
Abstract
The present study aimed at evaluating the anticancer and radiosensitizing potential of juglone against a chemoresistant and radioresistant tumor (B16F1 melanoma) growing on C57BL/6J mice. Volume doubling time, growth delay, and median survival were used to assess the in vivo anticancer and radiosensitizing potential of juglone. In vitro radiosensitizing potential of juglone was studied using clonogenic, comet, and reactive oxygen species induction assays. Treatment of tumor-bearing mice with sublethal doses of juglone caused a dose-dependent inhibition of tumor growth as evident from the growth delay and median survival values. Comet assay using tumor tissue and blood showed differential toxicity of juglone, where higher levels of DNA damage was seen in tumor tissue compared with blood cells. Pretreatment of tumor-bearing mice with optimum dose of juglone before radiation resulted in significant tumor growth inhibition compared with radiation alone. From the clonogenic assay, the authors observed a sensitization enhancement ratio of 1.37 for the combination treatment compared with radiation alone. Furthermore, comet assay studies revealed the potential of juglone to enhance the radiation-induced DNA damage and cause a delay in its repair. Juglone pretreatment before radiation also resulted in a significant elevation in the intracellular reactive oxygen species levels compared with radiation alone. In conclusion, the results of this study show the potential of juglone to inhibit the growth of melanoma in vivo. The study also revealed the potential of juglone to augment the radiation-induced cell death of melanoma cells, which may be attributed to oxidative stress–mediated DNA damage and its delayed repair.
Introduction
As per the recent World Health Organization (WHO) report, cancer remains a major global health threat with an estimated worldwide death of 12 million by the year 2020. The effectiveness of radiation in combating cancer was established more than 100 years ago, and it still continues to be one of the mainstays of cancer therapy with approximately half of all cancer patients receiving radiotherapy in some form during the course of their illness.1,2 Chemotherapy, on the other hand, has developed more recently, and since then extensive research efforts in the field of drug discovery and development have yielded a respectable armamentarium of potent anticancer agents. 3 Nevertheless, the treatment of most human solid tumors remains largely palliative, 4 and thus the need for more effective antineoplastic agents still remains. Intrinsic tumor resistance and normal tissue toxicity appear to be 2 major biological determinants in the failure of chemotherapy as well as radiotherapy. It is now generally believed that the effective treatment of most solid tumors is less likely to be achieved with 1 “magic bullet” and that a combination approach involving second-generation chemotherapeutics along with other treatment modalities may be required to attain better therapeutic outcome. 5 One of the strategies that has been proposed to improve the therapeutic outcome in cancer is to manipulate radiation-induced cell kill by combining a radiosensitizer or a chemotherapeutic agent along with radiotherapy. Many chemotherapeutic drugs have been combined with radiation with varying degrees of success, and many more are still under clinical evaluation. 6 Nevertheless, there is still a need to look for novel chemotherapeutic agents that could effectively be combined with radiation to improve the cancer therapeutic outcome.
Exploring the natural resources for the development of new anticancer agents has proven to be extremely beneficial. It is estimated that about 67% of the pharmaceuticals approved for cancer therapy during the period between 1974 and 1994 were derived from natural sources. 7 Quinones comprise a broad class of phenolic compounds that exhibit an array of pharmacological properties and are widely distributed in nature. Plants that are rich in quinones have been extensively used in traditional folk medicine for the treatment of various ailments, including cancer. 8 These compounds are of great interest to researchers, because of the fact that several important anticancer drugs such as doxorubicin, daunorubicin, mitomycin C, and so on contain the quinone nucleus. Many other natural compounds with quinone nucleus such as β-lapachone, plumbagin, and so on have also shown promising anticancer activity both in vitro and in vivo.4,9 Besides, several groups including ours have shown plumbagin to possess radiosensitizing potential as well.10-12
Walnut (Juglans regia L) belonging to the family Juglandaceae has a long history of use in traditional folk medicine for the treatment of various ailments. Different parts of the tree are reported to possess antimicrobial, anti-inflammatory, immunomodulatory, diuretic, laxative, and anticancer properties.13-15 The aqueous extracts of walnut are also reported to possess strong antioxidant properties.13,16 Herbal preparations of walnut are known to suppress the growth of spontaneous mammary adenocarcinoma in Swiss albino mice. 17 The major active ingredients of walnut include naphthoquinones such as juglone, α-hydrojuglone and β-hydrojuglone alongside others such as vitamin C, caffeic acid, ellagic acid, hyperin, and kaempferol. Juglone (5-hydroxy-1,4 naphthoquinone), a structural analogue of plumbagin is reported to possess potent cytotoxic properties in vitro against human cancer cell lines, including human leukemia (HL-60) cells, doxorubicin-resistant human leukemia (HL-60R) cells, human lung carcinoma (A549), and human gastric cells (SGC-7901).18-20 In an earlier study, we reported the effect of juglone on B16F1 melanoma cells in vitro and concluded that the cytotoxic potential of juglone was mediated by multifactorial mechanisms involving oxidative stress, cell membrane damage, and clastogenic effect ultimately leading to cell death by both apoptosis and necrosis. 21 In the present study, we evaluated the in vivo anticancer potential of juglone against a chemoresistant and radioresistant B16F1 melanoma solid tumor model. To our knowledge, there are no previous reports about the radiosensitizing potential of juglone. Therefore, the present research work was also aimed at evaluating the in vitro and in vivo radiosensitizing property of juglone against B16F1 melanoma cells.
Materials and Method
Chemicals and Reagents
Juglone, ethidium bromide, agarose (normal melting as well as low melting), ethylenediamine tetraacetic acid (EDTA), Eagle’s minimum essential medium (MEM),
Cell Lines and Cultures
B16F1 melanoma cells procured from the National Centre for Cell Sciences (Pune, India) were routinely grown in 25-cm2 flasks containing MEM supplemented with 10% fetal bovine serum, 1%
Preparation of Juglone Solutions
For in vitro studies, juglone was dissolved in absolute alcohol at a stock concentration of 10 mM and stored at −80°C. Working concentrations were prepared from the stock aliquots by serially diluting with MEM to the required concentrations. The concentration of juglone for the in vitro studies was chosen based on our earlier studies. 21 For in vivo studies, juglone was dissolved in minimum amount of alcohol and made up to the required concentrations with 20% PEG in saline (vehicle).
Animals and Tumor Model
C57BL/6J mice, procured from the National Institute of Nutrition (Hyderabad, India), were maintained under controlled conditions of temperature (23 ± 2°C), humidity (50 ± 5%), light (14 hours and 10 hours light and dark cycles, respectively) and were allowed access to sterile food and water ad libitum. The animal care and handling were according to the guidelines of the WHO (Geneva, Switzerland) and the Indian National Science Academy (INSA; New Delhi, India). For experimental purposes, 8-week-old to 12-week-old mice of either sex, weighing 20 to 25 g, from an inbred colony were selected. All the animal experiments were carried out with the prior approval from the institutional animal ethics committee (Kasturba Medical College, Manipal, India). Solid tumors were grown on the dorsal side of the mice by an intradermal injection of 5 × 105 viable cells.
Irradiation
All radiation studies were carried out using a 60Co gamma teletherapy facility (Theratron Atomic Energy Agency, Canada) at the Shirdi Saibaba Cancer Hospital (Manipal, India). The radiation doses used in the present investigation were based on our earlier studies (unpublished observation).
For in vitro experiments, exponentially growing cells with or without juglone treatment were exposed to gamma radiation at a dose rate of 1 Gy/min and source to surface distance (SSD) of 73 cm. For in vivo experiments, local tumor irradiation was carried out at a dose rate of 1.33 Gy/min in a field size of 4 in. × 4 in. (10.1 cm × 10.1 cm). Tumor-bearing animals with or without drug treatment were anesthetized (50 mg/kg body weight (BW) of ketamine and 8 mg/kg BW of diazepam), immobilized in a specially designed, well-ventilated perspex box in such a way that only the tumors protrude out into the radiation field. All the other parts of the body were shielded with 4 half-value layer (HVL) of lead (5 cm thick) kept in a shielding tray fitted to the collimator (Figure 1).

In vivo irradiation setup for the evaluation of radiosensitization potential of juglone
Assessment of In Vivo Anticancer Potential of Juglone
Acute toxicity studies of juglone
The acute toxicity study of juglone was performed as described previously. 22 Briefly, animals (fasted for 18 hours prior to juglone treatment) were divided into several groups of 10 each. Each group of animals was injected intravenously with different doses of juglone (prepared freshly as mentioned previously) namely, 0.5, 1, 3, 6, and 10 mg/kg BW. Immediately after the treatment, animals were provided with food and water, and the animals were monitored daily up to 14 days for mortality. Median lethal dose or LD50(14) was then calculated using probit analysis as described by Litchfield and Wilcoxon. 23
Selection of optimum dose for antitumor activity
Tumors were induced in the mice by injecting 5 × 105 viable cells intradermally on the dorsal side. When the tumors became palpable, the tumor diameter in 3 perpendicular directions was measured using a vernier caliper, and the tumor volume was calculated as described previously 24 using the following formula:

where D1, D2, and D3 are the tumor diameters in 3 perpendicular planes. Once the tumor size reached 100 ± 10 mm3, the animals were divided into the following groups of 10 each:
Group 1: Tumor-bearing animals in this group received 100 µL of vehicle (vehicle-treated control group)
Groups 2 to 4: Animals in this group were injected intravenously with different doses of juglone, namely, 0.5, 1, and 1.5 mg/kg BW (prepared freshly under sterile conditions) as single administration.
The best optimum dose from this study was used to evaluate the anticancer activity of juglone administered as multiple doses. Once the tumor size reached 100 ± 10 mm3, the tumor-bearing animals were divided into the following groups of 10 animals each:
Group 1: Animals in this group received 100 µL of vehicle (vehicle-treated control group)
Group 2: Animals in this group received repeated injections of the optimum dose of juglone dissolved in 100 µL of vehicle on days 1, 3, and 5 consecutively.
Assessment of Tumor Response
The tumor diameters were measured every alternate day using a vernier caliper, and the tumor volume was calculated and plotted against days posttreatment. The tumor response after various treatments was then assessed using (a) volume doubling time (VDT), which is the time required for the tumors to reach double the initial volume and (b) growth delay (GD), which is the difference in time between the treated and untreated tumors to reach 5 times the initial volume. The animals were further monitored for survival, and the median survival times were determined from the Kaplan–Meier analysis of survival. During the course of experimentation, animals that were under stress due to excess tumor burden were removed from the study (because of humane considerations) and were considered as censored in the survival analysis.
Alkaline comet assay
Tumors were induced in the mice by injecting 5 × 105 viable cells intradermally. Once the tumor size reached 100 ± 10 mm3, the animals were divided into the following groups of 12 each.
Group 1: Animals in this group received 100 µL of vehicle
Group 2: Animals in this group were injected intravenously with optimum anticancer dose of juglone (1 mg/kg BW)
A subgroup of 4 animals from the above treatment groups were euthanized on days 1, 7, and 15 posttreatment. At the indicated time intervals, the blood samples were collected in heparinized tubes by retro-orbital plexus. The tumors from these animals were also excised immediately, and single-cell suspension was prepared in ice-cold PBS as described elsewhere. 25 The differential genotoxic efficacy of juglone among the blood and tumor samples was then studied using the alkaline version of single-cell gel electrophoresis (comet) assay as described previously. 26 Briefly, microscopic slides were coated with 1.5% normal melting agarose and left overnight to dry. Cell suspensions (tumor as well as blood) were mixed with 190 µL of 0.75% low-melting-point agarose (LMPA) and were distributed on the coated slide. The slides were left to gel for 10 minutes at 4°C, before a third layer of 200 µL 0.75% LMPA was added to the slide and left for 10 minutes at 4°C. The slides were then dipped in cold lysing solution (2.5 M NaCl, 100 mM EDTA, 10 mM Tris base, and 0.2 M NaOH, 1% Triton X, 10% DMSO; pH 10) for at least 2 hours at 4°C. The slides were then transferred to an electrophoresis unit containing electrophoresis buffer (300 mM NaOH, 1 mM EDTA; pH ≥12.3), and were left immersed in the solution for 20 minutes to allow DNA unwinding before being subjected to electrophoresis. Electrophoresis was carried out for 20 minutes at a voltage of 0.7 V/cm and a current of 300 mA. Next, the slides were rinsed with neutralization buffer (0.4 M Tris; pH 7.4) and stained with ethidium bromide. The stained cells were then observed and photographed at 20× magnification using a fluorescence microscope (Olympus BX51, Olympus Microscopes, Tokyo, Japan) equipped with a 515- to 535-nm excitation filter, a 590-nm barrier filter, and a CCD camera (Cool SNAP-Procf Digital Color Camera Kit version 4.1, Media Cybergenetics, Silver Spring, MD). Images of a minimum of 100 cells per treatment were analyzed using the Komet software (version 5.5, Kinetic Imaging Ltd, Bromborough, UK). The mean olive tail moment (OTM) was used as a parameter to assess the level of DNA damage.
Assessment of Radiosensitizing Potential of Juglone
In Vivo Studies
Tumors were induced in the mice by injecting 5 × 105 viable cells intradermally. Once the tumor size reached 100 ± 10 mm3, the animals were divided into the following groups of 10 each:
Group 1: Animals in this group received 100 µL of vehicle
Groups 2 and 3: Animals in this group received local tumor irradiation with different doses of gamma radiation (10 and 30 Gy),
Groups 4 and 5: Animals in this group received the optimum dose of juglone followed by local tumor irradiation with 10 and 30 Gy dose of gamma radiation, respectively.
The tumor growth parameters after various treatments were then studied as described previously.
In Vitro Studies
Clonogenic assay
Cell survival following gamma radiation with or without juglone treatment was evaluated using clonogenic assay. 27 Exponentially growing B16 melanoma cells were treated with 5 µM juglone for 1 hour followed by different doses of γ-radiation (1, 3, 6, and 8 Gy). Cells were then trypsinized and appropriate numbers of viable cells were plated in 60-mm2 petri dishes (in triplicate) and were left undisturbed for 10 to 12 days. Thereafter, colonies containing at least 50 cells were counted as clonogenic survivors. All colony counts were corrected for plating efficiency (PE) to yield survival values of 100% for untreated controls, and the surviving fractions were calculated and plotted using linear quadratic model as mentioned previously. 28 Sensitizing efficiency of juglone was then expressed in terms of SF2 value (defined as the survival fraction of exponentially growing cells at 2 Gy radiation dose) and the sensitizer enhancement ratio (SER; defined as the ratio of D0 untreated cells/D0 treated cells, where D0 value represents the radiation dose that leads to 37% survival). To evaluate the nature of the interaction between juglone and gamma irradiation, the combination index (CI) was calculated according to the method of Chou and Talalay 29 as described elsewhere 30 using the following formula:
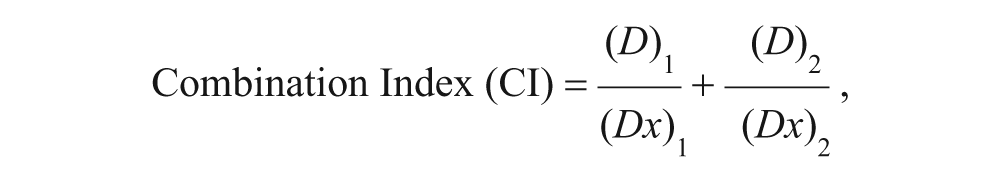
where (Dx)1 and (Dx)2 in the denominators are the doses (or concentrations) of juglone and radiation alone that give x% inhibition, whereas (D)1 and (D)2 in the numerators are the doses of juglone and radiation in combination that also inhibits x% (iso effect). CI < 1, CI = 1, and CI > 1 indicate synergistic, additive, and antagonistic effects, respectively.
DNA damage and repair studies using comet assay
B16F1 melanoma cells were incubated with or without juglone (5 µM) for 1 hour before irradiation (6 Gy) and during 3 and 6 hours posttreatment repair incubations. At the end of incubation times, DNA strand breaks were quantified using the alkaline comet assay 26 as described earlier.
Analysis of the intracellular reactive oxygen species (ROS) levels
Effect of various treatments on the intracellular ROS levels was studied using a fluorescent probe 2′, 7′-dichlorofluorescin diacetate (DCFH-DA). Briefly, cells were seeded in a 6-well plate at a density of 2 × 105/well. Twenty-four hours later, the cells were incubated with 10 µM concentrations of DCFH-DA for 30 minutes at room temperature in the dark. The unincorporated fluorescence probe was removed from the cells by 2 washes with PBS. The DCFH-DA-loaded cells were then treated with juglone alone (5 µM) or radiation alone (6 Gy), or in combination. At the end of various treatments, the cells were trypsinized, washed, resuspended in appropriate volume of PBS and transferred to 96-well black plates. The fluorescence intensity of the cells was then measured using a microplate fluorescence reader (Infinite M200, Tecan Group Ltd, Männedorf, Switzerland) at excitation and emission wavelengths of 488 and 525 nm, respectively.
Statistical analysis
The experimental data were expressed as mean ± standard error of the mean. The plating efficiency of the cells was determined, and the data points of the surviving fraction was fitted on to a linear quadratic model, SF (surviving fraction) = exp[−(αD + βD2)]. Survival studies were performed by Kaplan–Meier survival analysis using GraphPad Prism version 3.00 (San Diego, CA), and the median survival time was reported. For all other studies, one-way analysis of variance followed by Bonferroni’s post hoc test was used to compare the significance between various treatments.
Results
In Vivo Anticancer Potential of Juglone
Acute toxicity studies in mice
Acute toxicity studies were carried out in C57 mice by injecting different doses of juglone intravenously (Figure 2). We observed a dose-dependent increase in the animal mortality with animal death occurring within 48 hours of drug administration. Animals injected with 0.5 and 1 mg/kg BW juglone did not show any signs of toxicity, and no mortalities were seen during the 14 days of observation period. However, in animals injected with 3 mg/kg BW juglone, we observed 20% mortality along with toxic symptoms such as reduced activity, anorexia, and so on. Administration of 6 and 10 mg/kg BW juglone resulted in 60% and 100% mortality, respectively. The LD50(14) value for juglone was calculated using the probit method and found to be 4.2 mg/kg BW when administered intravenously.

Acute toxicity studies for intravenously administered juglone using probit method of analysis
Optimum dose selection for anticancer activity
Tumor-bearing animals were injected with single sublethal doses of juglone, namely, 0.5, 1, and 1.5 mg/kg BW, and the tumor growth kinetics were monitored and recorded as shown in Figure 3 and Table 1. The tumor size in the vehicle-treated mice increased steeply (approximately 66-fold) and reached an average size of 6600 mm3 within 14 days. On the other hand, juglone treatment resulted in inhibition of tumor growth in a dose-dependent fashion. Although the tumor size in mice treated with 0.5 mg/kg BW juglone reduced only marginally, the tumor size in mice treated with 1 and 1.5 mg/kg BW reduced significantly (P < .001) on day 14 compared with vehicle-treated controls (Figure 3, Table 1). However, there was no significant reduction in the tumor volume when the dose of juglone was increased from 1 to 1.5 mg/kg BW. Animal survival analysis revealed that the treatment of tumor-bearing mice with 0.5, 1, and 1.5 mg/kg BW juglone resulted in a median survival of 22, 26, and 27 days, respectively, compared with 19 days in vehicle-treated group. Based on these studies, 1 mg/kg BW juglone was considered the optimum dose for further studies. Studies were also carried out to determine the effect of repeated dose juglone treatment on the tumor growth kinetics, and the results are shown in Figure 3 (inset). Treatment of mice with repeated doses of juglone on days 1, 3, and 5 resulted in a significant (P < .01) improvement in antitumor activity compared with single dose administration.
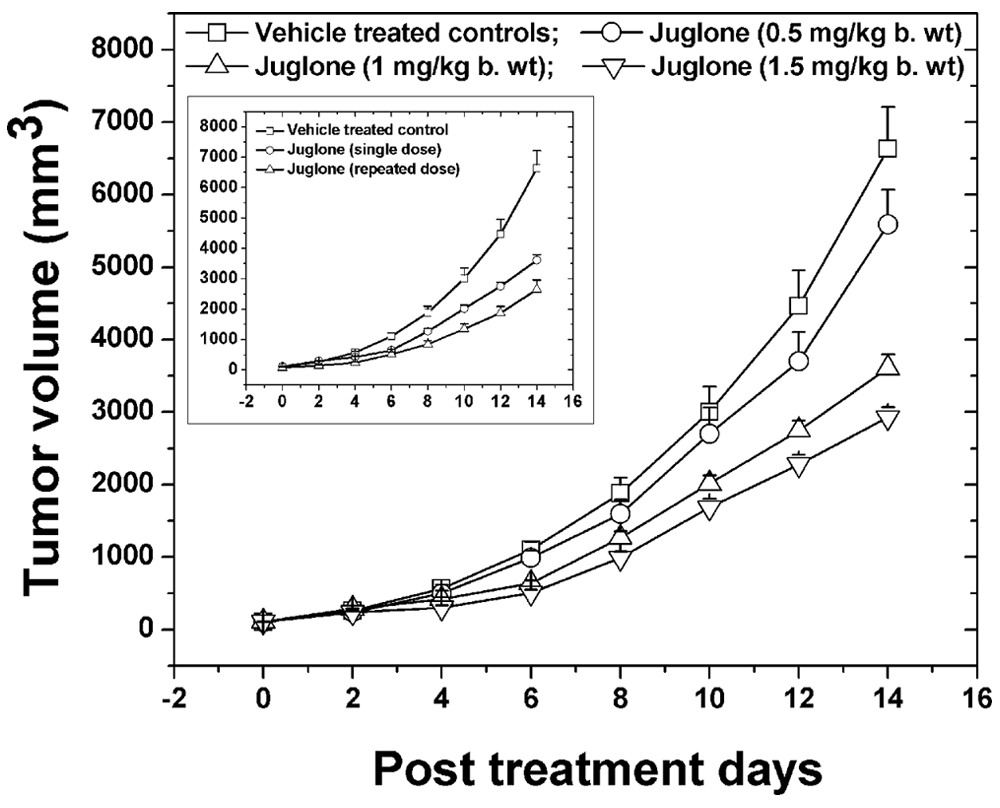
In vivo optimum dose selection studies for juglone administered as single dose via intravenous route of administration
Tumor Growth Kinetics and Survival Studies for Optimum Dose Selection Studies of Juglone In Vivo

Abbreviations: BW, body weight; VDT, volume doubling time (time required for the tumors to reach double the initial volume); SE, standard error; 5×, time required for the tumors to reach 5 times the initial volume; GD, growth delay (the difference in time between the treated and untreated tumors to reach 5 times the initial volume).
P < .001, compared with vehicle-treated controls.
Assessment of DNA damage in vivo
The effect of juglone treatment on the DNA damage levels in the tumors of mice was studied (Figure 4A). At different intervals of time after treatment, the tumors were excised and processed immediately for comet assay. We observed a significant elevation in the OTM levels (P < .01) in the tumor tissue 1 day after juglone treatment compared with corresponding vehicle-treated controls. On day 7 posttreatment, the damage largely got repaired with OTM values remaining only slightly higher than its vehicle-treated counterpart. However, on day 15 following juglone treatment, the OTM levels reached the baseline levels.

Effect of juglone (1 mg/kg body weight [BW]) treatment on DNA damage levels
The same sets of animals were used to study the differential genotoxic effect by measuring the DNA damage levels (OTM) in the peripheral blood (as an index of DNA damage in lymphocytes). The results of this study are depicted in Figure 4B. On day 1 posttreatment, we observed a significant increase (P < .05) in the OTM levels of animals treated with juglone compared with the respective controls. However, on day 7 and thereafter, these levels got back to normal levels seen in the control animals.
Radiosensitizing Potential of Juglone In Vivo
The optimum dose of juglone (1 mg/kg BW) from the previous study was then combined with radiation (10 and 30 Gy) to evaluate its radiosensitizing potential in vivo. Tumor response was measured in terms of VDT and GD as mentioned in the materials and methods section. The results of this study are depicted in Figure 5 and Table 2. We observed a significant increase in the VDT as well as tumor GD with combination therapy (juglone plus radiation) in comparison with radiation-alone group at both 10 Gy (P < .05) and 30 Gy (P < .001) radiation doses. Analysis of the survival patterns (Table 2) revealed that the treatment of tumor-bearing mice with 10 and 30 Gy radiation resulted in median survival values of 31 and 40 days, respectively. In addition, treatment of tumor-bearing mice with juglone before local tumor irradiation with 10 and 30 Gy caused further increase in the median survival to 34 and 46 days, respectively.

Radiosensitizing potential of juglone in vivo against B16F1 melanoma cells
Tumor Growth Kinetics and Survival Studies for Radiosensitizing Potential of Juglone In Vivo

Abbreviations: BW, body weight; VDT, volume doubling time (time required for the tumors to reach double the initial volume); SE, standard error; 5×, time required for the tumors to reach 5 times the initial volume; GD, growth delay (the difference in time between the treated and untreated tumors to reach 5 times the initial volume).
P < .05, compared with respective radiation alone groups.
P < .001, compared with respective radiation alone groups.
Radiosensitizing Potential of Juglone In Vitro
Juglone increases radiation sensitivity of melanoma cells
The effect of juglone on the radiation survival curves was studied using colony-forming assay. Cells were pretreated with 5 µM juglone (selected based on our previous studies) for 1 hour before treating with different doses of gamma radiation (1-8 Gy). As shown in Figure 6, treatment of melanoma cells with juglone prior to exposure to different doses of radiation resulted in a clear dose-dependent decrease in radiation-induced cell survival (SF2 = 0.303, D0 value = 1.16 Gy) compared with radiation alone group (SF2 = 0.545; D0 value = 1.59 Gy) suggesting the potential of juglone to increase the radiosensitivity of melanoma cells. The SER calculated based on the D0 value was found to be 1.37.

Radiosensitizing potential of juglone in vitro against B16F1 melanoma cells assessed using clonogenic assay
Juglone enhances radiation-induced DNA damage
The effect of juglone on the radiation-induced DNA damage in melanoma cells was determined using alkaline comet assay. Melanoma cells treated with 5 µM juglone alone or radiation (6 Gy) alone showed a 5.14-fold and 2.19-fold increase in the OTM (indicative of DNA damage), respectively, in comparison with untreated cells (Figure 7). Furthermore, treatment of cells with juglone before irradiation showed significantly higher OTM levels compared with independent treatments (P < .05 vs juglone alone; P < .01 vs radiation alone) as well as the control (P < .01).

Radiosensitization potential of juglone against melanoma cells in vitro assessed using alkaline comet assay
Juglone slows down the repair of radiation-induced DNA damage
To study if juglone has any effect on radiation-induced DNA damage repair, we performed the alkaline comet assay after incubating the cells with fresh medium for 0, 3, and 6 hours after various treatments. The results of this study are depicted in Figure 7. From these results, it can be seen that the cells treated with radiation alone repaired almost completely within 3 hours of incubation. Furthermore, cells treated with juglone alone showed significant reduction in OTM levels (indicative of DNA repair) at 3 hours compared with the respective 0-hour interval. However, near-complete repair was only seen at 6 hours incubation time. On the other hand, cells treated with a combination of juglone followed by radiation showed a remarkable slow down in the DNA damage repair kinetics with mean OTM values hovering around 21.5% (4-fold higher than control values) even at 6 hours incubation time.
Elevated levels of intracellular ROS by a combination of juglone and radiation
Recently, we demonstrated the potential of juglone to induce ROS in melanoma cells. 21 To investigate the relationship between ROS generation and the observed radiosensitizing effect of juglone, we employed an ROS-sensitive dye DCFH-DA to detect ROS production fluorimetrically. From the results of this study (Figure 8), treatment of melanoma cells with either juglone (5 µM) or radiation (6 Gy) resulted in 3.2-fold and 2.9-fold increase in the levels of intracellular ROS, respectively, compared with untreated control. Furthermore, a combination treatment of juglone followed by radiation resulted in 5.26-fold increase in ROS levels compared with untreated controls. The increase in the levels of intracellular ROS in the combination treatment was statistically significant (P < .001) in comparison with individual treatments as well as untreated controls.

Effect of juglone, radiation, and combination treatment on the intracellular reactive oxygen species (ROS) levels in melanoma cells assessed using DCFH-DA assay
Discussion
Intrinsic chemoresistance/radioresistance as well as the presence of hypoxic cells make the effective treatment of most solid tumors a daunting task. To improve the therapeutic outcome in cancer, several attempts have been made to combine a chemical agent/radiosensitizer with radiotherapy. However, clinical usefulness of most sensitizers remains ineffective and controversial. 2 As a result, efforts are now being directed at combining chemotherapeutic agents with radiation to improve the therapeutic outcome. In this regard, various chemotherapeutic agents such as cisplatin, gemcitabine, paclitaxel, docetaxel, irinotecan, and so on have been combined with radiation with varying degrees of success.6,31 However, chemoradiotherapy with conventional chemotherapeutic agents is associated with toxicity, 32 which necessitates the search for novel anticancer compounds with radiosensitizing properties.
Owing to their ease of availability, cost-effective nature, and safer toxicity profiles, screening of plant and other natural products has gained considerable interest in modern drug discovery programs. 33 Walnut has been used in traditional folk medicines for the treatment of cancer. In the present study, we investigated the in vivo anticancer and radiosensitizing potential of the main active constituent of walnut, juglone, against B16F1 melanoma cells. The acute toxicity studies carried out using the probit analysis revealed that juglone administered by intravenous route had an LD50 value of about 4.2 mg/kg BW. To our knowledge, there are no previous reports of any toxicity studies of juglone administered via intravenous route in mouse. However, Westfall et al 34 reported an LD50 value of 2.5 mg/kg BW in mice when administered through the oral route. Registry of Toxic Effects of Chemical Substances (RTECS, National Library of Medicine) reported an LD50 value of 25 mg/kg BW for juglone in mice administered via intraperitoneal route. In another report, administration of juglone in dogs at a dose of 5 mg/kg BW intravenously was found to cause slight hemorrhages in the lungs with no direct effect on cardiovascular system. 35 From our studies, we observed slight toxic symptoms such as weight loss and reduced activity when juglone was injected at a dose of 3 mg/kg BW. However, no toxic effects were seen in animals treated with lower doses of juglone (up to 1.5 mg/kg BW).
Okada et al 36 studied the effect of juglone in vivo against Ehrlich ascites tumor cells transplanted in Swiss mice and reported that juglone induced mitotic abnormalities such as decreased mitotic figures with concomitant metaphase arrest, indicating its therapeutic potential. Similar observation was also made by Sugie et al, 37 wherein male F344 rats administered with juglone had lower incidence and multiplicity of azoxymethane-induced intestinal tumors. On the contrary, Monks et al 38 reported that juglone promoted the 7,12-dimethylbenz[a]anthracene (DMBA)-initiated skin carcinomas in SENCAR mice when applied topically for 40 weeks. In another study, 39 juglone treatment promoted the skin tumors in female ICR/Ha Swiss mice pretreated with DMBA. However, treatment of these mice with juglone alone (without DMBA pretreatment) did not induce any tumors in these mice. This shows that there is still ambiguity regarding the in vivo anticancer potential of juglone, which formed the basis of the present investigation. In the present study, based on the acute toxicity data, sublethal doses of juglone were chosen for evaluation of its in vivo anticancer activity. Treatment of tumor-bearing mice with juglone (1 mg/kg BW) resulted in significant inhibition (P < .001) in tumor growth parameters as evidenced by the 5×, GD, and median survival values. Results from the present investigation indicate the potential of juglone as an anticancer agent, in agreement with the earlier findings.36,37
Reactive oxygen species are generally very small, highly reactive molecules that can be generated as by-products of cellular metabolism. The overproduction of ROS can cause damage to various biomolecules, including lipids, DNA, and proteins, ultimately leading to cell death. It is well documented that increased generation of ROS leading to an altered redox homeostasis is a common feature in most cancer forms. 40 Taking that into consideration, it has been hypothesized that pro-oxidant compounds that induce ROS in the cells result in the dysregulation of already altered ROS levels in cancer cells leading to their loss of viability. In contrast, untransformed normal cells having a normal redox status are believed to be more resistant to the ROS insults. This diversity in the biochemical property of cancer and normal cells are now being exploited for their possible beneficial role in cancer therapy. 41 To that effect, numerous studies have been carried out to evaluate the preferential sensitivity of cancer cells to small molecule pro-oxidants such as quinone-based electrophiles, 9 redox cyclers, 42 and organic endoperoxides. 43 Juglone, a small molecule pro-oxidant compound belonging to the quinone class, induces apoptotic and necrotic cell death in cancer cells essentially by oxidative stress–mediated mechanisms.19,21 Based on the existing literature as well as from our previous studies, the observed anticancer potential of juglone in vivo may be attributed at least in part to its ROS-generating ability.
Reactive oxygen species are known to induce DNA damage by causing single-strand breaks, double-strand breaks, and so on. The alkaline version of the single-cell gel electrophoresis (comet) assay is a versatile method to detect DNA single-strand breaks, double-strand breaks, or cross-links in virtually any cell type. 44 To study if the observed tumor growth inhibition could be a result of DNA damage induced by juglone, we measured the DNA damage levels in the tumor tissue. Juglone treatment caused a significant (P < .01) elevation in the DNA damage levels of tumor tissue on day 1 posttreatment. On day 7 posttreatment, the OTM levels were only slightly higher than the control levels indicating repair of the damaged DNA. Thereafter, the OTM levels reached normal values. Tirapazamine, a well-known chemotherapeutic agent was also shown to induce DNA damage in the tumor cells in vivo, and this DNA damage in the tumor tissue was reported to be closely associated with the tumor cell kill. 45 From the present study, it is clear that the DNA damage induced by juglone in the tumor tissue could have resulted in the tumor cell kill ultimately resulting in the observed delay in tumor growth parameters.
Conversely, many chemotherapeutic agents are also reported to induce DNA damage in blood lymphocytes at their therapeutic doses. 46 Therefore, to evaluate the effect of juglone on the DNA damage levels in the blood cells, alkaline comet assay was performed using the blood cells of the animals treated with optimum dose of juglone. Results indicate that on day 1 posttreatment, juglone induced significant (P < .05) increase in the OTM levels, which, however, reversed back to normal levels on day 7 and thereon. The present results suggest that juglone also causes moderate levels of initial normal tissue toxicity, which, however, may be mitigated by formulating juglone using various drug-targeting strategies as was demonstrated for plumbagin in our earlier report. 47
As discussed earlier, a bimodality approach involving a combination of chemotherapeutic agent along with radiation may help in improving the therapeutic outcome in cancer. Many of the potent phytochemicals such as plumbagin, withaferin A, and so on have been shown to augment the radiation-induced cell killing against various cell lines in vitro.12,48 Furthermore, our earlier studies have also demonstrated the radiosensitizing potential of these plant-based compounds against various cancer models in vivo.10,24 To our knowledge, there are no earlier reports about the potential of juglone to modify the radiation-induced cell killing. In the present study, exposure of melanoma cells to juglone prior to treatment with different doses of γ-radiation resulted in a significant reduction in the surviving fraction of melanoma cells in comparison with the radiation-alone group (Figure 6). A sensitizer enhancement ratio of 1.37 was observed, which may be considered as significant levels of sensitization based on the earlier reports where SER value of >1.1 were considered significant. 49 Also, the analysis of the clonogenic assay data using the method of Chou and Talalay 29 gave a mean CI value of 0.99 suggesting that the interaction between juglone with gamma radiation was more of an additive nature and not synergistic. Several other clinically approved anticancer agents are also known to elicit an additive response in combination with radiation. 50 These findings further corroborate the results of our in vivo studies where treatment of mice with juglone before local tumor irradiation resulted in significant delay in the tumor growth parameters as compared with independent treatments (Table 2, Figure 5). These results clearly indicate the ability of juglone to enhance the radiosensitivity of melanoma cells both in vitro and in vivo.
Effective repair of the damaged DNA is a critical event that determines the radiosensitivity of cancer cells, and inhibition of DNA damage repair is proposed to be one of the key mechanisms in radiosensitization. β-Lapachone, a plant-based quinone with established anticancer properties, was shown to enhance the radiosensitivity of cancer cells by inhibition of DNA repair mechanisms. 51 In the present investigation, we studied the effect of juglone on the radiation-induced DNA damage and repair. At 0 time (immediately after various treatments), we observed significantly elevated OTM levels in the combination treatment compared with the individual treatments. Results from the repair studies shows significantly higher levels of OTM in combination treatment compared with the independent treatments even at 6 hours incubation. However, at the 24 hours incubation time point, the DNA damage levels in all the treatment groups reached baseline levels (data not shown). These results clearly indicate the potential of juglone to slow down the repair of radiation-induced DNA damage in melanoma cells, which may have resulted in the observed radiosensitizing effect of juglone.
It is a known fact that under aerobic conditions, radiation causes generation and accumulation of ROS such as superoxide anion, hydrogen peroxide, singlet oxygen, hydroxyl radical, and peroxyl radical, which could damage DNA as well as hinder its repair process. The intracellular antioxidant defense mechanisms normally help restore the elevated oxidative status in the cells. However, the presence of an agent that could deplete the cellular antioxidant defenses before subjecting to radiation could enhance the cell kill in cancer. We earlier reported the potential of juglone to deplete the intracellular reduced glutathione and induce oxidative stress in melanoma cells. 21 In the present study, combined treatment of melanoma cells with juglone followed by radiation resulted in significant increase in the intracellular ROS levels compared with the control as well as individual treatments, which may have resulted in the observed radiosensitization effect by enhancing DNA damage and slowing down its repair.
The toxicity of quinones is generally attributed to their ability to undergo redox cycling with consequent production of semiquinone radicals and ROS leading to oxidative stress–mediated cell death. Although the exact mechanism of radiosensitization potential of juglone is not clearly understood, the observed additive nature of its radiosensitizing potential against melanoma cells may be attributed to the ability of juglone to induce ROS-mediated DNA damage (comet assay) leading to cell death (as evidenced by clonogenic assay). Also, the presence of hydroxyl group on the position 5 of the naphthoquinone ring may have contributed to the observed ROS levels that resulted in the radiosensitizing effect of juglone. Similar findings were also reported in earlier studies where juglone with hydroxyl group in the fifth position was reported to considerably increase the toxicity against rat hepatocytes.52,53 However, the involvement of other mechanistic pathways in the radiosensitizing potential of juglone needs further investigation.
In conclusion, the present study has shown the potential of juglone to inhibit the growth of melanoma cells in vivo. Also, this study has for the first time shown the potential of juglone to enhance the radiation-induced cell kill in melanoma cells both in vitro and in vivo. In view of these findings, and the existing earlier reports of its use in traditional folk medicine, juglone deserves further studies on its toxicity aspects using a spectrum of preclinical models.
Footnotes
Acknowledgements
We would like to thank Dr M. S. Vidyasagar, Professor and Head, and Dr J. G. R. Solomon, Department of Radiotherapy and Oncology, Kasturba Hospital, Manipal, for providing the irradiation facility and for dosimetric calculations, respectively. The authors are also thankful to Dr K. Satyamoorthy, The Director, Manipal Life Sciences Centre, Manipal University, Manipal, for providing the facilities and encouragement during this study.
The author(s) declared no potential conflicts of interests with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article:
The financial support from Indian Council of Medical Research (ICMR), New Delhi, India (IRIS No. 2005-00150), to carry out this study is gratefully acknowledged.