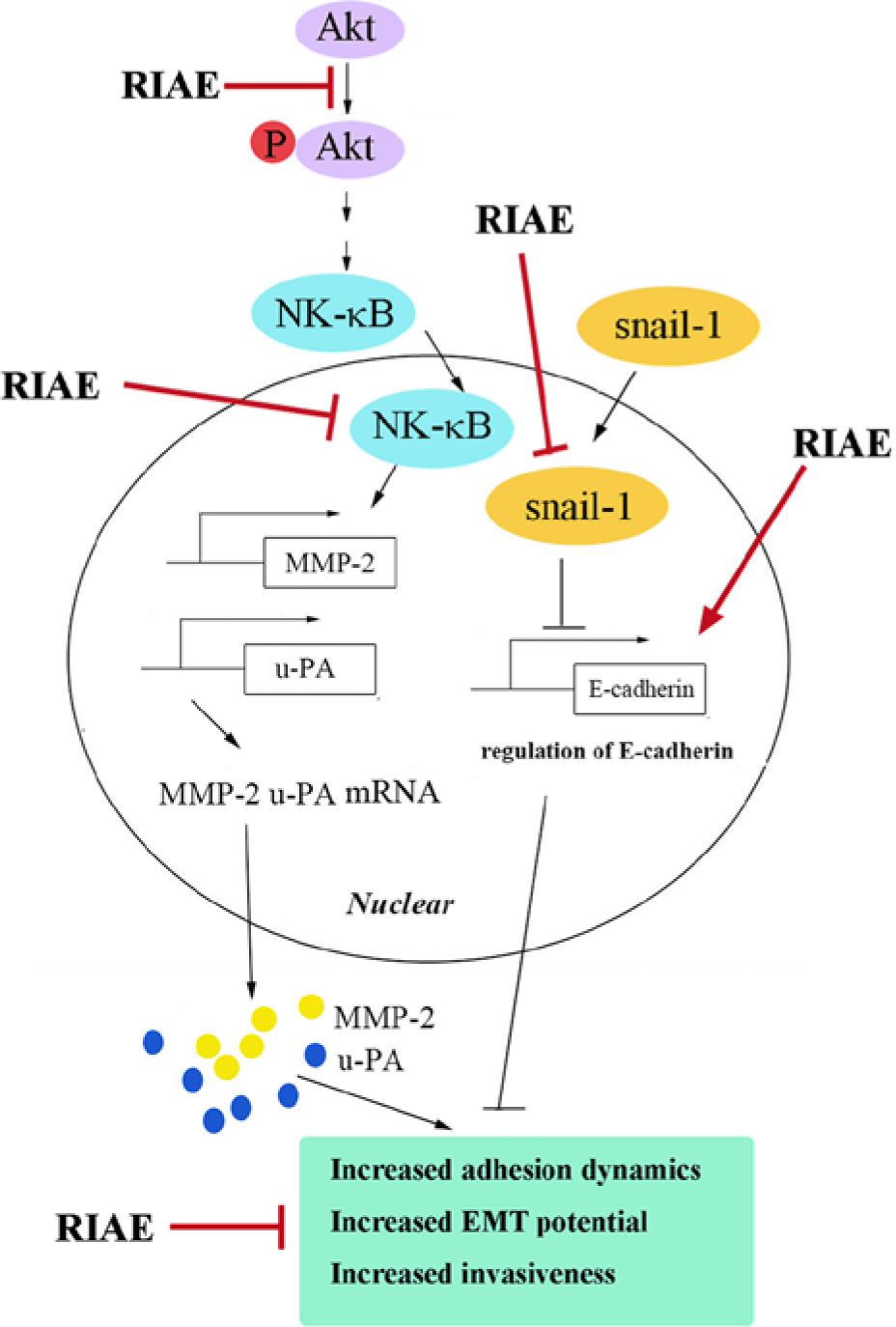
Proposed molecular targets in the anti-migratory and anti-invasive efficacy of RIAE in A549 cells. RIAE targets the snail-1-mediated E-cadherin expression and the translocation activities of MMP-2 and u-PA, which prevents the loss of cell–cell contact and inhibits the invasive and EMT potential of A549 cells.
Discussion
Most patient death occurs because of cell metastasis to the brain or bone in most lung cancer cases.37,38 Therefore, treatment against lung cancer is one of the most important areas of research in medicine and in the development of new and better strategies in treating or preventing the metastatic stage of lung carcinoma to improve patient outcome. An alternative approach to the decline of cancer progression can possibly contribute to the increased interest in chemoprevention. One approach in the management of advanced lung carcinoma could be the use of natural products, which are also known as “nutraceuticals.” These products are relatively nontoxic, low cost, physiologically bioavailable, and have multiple molecular targets in different types of cancer cells.39,40 Single-agent intervention, such as chemotherapy drugs, fails to produce the expected outcome in clinical trials, which leads to the increased popularity of combinations of nutraceuticals. Therefore, combinations of natural compounds that mimic real-life situations and are competent in targeting multiple targets with very little or virtually no toxicity are needed.
Therefore, the results from this study are highly significant because we report for the first time that RIAE (a) inhibits the invasive and migratory potential of a highly metastatic A549 lung cancer cell line; (b) elevates the expression of E-cadherin; (c) decreases phospho-Akt, Snail-1, and NF-κB levels and increases the phospho-GSK-3β and α-catenin expression; (d) reduces the proteinase and transcription activities of MMP-2 and u-PA via the downregulation of p-Akt; and (e) anti-invasive and antimigratory effects are in part by promoting E-cadherin and decreasing the snail-1expression in A549 cells.
The EMT process disrupts E-cadherin-mediated cell–cell adhesion, elicits N-cadherin expression, and changes the epithelial phenotype into a looser spindle-like mesenchymal morphology, which leads to invasion and metastasis. 17 The epithelial molecule E-cadherin, which is known for regulating cell-to-cell contact, cell polarity, and cell shape,20,41 connects the adjacent cells through homophilic interactions and is linked to the cytoskeleton though the multi-catenin complex, including β-catenin and α-catenin. 21 Earlier reports have indicated that the partial loss of E-cadherin and α-catenin is associated with poor prognosis and cancer progression in different types of human cancer cells.23,42,43 An overexpression of E-cadherin levels in some invasive carcinoma cells can inhibit migratory and metastatic abilities, but knockdown or blocking the E-cadherin expression in noninvasive cancer cells activates their invasiveness and metastasis efficiency. 44 In various types of tumor cells, the expression of E-cadherin is lost at an early stage of tumor development, which causes the resulting tumors to exhibit mesenchymal phenotypes that promote cell invasion and metastasis. This study shows that RIAE significantly decreased the expression of N-cadherin and increased the α-catenin level, which is known to bind with E-cadherin and promote cell-to-cell contact. 45 Therefore, RIAE treatment could increase the E-cadherin level at the cellular membrane and promote E-cadherin and α-catenin association, which restores the cell-to-cell contact that is detrimental to the invasive potential of A549 cells. Future studies are needed to understand the effect of RIAE effect on such molecular interactions between E-cadherin and α-catenin.
Several transcription factors were implicated in the transcriptional repression of E-cadherin, including zinc-finger proteins snail-1, ZEB-1, slug, SIP-1, and basic helix-loop-helix factors E47 and Twist.25-27,46 These transcription factors can reduce a subset of the genes that encode E-cadherin, claudin, occludin, and cytokeratin to induce EMT and tumor invasion. We demonstrated that RIAE could repress the snail-1 expression and increase the E-cadherin protein level. However, the regulation of RIAE on the expression of other transcription factors, such as Twist-1, slug, and ZEB-1, in A549 cells should be further defined. Our studies showed that snail-1 knockdown results in E-cadherin expression that is significantly increased by RIAE treatment and might be responsible for the biological effects of RIAE against metastatic A549 cells.
The downregulation of epithelial markers E-cadherin and α-catenin are expected during EMT, and Akt is reported to upregulate the snail-l expression, which promotes EMT. 28 This process is coordinated by the concomitant appearance of MMP secretion. Highly invasive ovarian cancer cells exhibited an increase in u-PA, u-PA receptor, MMP-2, or MMP-9 activity, which results in strong proteolytic activity in specific sites, such as areas of cell-to-cell or cell-to-ECM contacts, to promote cell invasion and metastasis. 14 The Sonic hedgehog (Shh) expression in the gastric tumor was positively correlated with lymph node metastasis, and the phosphoinositide 3-kinase/Akt depletion blocked the Shh-induced EMT, which is the activity of matrix metalloproteinase-9, and tumor invasiveness and metastasis. 47 In the present study, RIAE treatment inhibited the phosphorylation of Akt with a concomitant reduction in the levels of MMP-2 and u-PA. The involvement of the PI3K/Akt pathway was further supported by the use of the PI3K-specific inhibitor LY294002 in our experimental model, which shows that treatment with LY294002 can inhibit the expressions MMP-2 and u-PA and diminish the invasion of A549 cells. The results from this study show that the RIAE-caused attenuation of MMP-2 and u-PA expression partly contributes to the antimigratory and anti-invasive efficacy of RIAE.
Specific microRNAs (miRNAs) have been shown to regulate either a single step or multiple steps of cancer metastasis and EMT by downregulating the expression of their target genes. 48 The increasing awareness on the importance of miRNAs in tumorigenicity 38 has led to the accumulation of evidence that support the involvement of miRNAs in lung development. For example, the miR-17-92 cluster is required to promote the proliferation of lung progenitors, 49 whereas miR-34c, miR145, and miR-142-5p are required to suppress lung cell growth. 50 Earlier reports indicated that tea polyphenol (−)-epigallocatechin-3-gallate (EGCG) can suppress lung cancer cell growth by upregulating the miR-210 expression caused by stabilizing hypoxia-induced factor-1α. 51 Therefore, more studies are needed to investigate the potential cross-linking of the RIAE signaling with the microRNAs epigenetic regulation in lung cancer in the future.
Given the molecular complexity of the signaling pathways involved in cancer development and progression, multiple target therapy may be a successful strategy as an anticancer treatment. In conclusion, this study showed for the first time that RIAE can inhibit lung cancer cell invasion and migration not only by inducing E-cadherin and α-catenin but also by suppressing MMP-2 and u-PA transcription and enzyme activities through the Akt pathway. This study further suggests that RIAE can be a template for new anticancer drug development that can reverse EMT.
Footnotes
Authors’ Note
Yih-Shou Hsieh contributed equally as first author.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants of National Science Council, Republic of China (NSC 98-2313-B-166-004-MY3 and NSC 98-2313-B-040-004-MY3). Flow cytometry and confocal microscope studies were performed in the Instrument Center of Chung Shan Medical University, supported by National Science Council, Ministry of Education, and Chung Shan Medical University.