
Abstract
Cerebrovascular changes occur in Alzheimer’s disease (AD). The progesterone receptor membrane component-1 (PGRMC1) is a well identified hormone receptor with multiple functions in AD. This study aims to explore the involvement of PGRMC1 in the regulation of vascular endothelial function, providing new therapy options for AD. Single-cell sequencing revealed that the expression of PGRMC1 is lower in AD. By bioinformatics analysis, we found PGRMC1 was associated with regulation of cell proliferation, angiogenesis and etc. To understand the functional significance of PGRMC1, knockdown and overexpression were performed using human brain microvascular endothelial cells (HBMVECs), respectively. Cell proliferation assay, migration assay, tube formation assay were performed in experiments. We demonstrated that the overexpression of PGRMC1 promoted the cellular processes associated with endothelia cell proliferation, migration, and angiogenesis, significantly. In conclusion, PGRMC1 may contribute to the modulation of HBMVECs function in AD. This finding may offer novel targets for AD treatment.
Keywords
Introduction
Alzheimer’s disease (AD) is the most prevalent cause of dementia, with numbers projected to attain 115.4 million worldwide by 2050.1,2 β-amyloid (Aβ) and hyperphosphorylated tau pathology are major pathological hallmarks of AD. 3 Furthermore, cerebrovascular dysfunction plays a critical role in the earliest pathogenesis of AD. As a strong contributor to vascular dysfunction, Cerebral amyloid angiopathy (CAA) is found in the majority of AD cases. The vascular accumulation of Aβ is the causative agent of CAA, with the characteristics of dysfunctional hemodynamics and leaky blood-brain barrier (BBB). CAA ultimately leads to clearance failure and further accumulation of Aβ in the cerebrovasculature and brain parenchyma.4,5 The perivascular deposition of Aβ in the brain leads to endothelial cell dysfunction and death. 6
Progesterone receptor membrane component 1 (PGRMC1) is a protein of the MAPR family which is involved in multiple functions, including cytochrome P450 regulation, 7 autophagy, membrane trafficking, 8 fertility, 9 steroidogenesis, 10 and AD.11,12 In central nervous system, progesterone has been demonstrated to exert neurogenic and neuroprotective effects via PGRMC1. It has further been shown that PGRMC1 binds Aβ oligomers in AD brain tissue and that this interaction mediates synaptotoxic effects of Aβ. 13 However, the role of PGRMC1 in endothelia cell especially in AD has not been investigated yet. It has been demonstrated PGRMC1 promotes luteal vascularization in mice. 14 PGRMC1 might also contribute to VEGF-induced neovascularization in tumor tissue. 15
In the present study, we investigated the expression of PGRMC1 in AD and explored the role of PGRMC1 in human brain microvascular endothelial cells (HBMVECs) by bioinformatic analysis and experimental verification.
Materials and Methods
Single-Nucleus Transcriptome Analysis of the Prefrontal Cortex in Alzheimer’s Disease
The Clinical Information of Samples.
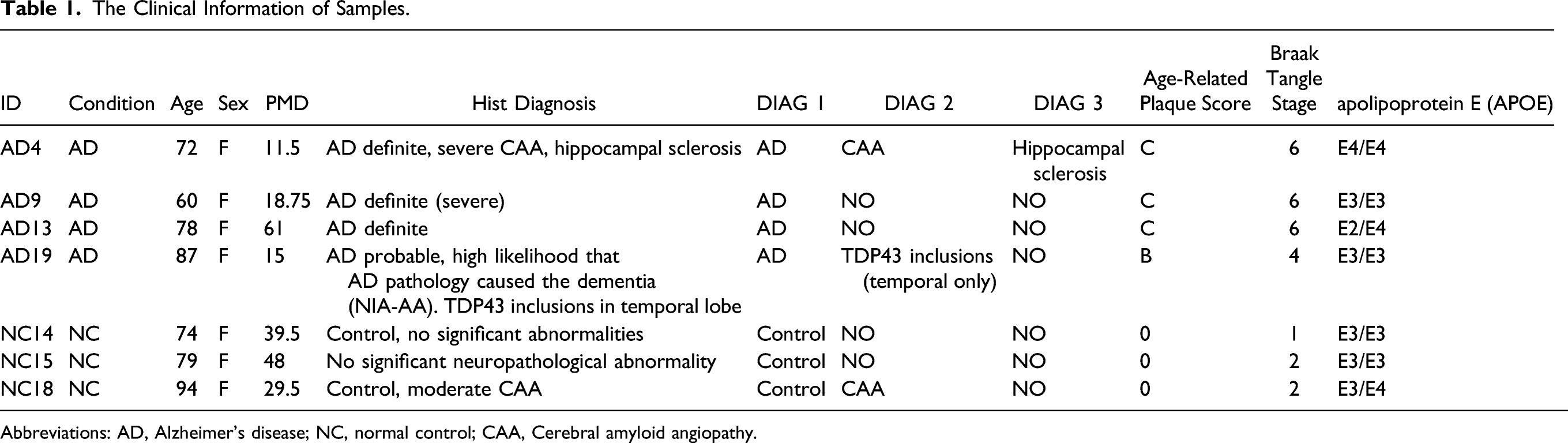
Abbreviations: AD, Alzheimer’s disease; NC, normal control; CAA, Cerebral amyloid angiopathy.
And then data was reanalyzed using R software version 4.0.1 with the package Seurat version 2.460. The “Seurat” package was used to perform the single-cell RNA-seq analysis. The batch effect from studies was removed with regularized negative binomial regression by the “Seurat” package. Dimension reduction of the pre-processed matrix was performed by principal component analysis (PCA). t-SNE visualization of the snRNA-seq data was again performed using the reduced PCA space. Cell cluster identities were manually defined with the cluster-specific marker genes or known marker genes. 16 We constructed and analyzed PPI using BisoGenet in Cytoscape. Genes with the degree centrality (DC) values higher than 60 were selected for subnetwork construction using CytoNCA. 17
RNA-Seq and Data Analysis
Total RNA was isolated using the TRIzol reagent, as described by the manufacturer (15596018, Invitrogen). RNA-seq libraries were constructed by NEBNext® Ultra™ RNA Library Prep Kit for Illumina®. Sequencing-by-synthesis is carried out on the IlluminaHiSeq platform. Raw reads from Illumina sequencing were subjected to adaptor trimming and filtering of low-quality reads by fastp. Differential gene transcripts were analyzed with DESeq2 and enriched for the Gene Ontology/Kyoto Encyclopedia of Genes and Genomes pathway.
Vascular Endothelial Cells Culture
HBMVECs were obtained from Type Culture Collection of the Chinese Academy of Sciences (Shanghai, China). HBMVECs were cultured in DMEM (Dulbecco’s modified Eagle medium) supplemented with the 10% fetal bovine serum (FBS, Gibco, Gran Island, NY, USA) and 100 μg/ml penicillin/streptomycin (Invitrogen; Thermo Fisher Scientific, Inc. USA).
Lentivirus Package and Transfection
PGRMC1 overexpression was accomplished by cloning PGRMC1 cDNA into PCDH vector, and the empty vector was used as a control. The vectors were transfected HEK293T cells for lentivirus packaging. HBMVECs were then infected by HEK293T cell supernatant and were selected in media supplemented with 5 μg/ml of puromycin.
For small interfering RNA oligonucleotides (siRNA) transfection experiments, sequences were transfected into HBMVECs with Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions (PGRMC1-siRNA1 GCCTCTGCATCTTCCTGCTCTACAA, PGRMC1-siRNA2 TCATGGCCATCAACGGCAAGGTGTT PGRMC1-siRNA4 CGGCAAGGTGTTCGATGTGACCAAA). The HBMVECs were harvested after 48 hours. siRNAs were designed and synthesized chemically (GenePharma, RiBio).
CCK-8 Assay
HBMVECs and HBMVECs-PGRMC1 were seeded into 96-well plates at a density of 5 × 103 cells per well. Proliferative viability of HBMVECs was detected using Cell Counting Kit-8 (CCK-8) (Beyotime, C0038) according to the manufacturer’s specifications. Five replicates were performed for each condition. For CCK-8 assay, the cells in 96-well plates were added with CCK-8 solution and incubated for another 1 hour at 37°C (for 0, 24, 48, 72 and 96 hours). 18
Human Brain Microvascular Endothelial Cells Tube Formation Assay
HBMVEC tube formation assay was performed. Briefly, 200 μL of pre-cooled Matrigel (Collaborative BiomedicalProducts) was transferred to each well of a 24-well plate and polymerized for 30 minutes at 37°C. HBMVECs (2 × 104) in 200μL of conditioned medium were added to each well and incubated at 37°C and 5% CO2 for 12 hours. Capillary tube structure was photographed under a × 100 bright-field microscope and quantified by measuring the total length of completed tubes. Four replicates were performed for each condition.
Migration Assay
Migration assay was performed using a 24-well transwell chamber system (Costar 3422, Corning Inc., NY, USA). Briefly, cells were digested with .05% trypsin, and suspended in an FBS-free DMEM culture medium. Cells were then plated into the upper chamber (3 × 103 cells/well). At the same time, 650 μL of DMEM with 10% FBS was added to the lower chamber of the well and the plates were incubated for 5 hours at 37°C with 5% CO2. After incubation, cells on the upper surface of the membranes were removed gently with cotton swabs. Cells that had entered the lower surface of the filter membrane were stained with .1% Hoechst 33342 (H3570, Invitrogen) for 30 minutes at room temperature, and washed three times with PBS. Four randomly selected fields in each well were image captured with the ImageXpress Micro HCS (Molecular Devices), and migrated cells were counted. n = 3 independent experiments.
Western Blotting
Total cell lysates were extracted from the HBMVECs and PGRMC1 overexpressed HBMVECs using the NP-40-lysis buffer. The lysates were transferred to the SDS loading buffer and separated by SDS-PAGE and transferred to PVDF member and blocked with 5% skim milk in TBST. The primary antibodies were incubated overnight at 4°C, and horseradish peroxidase-conjugated secondary antibody at room temperature. The final vision blot was visualized using chemiluminescence reagent (ECL) kit (Beyotime Biotechnology).
Quantitative Real-Time PCR
Total RNA was extracted from HBMVECs using Trizol reagent (Invitrogen) according to the manufacturer’s instructions. The first strand of cDNA was synthesized from 2 μg of total RNA using random primers and reverse transcriptase (Invitrogen). Real-time quantitative polymerase chain reaction (PCR) analyses were performed using the CFX96 Real-Time PCR System (Bio-Rad). GAPDH was used as a loading control for normalization. Each sample was measured in duplicates. The primer sequences are as follows: human GAPDH: sense, 5´- GGACTCATGACCACAGTCCA-3´; antisense, 5´- TCAGCTCAGGGATGACCTTG -3´; human PGRMC1: sense, 5´- TGACCAAAGGCCGCAAATTC-3´; antisense, 5´- CCTCCTTCAGCAGTTTGCC -3´.
Results
Progesterone Receptor Membrane Component-1 is Downregulated in Alzheimer’s Disease
To investigate the potential role of PGRMC1 in AD, we first analyzed the expression level of PGRMC1 in AD and NC, and found that the transcripts of PGRMC1 in AD samples was dramatically decreased compared with the NC samples from both frontal cortex and temporal cortex tissues (Figure 1A). (A) Volcano plots showing DEGs between alzheimer’s disease and normal control in frontal cortex and temporal cortex, respectively. (B) Single-cell RNA sequencing analysis and the expression of PGRMC1 in single-cell dataset GSE157827. (C) Expression of PGRMC1 in different kinds of cells from single-cell dataset GSE157827 ns P > .05 *P < .05, ****P < .0001. Abbreviations: PGRMC1, progesterone receptor membrane component-1.
To establish a baseline profile of cell populations, t-SNE visualization of cell types isolated from the AD and NC samples, with yielded 36 unique cell clusters (AD: 16; NC: 20). Once cell clusters have been identified, these are interpreted according to marker genes from Shun-Fat Lau’s study
16
(Figure 1B). There are six kinds of cells: astrocytes (AQP4, ADGRV1, GPC5, and RYR3), endothelial cells (CLDN5, ABCB1 and EBF1), excitatory neurons (CAMK2A, CBLN2 and LDB2), inhibitory neurons (GAD1, LHFPL3 and PCDH15), microglia (C3, LRMDA and DOCK8), and oligodendrocytes (MBP, PLP1 and ST18). Violin plots showed the expression of representative marker genes across different clusters in both groups (Supplementary Figures 1–12). PGRMC1 exhibited a decrease in AD endothelial cells, microglia, astrocytes, excitatory neurons, inhibitory neurons, and oligodendrocytes (Figure 1C). In the latter four kinds of cells, comparisons were statistically significant. To assess the function of PGRMC1 in AD endothelial cells, we divided ECs in AD to two groups according to PGRMC1 expression. (High PGRMC1-expression group: > 0; Low PGRMC1-expression group: = 0) The differentially expressed gene (DEG) PPI network was comprised 1291 nodes and 28036 edges, and the subnetwork was acquired through selecting the DC which was higher than 60, including 289 nodes and 11315 edges (Figure 2A). Based on the subnetwork, the enriched gene function was highly correlated with regulation of cell proliferation, angiogenesis and etc. (Figure 2B). (A) PPI construction network and topological analysis of DEGs in between low- and high-PGRMC1 expression endothelial cells. (B) Functional enrichment of DEGs between low- and high-PGRMC1 expression endothelial cells. (C) Western blot results showing PGRMC1 was overexpressed in HBMVECs. (D) Real-time PCR results showing the efficiency of PGRMC1 siRNA transfection in HBMVECs. **** P < .0001. (E) Volcano plot showing the fold change and significance of gene expression values between PGRMC1-overexpressing and control HBMVECs. (F) Functional enrichment of DEGs between PGRMC1-overexpressing and control HBMVECs. Abbreviations: PGRMC1, progesterone receptor membrane component-1; HBMVECs, human brain microvascular endothelial cells.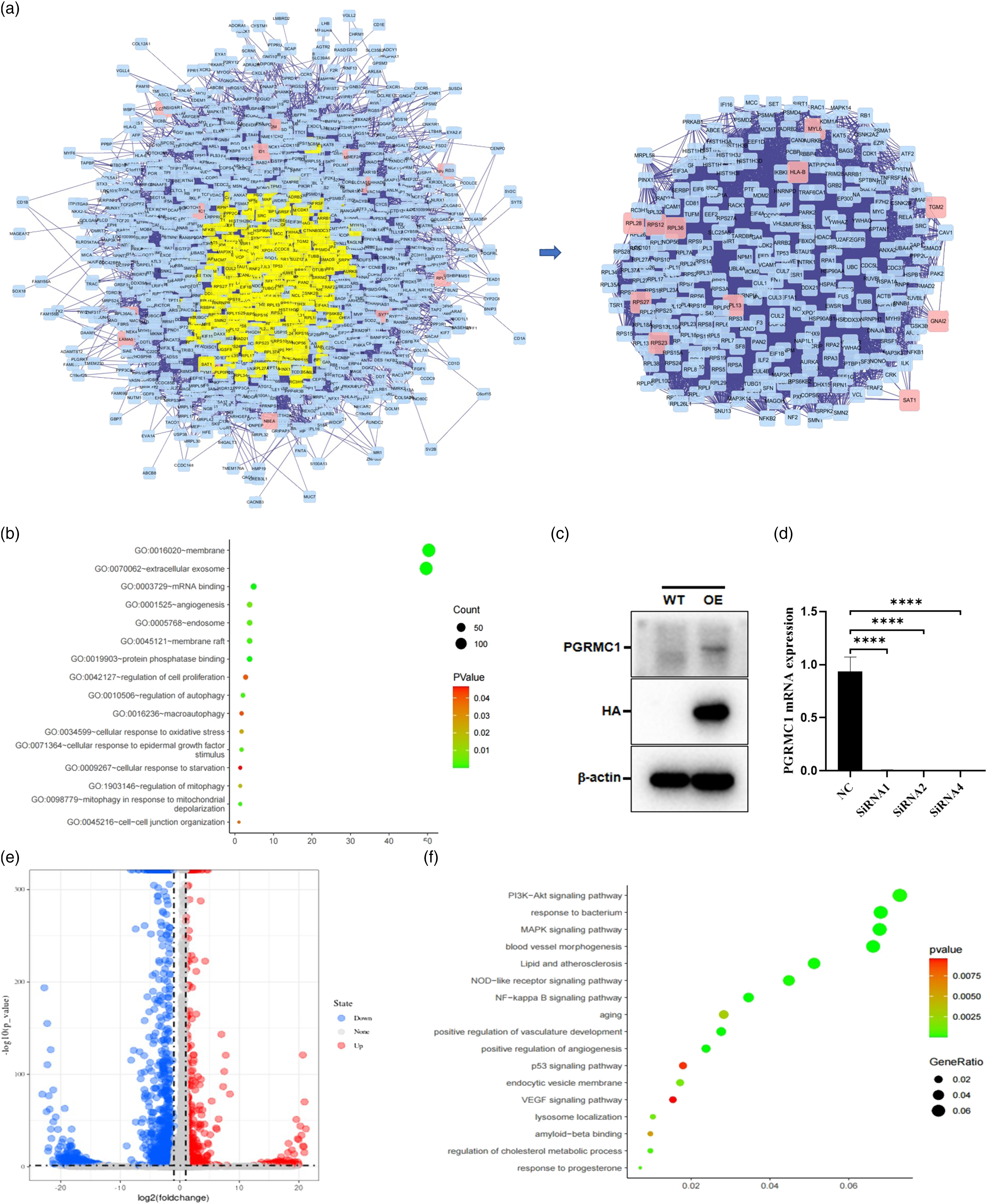
To understand the roles of PGRMC1 in the development of AD, we attempted to overexpress and knockdown PGRMC1 in HBMVECs. First, HBMVECs which stably expresses PGRMC1 was constructed by lentivirus infection and puromycin selection. The protein expression of PGRMC1 was verified by Western blot. As shown in Figure 2C, PGRMC1 was overexpressed compared with control vector. siRNA sequences knocked down PGRMC1 mRNA levels as determined through qPCR in Figure 2D. To validate the above results, we performed the high-throughput sequencing for above PGRMC1 stable cell lines and control. Differential gene expression analysis (P < .05, 2-fold threshold) revealed significant change in 578 (up) and 1559 (down) genes (Figure 2E). Functional enrichment analysis revealed that DEGs were highly correlated with PI3K-Akt signaling pathway, Lipid and atherosclerosis, aging, positive regulation of angiogenesis, VEGF signaling pathway, lysosome localization, amyloid-beta binding, regulation of cholesterol metabolic process, response to progesterone (Figure 2F).
Progesterone Receptor Membrane Component-1 Promotes Human Brain Microvascular Endothelial Cells Proliferation, Migration and Tube Formation
Endothelial proliferation, migration, and tube formation in response to VEGF stimulations are key processes during angiogenesis. To explore the effects of PGRMC1 on HBMVECs, we generated HBMVECs stably expressing PGRMC1. In this study, we demonstrated that PGRMC1 overexpression prominently promoted migration, 48 hours later (P < .05) (Figure 3A, B and E). Correspondingly, PGRMC1 knockdown inhibited cell migration (P < .01) (Figure 3C, D and F). We also found that the overexpression of PGRMC1 significantly promoted proliferation in vitro (Figure 3G). Moreover, overexpressing PGRMC1 in HBMVECs resulted in a significant increase in vessel structures (Figure 4A and B). While, PGRMC1 knockdown inhibited VEGF-induced angiogenesis 12 hours later (Figure 4C and D). The total tube length in the control cells was significantly shorter than those in the PGRMC1 overexpressing cells (P < .05). And the total tube length in the PGRMC1 knockdown cells was significantly shorter than those in NC cells (Figure 4E) (P < .05). Similarly, PGRMC1 overexpressing cells have higher number of branch points than the control group (P < .05) and PGRMC1 knockdown cells (P < .0001), significantly (Figure 4F). Altogether, the above results demonstrated PGRMC1 promotes HBMVECs proliferation, migration and tube formation. Progesterone receptor membrane component-1 promotes cell migration and cell proliferation in vitro. (A-D) Representative images of cells that migrated through the membranes after 48 hours. (magnification ×40). (E-F) The numbers of cells that migrated through the membranes after 48 hours. Data were presented as mean ± SD. The results were reproducible in three independent experiments. (G) The viability of human brain microvascular endothelial cells were tested by CCK-8 assay. *P < .05, **P < .01, ****P < .0001. Progesterone receptor membrane component-1 promotes angiogenesis in vitro. (A-D) Representative images of angiogenic response after 12 hours. (magnification ×100). (E) The number of branch points of angiogenic response after 12 hours. (F) Total tube lengths of angiogenic response after 12 hours. Data were presented as mean ± SD. The results were reproducible in four independent experiments. *P < .05, **P < .01, ***P < .001, ****P < .0001.

Discussion
As the most common cause of dementia, AD severely affects the life quality of elderly patients. Currently, no effective medications are available for AD, because of the limited understanding of its pathophysiological mechanism. 19 Recent researchs demonstrated that frontal periventricular and posterior white matter hyperintensities are associated with CAA in familial AD. 20 Vascular amyloid deposits could be early events in AD and lead to decreased Aβ clearance, and, finally, lead to neurodegeneration. 21 However, new biomarkers and therapeutic targets for CAA in AD are urgently needed.
Previous studies have reported that the PGRMC1 played an important role in the nervous system, especially in the pathological mechanism of AD. Its possible molecular pathways involved in AD include binding to Aβ oligomers, regulation of Ca ion homeostasis, glial cell BNDF secretion, nerve cell apoptosis, nerve cell glucose metabolism, Ras signaling pathway, cholesterol metabolism, etc.11,12,22 In the present study, for the first time, the expression level of PGRMC1 in single-cell RNA-sequencing data of AD were explored. Based on the analysis of transcriptome data of Shun-Fat Lau’s study, 16 PGRMC1 was found to be low expressed in the frontal cortex and temporal cortex tissues of Alzheimer’s disease. We next detected the expression of PGRMC1 in different cell types. The results showed a downward trend in all kinds of cells in AD, including endothelial cells, microglia, astrocytes, excitatory neurons, inhibitory neurons, and oligodendrocytes. Although the decrease in endothelial cells did not reach statistical significance, we found that the DEGs associated with PGRMC1 were highly enriched in regulation of cell proliferation and angiogenesis. Similar findings have been demonstrated in our HBMVECs overexpressed PGRMC1. PGRMC1 overexpression enhanced the proliferation of HBMVECs. Prior studies that have noted the importance of PGRMC1 in cell proliferation and metastasis, such as breast cancer, oral cancer.23,24 What is surprising is that, we demonstrated that PGRMC1 promoted HBMVECs tube formation. To date, few studies have reported the role of PGRMC1 in angiogenesis. Nevertheless, the detailed mechanism remains to be determined yet.
Conclusion
Our findings identified PGRMC1 as a critical gene for AD and revealed its involvement in proliferation, migration, and angiogenesis of HBMVECs. Further study on pathogenesis and hormone might provide new insight into the potential treatment of AD.
Supplemental Material
Supplemental Material - Progesterone Receptor Membrane Component-1 May Promote Survival of Human Brain Microvascular Endothelial Cells in Alzheimer’s Disease
Supplementary Material for Progesterone Receptor Membrane Component-1 May Promote Survival of Human Brain Microvascular Endothelial Cells in Alzheimer’s Disease by Xin Xu, Xiangyan Ruan, Rui Ju, Zhikun Wang, Yu Yang, Jiaojiao Cheng, Muqing Gu, and Alfred O. Mueck in American Journal of Alzheimer's Disease & Other Dementias®.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the first Beijing maternal and child health specialist demonstration units ‘Menopausal Health Specialist’ [[2017]35] and Beijing Municipal Administration of Hospitals’ Ascent Plan [DFL20181401].
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.