
Abstract
The use of multiple drugs in cancer therapy increases the efficacy of the potential therapeutic effects. In this study, the authors investigated the adjuvant effects of an ethanol extract of solid-state cultivated Taiwanofungus camphoratus (TCEE) and amphotericin B (AmB) in the human cancer cell lines RPMI7951 and MG63. Taiwanofungus camphoratus is a well-known Chinese medicine in Taiwan, and AmB is a widely used antifungal agent. The authors demonstrated that TCEE pretreatment followed by AmB treatment effectively inhibited cell growth. The combination of sublethal doses of TCEE and AmB revealed a significant growth inhibitory effect in both cell lines. The combination of TCEE and AmB but not AmB alone induced phosphatidylserine externalization and loss of mitochondrial membrane potential. Cell cycle analyses revealed that combination of TCEE and AmB triggered G2/M arrest and significant apoptosis after 48 hours. These effects were greater than those achieved using TCEE or AmB alone. Furthermore, the authors demonstrated that the drugs increased the levels of p21Cip1/Waf1 and pro-apoptotic protein Bax and reduced the level of anti-apoptotic protein Bcl-2. Taken together, the results showed that the combination treatment of TCEE and AmB displays strong adjuvant effects, which are indicated by the inhibition of cell proliferation in 2 human cancer cell lines, RPMI7951 and MG63. These findings suggest possible therapeutic applications and alternative medicines using this drug combination.
Introduction
The heptaene macrolide amphotericin B (AmB) is a broad-spectrum fungistatic and fungicidal drug that has been used for more than 50 years 1 to treat systemic fungal infections of immune-compromised hosts, such as patients with leukemia, 2 AIDS, 3 and organ transplantation candidates. 4 AmB interacts specifically with ergosterol-containing membranes and forms micelles. The insertion of the drug moieties of AmB into the acyl side chains of ergosterol induces the formation of the membrane pore complex and results in the efflux of certain main electrolytes from the cytosol. The disruption of membrane pumps and membrane potential induces apoptosis in the ergosterol-containing cells. 5 At high doses, essential cytoplasmic components, especially proteins, are released extracellularly. This leads to further apoptotic destruction of the cell. Cell shrinkage and reduction of intracellular electrolytes, particularly potassium ions, are accompanied by DNA fragmentation and caspase 3–like activity, leading to cellular apoptosis. 6 Recently, a cell model has been built that serves as a framework for understanding how the intracellular K+ and Na+ concentration changes induced by the cation-selective AmB channels enhance multiple survival response pathways before they are overcome by the more sustained ion fluxes, Ca2+-dependent apoptotic events, and cell lysis effects that are associated with the formation of AmB aqueous pores. 7
Taiwanofungus camphoratus (syn Antrodia camphorata) has been extensively used as a local remedy and has several pharmacological functions including antihepatotoxic, 8 antihypertensive, 9 anti-inflammatory,10,11 and antioxidant activities.12,13 In addition, its cytotoxic effects in various tumor cell lines have been demonstrated in many studies.14-17 The bioactive constituents of the ethanolic extract of Taiwanofungus camphoratus (TCEE) include diterpenoids, triterpenoids, sesquiterpene lactone, benzenoids, and polysaccharides. 18 Generally, extracts from T camphoratus inhibit proliferation and induce apoptosis in various types of cancer cell lines.15,19
The cocktail strategy that is used in cancer therapy has advantages such as reduced toxicity or drug resistance. AmB has been widely used to treat cancer patients, especially stem cell transplantation recipients and patients with leukemia, suffering from life-threatening systemic fungal infections. 20 A recent study has shown that AmB potentiates the cytotoxicity of cisplatin in various cancers.21,22 The possibility that AmB may also be used in cancer therapy has been proposed. 23 TCEE has been reported to possess potent anticancer effects. 18 However, it is not known whether a conventional antifungal agent, such as AmB, that is combined with a traditional Chinese remedy enhances inhibitory effects in cancer cells in vitro. Nevertheless, recently a novel dosing regimen of pretreatment with TCEE followed by the administration of AmB has been conducted to effectively induce apoptosis and inhibit tumor growth resulting in a significant repression of tumor growth in HT29 xenografts. 24 In this study, we further investigated the effects of the combination treatment of TCEE and AmB in RPMI7951 and MG63 cancer cells. We demonstrated the effects of this drug combination on specific key regulatory proteins of the cell cycle and apoptosis as previously reported. We demonstrated that combination treatment with TCEE and AmB significantly decreased cell viability and induced apoptosis in RPMI7951 and MG63 cells in a dose-dependent manner. The results indicate that the aforementioned drug combination exerts an anticancer effect in both cell lines. These findings demonstrate the significant anticancer activity of the combination treatment of TCEE and AmB.
Materials and Methods
Amphotericin B and TCEE Preparation
Amphotericin B was purchased from Sigma Aldrich Co (St Louis, MO). A pulverized crude extract of solid-state cultivated T camphoratus was provided by Well Shine Biotechnology Development (Taipei, Taiwan, ROC) and prepared as previously described. 11
Cell Culture
The human melanoma cell line RPMI7951 and the osteosarcoma cell line MG63 were purchased from the Food Industry Research and Development Institute (Hsinchu, Taiwan, ROC) and cultured in MEM that was supplemented with 10% fetal bovine serum (GIBCO, Paisley, Scotland), 100 IU/mL penicillin, and 100 mg/mL streptomycin (GIBCO, Grand Island, NY) under standard culture conditions (37°C, 95% humidified air with 5% CO2).
MTT Assay
The cells were seeded in 12-well culture plates at a concentration of 5 × 105/well 24 hours prior to treatment. The cells were pretreated with vehicle or low-cytotoxic doses of TCEE (100 µg/mL) for 24 hours, followed by treatment with vehicle or AmB (1.5-12 µM) for an additional 24 hours. 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide (MTT) solution (0.55 mg/mL) was added to each well and incubated at 37°C for 4 hours. The culture medium was removed and mixed with 500 µL of dimethylsulfoxide. Aliquots of 200 µL were transferred into 96-well plates. The optical density was measured at 550 nm using a microplate autoreader (Bio-Tek Instruments, San Diego, CA).
Annexin V–Propidium Iodide Staining and Mitochondrial Membrane Potential Analysis
Briefly, the cells (5 × 105) were cultured in 6-well plates and treated with TCEE (100 µg/mL) or vehicle for 24 hours followed by AmB (1.5-6.0 µM) or vehicle for an additional 24 hours. After treatment, the cells were collected and stained with annexin V and propidium iodide (PI) or stained with MitoProbe JC-1 (BioVision, Mountain View, CA) according to the manufacturer’s instructions. The resulting fluorescence was detected using flow cytometry (Becton Dickinson, Franklin Lakes, NJ).
Cell Cycle Analysis
The cellular DNA content was determined using flow cytometric analysis of PI-labeled cells. After the indicated treatment, the cells were harvested, fixed in ice-cold 70% ethanol, stored at 4°C, washed with phosphate-buffered saline (PBS; pH 7.2), treated with 25 mg/mL RNase A at 37°C for 30 minutes, and stained with 50 µg/mL PI for 30 minutes. A minimum of 10 000 cells per sample was used for analysis using CellQuest software. Apoptotic nuclei were identified as a subdiploid DNA peak and were distinguished from cell debris based on the forward light scatter and PI fluorescence.
Western Blot Analysis
The cells were harvested at the indicated times following the combined treatment with TCEE (100 µg/mL) and AmB (1.5-6.0 µM) (pretreatment with TCEE for 24 hours followed by treatment with AmB for an additional 24 hours) or vehicle alone. The harvested cells were pelleted by centrifugation, washed with PBS, and lysed with RIPA buffer. The protein content was measured using a protein assay kit (Pierce, Appleton, WI). A 40-µg aliquot of the total protein was separated using 12% sodium dodecyl sulfate-olyacrylamide gel and transferred to polyvinylidene fluoride membranes (Amersham Biosciences, Piscataway, NJ) for immunological detection of the proteins. The antibodies (Ab) that were used in this study included anti-Bax Ab, anti-Bcl2 Ab, anti-p21 Ab, anti-Cyclin B1 Ab (Upstate, Lake Placid, NY), anti-CDK2 Ab (Calbiochem, Darmstadt, Germany), anti-β actin mAb (Chemicon, Temecula, CA), anti-rabbit HRP-linked Ab, and anti-mouse HRP-linked Ab (Cell Signaling, Danvers, MA). The band intensity was quantified using an arithmetic analysis with ImageJ software (NCBI). The protein levels were normalized to β-actin levels and displayed as the relative expression values.
Statistical Analysis
The data were expressed as the means ± standard deviations (SDs) and analyzed using 2-tailed Student’s t test to assess the statistical significance. P < .05 was considered to be statistically significant.
Results
The Cytotoxicity of TCEE and AmB in RPMI7951 and MG63 Cancer Cell Lines
The MTT assay was used to investigate the effects of AmB and TCEE on cell proliferation in RPMI7951 and MG63 cells. As shown in Figure 1A and B, both AmB and TCEE inhibited cell proliferation in a dose-dependent manner. To investigate the combined effects of AmB and TCEE, low-toxic doses of AmB (0-12 µM) and TCEE (100 µg/mL) were applied. Two different treatment regimens (cotreatment and pretreatment) were performed to evaluate the effects of AmB plus TCEE. Although cotreatment with TCEE and AmB exhibited only mild antiproliferative effects, pretreatment with TCEE dramatically enhanced AmB-induced cytotoxicity (Figure 1C). Therefore, combination of TCEE and AmB in a sequential manner significantly inhibited the growth of RPMI7951 and MG63 cells compared with treatment with either drug alone (Figure 1A and B).

The cytotoxic effect of AmB and TCEE used alone or in combination
Combination of TCEE and AmB Induces Apoptosis in RPMI7951 and MG63 Cells
Previous studies have reported that extracts of T camphoratus induce apoptosis using in vitro culture systems 19 and that AmB induces the disruption of membrane pumps and the membrane potential to promote apoptosis in ergosterol-containing cells. 5 Therefore, we investigated whether the cytotoxicity that was induced by AmB, TCEE, or their combination was mediated by apoptosis. We examined the apoptotic effect of AmB alone and noted that the percentages of apoptotic cell death were 2.0 ± 0.7% at 0 µM AmB, 5.5 ± 1.0% at 3 µM AmB, and 6.1 ± 2.0% at 36 µM AmB. The percentages of viable cells (annexin V- and PI-negative cells) were 92.3 ± 1.3% at 0 µM AmB, 89.1 ± 1.3% at 3 µM AmB, and 86.9 ± 3.1% at 36 µM AmB (Figure 2A). Therefore, AmB that was used within the range of tested concentrations induced only mild apoptosis. In addition, we did not detect any dose-dependent effects following AmB treatment.

The apoptotic effects of TCEE and AmB in RPMI7951 and MG63 cells
We examined the apoptotic effects of the sequential treatment of TCEE and AmB. RPMI7951 and MG63 cells were treated with the combination of TCEE and AmB and stained with annexin V and PI. Figure 2B illustrates the fluorescent detection of PI and annexin V-FITC. The data revealed that the combined effects of the drugs significantly increased compared with the effect of either drug alone. The percentages of early apoptotic cells (annexin V–positive cells) in control cells were 2.0 ± 0.9% in RPMI7951 cells and 4.2 ± 0.6% in MG63 cells. However, 18.6 ± 2.2% and 10.3 ± 1.3% of apoptotic cells were observed in RPMI7951 and MG63 cells, respectively, following the combined treatment of 100 µg/mL of TCEE and 6 µM of AmB (P < .01). These results demonstrate that the combination of TCEE and AmB decreases cell viability and significantly increases apoptotic death in RPMI7951 and MG63 cells (P < .01; Figure 2C and D).
Combination of TCEE and AmB Leads to Loss of Mitochondrial Membrane Potential
We further confirmed the apoptotic effects of the combination of TCEE and AmB by detecting the loss of the mitochondrial membrane potential. As illustrated in Figure 3, sequential treatment of TCEE and AmB resulted in a dose-dependent decrease in the mitochondrial membrane potential in RPMI7951 and MG63 cells. In cells that were treated with 100 µg/mL of TCEE for 24 hours and 6 µM AmB for 24 hours, the JC-1 red/green fluorescence intensity ratio decreased to 6 ± 1.2% in RPMI7951 cells and 7.5 ± 1.0% in MG63 cells, indicating that the combination significantly induced mitochondrial dysfunction (P < .01), which confirmed our earlier observations.

Loss of the mitochondrial membrane potential in RPMI7951 and MG63 cells following the combined treatment with TCEE and AmB
To determine whether the drug combination altered Bcl-2 family protein levels, we examined the protein expression profiles of Bcl-2 and Bax. Sequential treatment of TCEE and AmB significantly reduced Bcl-2 levels and increased Bax levels (Figure 4A). The Bax-to-Bcl-2 ratio was determined for each combination after normalizing the protein levels to the β-actin protein level within each group (Figure 4B). The Bax-to-Bcl-2 ratio was significantly higher in cells that were pretreated with TCEE followed by higher doses of AmB (4.5 and 6 µM) compared with that in the control group (P < .05). Collectively, our results show that the sequential treatment of TCEE and AmB inhibits cell viability by increasing apoptosis.
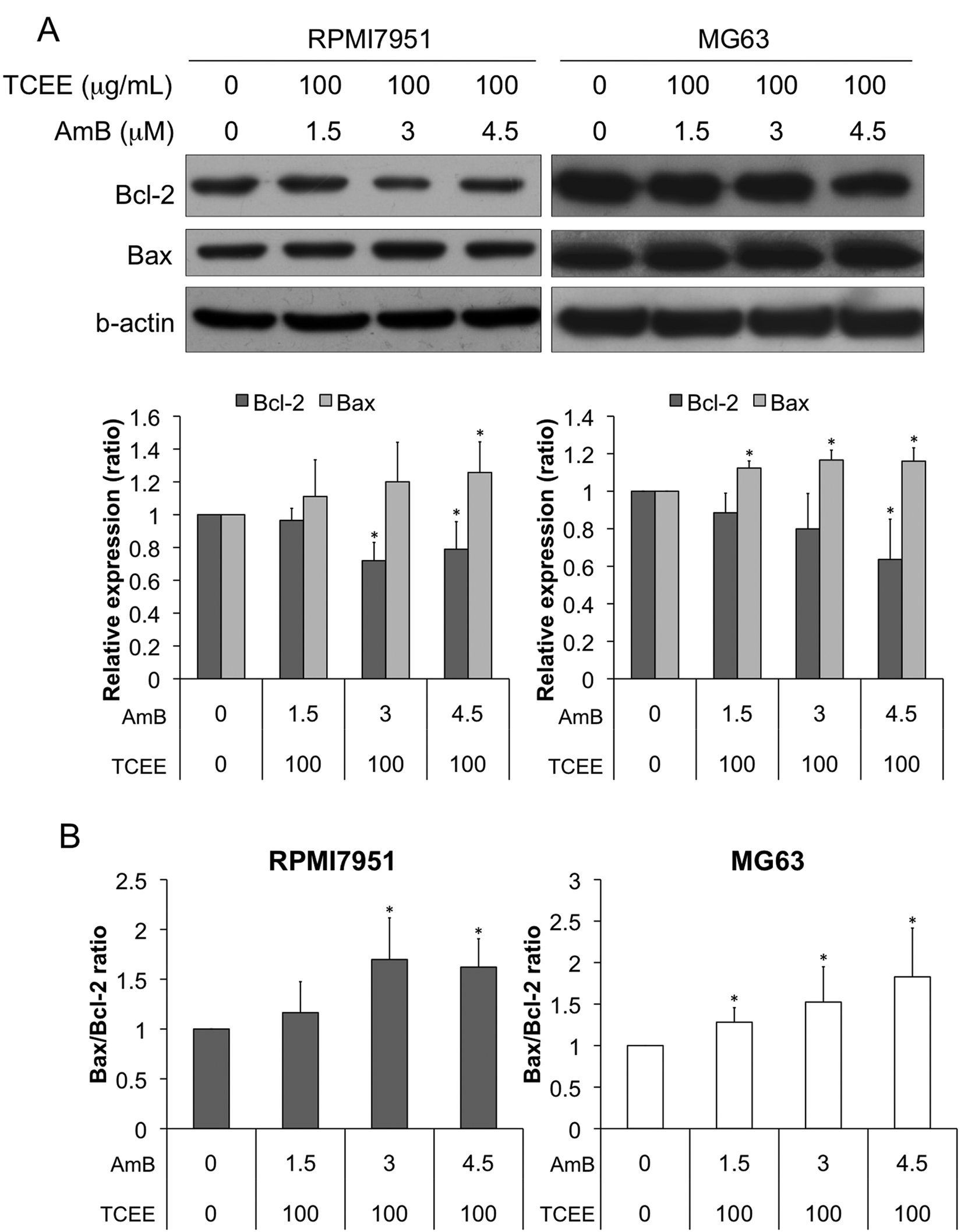
Increased levels of anti-apoptotic proteins in RPMI7951 and MG63 cells following the combined treatment with TCEE and AmB
Combination of TCEE and AmB Arrests the Cell Cycle in the G2/M Phase
In addition to apoptosis, cell cycle arrest may also contribute to the anti-proliferative effects of the combined treatment of TCEE and AmB. To verify this hypothesis, cell cycle analysis was performed in cells that were treated with different combinations of TCEE plus AmB. Although AmB alone did not alter the cell cycle distribution (Figure 5A), sequential treatment of AmB and TCEE arrested the cell cycle at the G2/M phase (P < .01; Figure 5B and C). Following the treatment with 100 µg/mL of TCEE and 3 µM AmB, the percentage of cells at the G2/M phase increased from 22.8 ± 0.8% to 32.0 ± 0.8% in RPMI7951 cells and from 19.4 ± 2.0% to 37.4 ± 2.4% in MG63 cells. The numbers of cells in the sub-G1 population were elevated in both cell lines. The results indicate that the combined treatment of AmB and TCCE arrests the cell cycle in the G2/M phase.
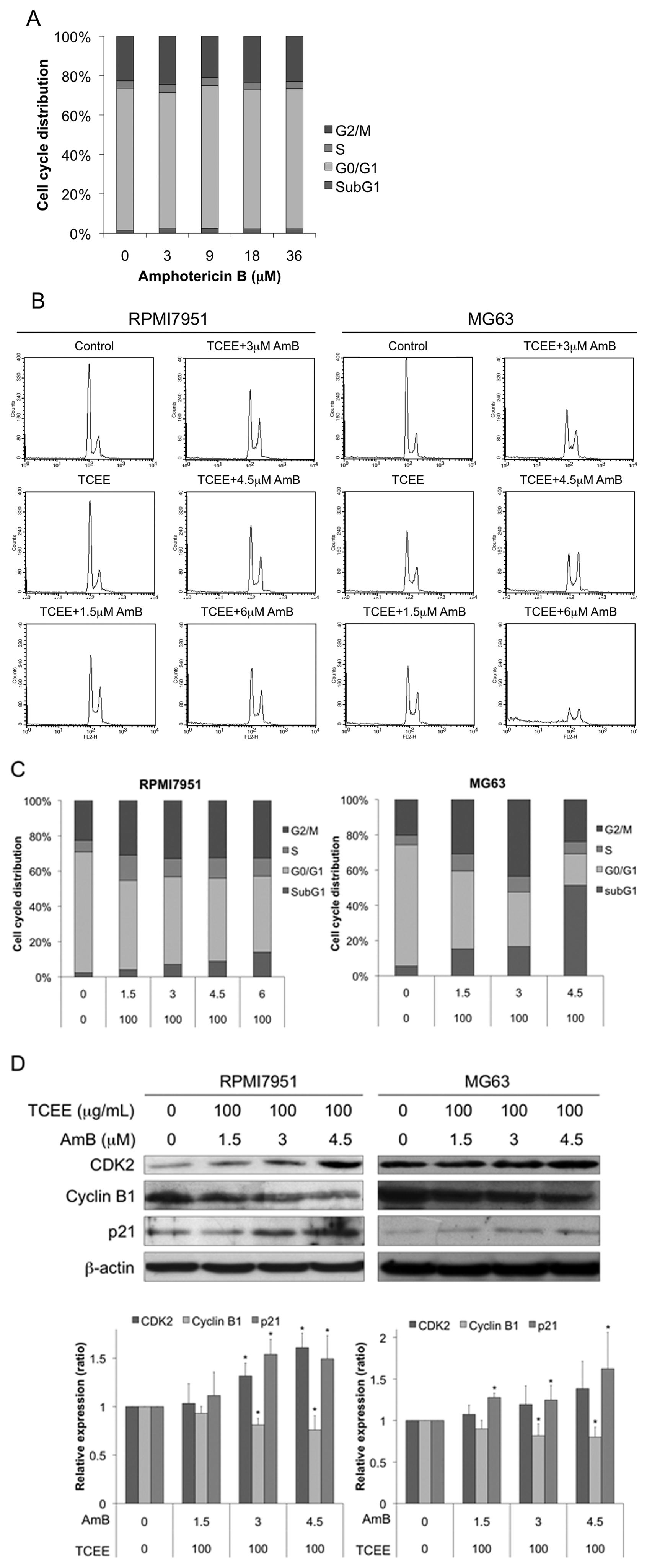
Induction of G2/M arrest in RPMI7951 and MG63 cells that were treated with TCEE and AmB
We investigated the mechanisms that were involved in G2/M arrest. We detected the alterations of key G2/M cell-cycle regulatory proteins and found that the cyclin B1 level was reduced and CDK2 and p21 levels were increased in cells that were treated with TCEE and AmB (Figure 5D). Therefore, our results show that the cell cycle arrest that is induced by the sequential treatment of TCEE and AmB may be attributed to their inhibitory effects on cell proliferation. Collectively, our results suggest that cytotoxicity that is induced by a combination of TCEE and AmB is partially mediated by perturbations in the regulation of the CDK2/cyclin B1 complex and leads to G2/M arrest.
Discussion
Chemotherapy with multiple drugs is commonly used in cancer therapy. The optimal combination of the drugs may be attributed to decreased drug toxicity, reduced drug resistance, and increased drug efficacy. The present study showed that RPMI7951 and MG63 cancer cells that were sequentially treated with TCEE and AmB significantly inhibited cell proliferation in a dose-dependent manner consistent with that previously reported on HT-29 cancer cell line. 24 In addition, TCEE enhanced the effect of AmB, which was indicated by reduced inhibitory effects in cancer cells that were treated with each drug alone. Nevertheless, the enhanced effects were only detected if the combined treatment was conducted in a sequential manner. Previous studies have suggested that in addition to the ergosterol that is contained in the fungi membranes, AmB interacts with the cholesterol in mammalian cells, especially low-density lipoproteins. 25 A previous study has showed that AmB has a strong affinity for 7-dehydrocholesterol (7-DHC) and suggested that AmB may be used as a therapeutic agent against some cancers such as hepatoma. 26 This hypothesis is based on studies that have investigated the selective substitution of cholesterol with one of its biosynthetic precursors (eg, 7-DHC) in hepatoma or a combination of a high-cholesterol diet and specific inhibitors of cholesterol biosynthesis.23,27 Although the extract of T camphoratus contains sterols with ergostane or lanostane skeletons, it is likely that TCEE pretreatment in cancer cells increases the susceptibility of the plasma membrane to AmB by the substitution or insertion of ergostane triterpenoids or lanostane triterpenoids. Using the proposed mechanism, TCEE potentiates the anticancer effects of AmB.
Previous studies have revealed that the cytotoxicity that is mediated by the mitochondrial apoptotic pathway is the major mechanism for TCEE or AmB activities.16,19,28 However, the absence of AmB-induced apoptosis in these cancer cell lines might be an indication of how effective was the activation by AmB of cell-specific survival factors as reported. 29 Therefore, the pretreatment of TCEE might possibly present not only ergosterol-like triterpenoids but also an inhibitor of AmB activated cell-specific survival factors leading to a significant increase in the apoptotic index at a lower AmB concentration. In addition to AmB, the extract of T camphoratus exhibits adjuvant antitumor effects when combining with traditional chemotherapeutic agents, such as mitomycin C, methotrexate, 29 and cisplatin. 30 Furthermore, as reported in the review by Cohen, 7 an increased intracellular calcium concentration by TCEE in cancer cells associated with the formation of AmBaq-pores might be another potential possibility, which leads to the abrupt decrease of the concentration threshold for the increased apoptotic and potentially synergistic effects by AmB that are observed in this study. Nevertheless, we discovered that the cancer cells that were treated with the drug combination exhibited phosphatydylserine externalization and loss of mithochondrial membrane potential in our study. Notably, the sublethal concentration of AmB (3 µM) that was applied in the experiments showed enhanced anticancer effects. Therefore, the addition of various concentrations of TCEE significantly triggered apoptosis. This nontoxic concentration of AmB in combination with TCEE may bypass AmB-induced nephrotoxicity, which limits AmB application.
We investigated other underlying mechanisms that mediate cytotoxicity following the combination treatment of TCEE and AmB. We found that the combination of AmB and TCEE arrested the cell cycle at the G2/M phase. Cell cycle progression is modulated by a complex of regulatory molecules, such as cyclin-dependent protein kinases (CDKs), cyclin-dependent protein kinase inhibitors (CDKIs), and cyclins. 31 The Cdc2 protein (also known as CDK1) and cyclin B1 is a complex that is essential for the cell cycle to progress from the G2 phase to the M phase. However, CDKIs, such as p21 and p27, inhibit the cell cycle. 32 Our data revealed that treatment with TCEE plus AmB simultaneously induced G2/M arrest and apoptosis. This result indicates that the main cytotoxic mechanisms of AmB plus TCEE are the arrest of the cell cycle and the induction of apoptosis. We examined the levels of key regulatory proteins of the G2/M phase and apoptosis to elucidate the mechanisms of G2/M arrest and apoptosis following the combination treatment. Elevated p21 and Bax levels were observed in RPMI7951 and MG63 cells that were treated with a combination of TCEE and AmB. Therefore, the combination treatment with the drugs produced an enhanced effect, resulting in a larger alteration of protein levels.
In conclusion, AmB and TCEE enhanced cell cycle arrest and apoptosis. The antitumor efficacy may be mediated by alterations in the levels of key proteins that regulate the cell cycle and apoptosis. Our results indicate the novel possibility that the drug combination of a conventional antifungal agent and a traditional Chinese medicine may serve as an alternative medicine for patients suffering from malignancies.
Footnotes
Acknowledgements
We thank Li-Hsin Chen and Pei-Ming Chu for their help with the article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.