
Abstract
Background:
Antibody resistance, both de novo and acquired, is usually related to high risk of recurrence and lower survival rate in gynecologic cancers. Prevention or reversal of the resistance often yields beneficial clinical results. It was reported that anti-human epidermal growth factor receptor 3 monoclonal antibody was effective against trastuzumab-resistant breast cancer cells. Here in our laboratory, an acquired trastuzumab-resistant ovarian cancer cell line, SKOV3-T, was established previously. Further, human epidermal growth factor receptor 3 was observed to be upregulated in this cell line by microarray detection, suggesting that the antagonist against human epidermal growth factor receptor 3 might be effective to inhibit the resistant cells.
Methods:
We developed an anti-human epidermal growth factor receptor 3 monoclonal antibody, LMAb3, and its affinity to bind human epidermal growth factor receptor 3 was calculated by the Biacore method. Preliminarily, LMAb3’s antitumor activity was evaluated in vitro using cell growth/proliferation and clone formation assays in the breast cancer cell line MCF-7. Furthermore, LMAb3 was also evaluated for its inhibitory effect on the carcinogenicity of the SKOV3-T cells, which were induced to overexpress human epidermal growth factor receptor 3, both in vitro and in vivo. The possible underlying signal transduction mechanisms were also identified by Western blot in the MCF-7 and SKOV3-T cells.
Results:
LMAb3 was able to inhibit the cell growth/proliferation, clone, and tumor formation both in vitro (in the MCF-7 and SKOV3-T cells) and in vivo. The underlying mechanism of LMAb3 possibly involves inactivation of the HER family proteins (human epidermal growth factor receptor 1, human epidermal growth factor receptor 2, and especially human epidermal growth factor receptor 3) as well as the downstream mitogen-activated protein kinase and protein kinase B pathways.
Conclusion:
Our work suggests that satisfactory curative effects might be achieved with LMAb3 to treat the trastuzumab-resistant, human epidermal growth factor receptor 3-positive cases of gynecologic cancers.
Introduction
The ErbB (erb-b2 receptor tyrosine kinase) family includes the epidermal growth factor receptor (EGFR), ErbB2 (human epidermal growth factor receptor 2 [HER2]/neu or CD340), ErbB3 (human epidermal growth factor receptor 3 [HER3]), and ErbB4 (human epidermal growth factor receptor 4 [HER4]). They are often abnormally activated in various human cancers. 1 In the recent decades, HER2, the most well-known biomarker for cancer therapy, has been found to correlate with the worst prognosis in several solid tumors, including ovarian, breast, colon, and prostate. 2 –5 Thus, many specific anti-HER2 monoclonal antibodies (mAbs) were approved for clinical cancer therapy by the Food and Drug Administration. For instance, trastuzumab (Herceptin, Genentech, San Francisco, CA, USA), one of the most successful anticancer antibodies, 6,7 offered promising therapeutic advantages not only in breast cancer but also in other tumor types such as gastric cancer. Additionally, in 2012, pertuzumab (Omnitarg, Genentech, San Francisco, CA, USA) was approved for the treatment of metastatic breast cancer with high or low expression levels of HER2. It is a HER heterodimerization inhibitor that binds the domain II of extracellular HER2. Compared to trastuzumab, pertuzumab influences a broader array of downstream signaling pathways through the inhibition of lateral signal transduction. 8 –12 There has also been a lot of excitement about the development of antibody–drug conjugates (ADCs) with improved therapeutic effects, for example, trastuzumab emtansine (Kydcyla/T-DM1, Genentech, San Francisco, CA, USA/Roche, Basel, Switzerland) against metastatic HER2-positive breast cancer. 13,14
Although ADCs against cancers have made significant clinical achievements, in many cases, patients do not respond to the antibody at the beginning. Besides, many patients who receive antibody treatment relapse due to subsequent antibody resistance. 15,16 Examples include resistance against the anti-CD44v6 mAb bivatuzumab 17,18 or anti-EGFR mAbs cetuximab and panitumumab. 19 –22 Owing to this resistance, it was recommended that anti-EGFR mAbs should be given only to patients with KRAS wild-type metastatic colorectal cancer. 23,24 However, the overall response rate is still not high, rangingfrom 17% to 60%. 25 –34
Intrinsic or developed resistance against HER (especially EGFR or HER2)-targeting agents is a clinical problem. Trastuzumab resistance is usually observed in clinical trials. Many reports have focused on optimizing the therapeutic strategies, 35,36 including combination therapy, for example, mAb plus chemotherapy or other mAbs. In a phase III study on women with HER2-positive breast cancer, treatment with trastuzumab and lapatinib (which inhibits both HER2 and EGFR) was successful in extending the progression-free survival by 4 months. 37 Additionally, in a randomized clinical trial, patients with breast cancer who progressed even after previous trastuzumab therapy were treated with trastuzumab plus capecitabine. This yielded significant benefits compared to treatment with capecitabine alone. 38 In all, multiantibody treatments against targets such as the HER family, vascular endothelial growth factor, and IGF-1R have been identified as beneficial to cancer therapy.
The human HER3 is a 185-kDa transmembrane glycoprotein, which is encoded by the ERBB3 gene residing in the chromosome 12q13. It is the only member of the HER family that lacks an intrinsic tyrosine kinase activity. Therefore, its role in cancer has been underestimated for a long time. Further, EGFR and HER2 are the preferred dimerization partners of HER3. 39,40 Heterodimers HER1/HER3 and HER2/HER3 are the most active signaling complexes in this family. 41 Moreover, the HER3/protein kinase B (AKT) pathway in cancer has been studied in several tumor types, including breast, lung, gastric, head and neck, ovarian, and melanoma. In view of its overexpression in tumors and compensatory role in the HER signaling, HER3 is an interesting target that can be utilized to inhibit the HER family signaling. In fact, therapeutic targeting of HER3 has been shown to be an effective way to overcome drug resistance and significantly enhance the antitumor activity of other therapies. The HER3 may be considered a valuable biomarker to predict the efficacy of EGFR- and HER2-targeted therapy of non–small-cell lung cancer and HER2-positive breast cancer, respectively. High levels of HER3 were observed in breast cancer cells and xenograft tumors after treatment with HER2-targeting tyrosine kinase inhibitors (TKIs), and complete inhibition of HER3 was required for optimal anti-HER2 targeting. 36,42 –44 Furthermore, HER3 is associated with endocrine resistance to HER inhibitors in breast cancer, and the treatment of HER2-driven breast cancer cell lines and xenograft tumors with HER-targeting TKIs leads to a rapid compensatory increase in HER3. 35 It was also reported that HER3 confers resistance in HER2-positive breast cancer cells through the phosphatidyl inositol 3-kinase (PI3K)/AKT pathway. 45 Thus, inhibition of the HER3 signaling is known to overcome therapeutic resistance in several cases.
Several drugs that block the signaling of HER3, including fully human anti-HER3 antibodies, bispecific antibodies, and TKIs, have already entered clinical trials. Particularly, HER3 mAbs can inhibit the ligand-induced phosphorylation of HER2, HER3, and downstream effectors including the extracellular signal-regulated kinase (ERK) 1, ERK2, and AKT.
In our previous work, an acquired trastuzumab-resistant cell line model of human ovarian cancer, SKOV3-T, was established. Moreover, upregulation of HER3 was detected in this cell line by microarray analysis, and its pivotal role in cell proliferation was preliminarily identified. 46 In this study, we developed a novel human anti-HER3 antibody, LMAb3, and investigated its inhibitory biological function in the SKOV3-T cells, both in vitro and in vivo.
Methods
Reagents
Trastuzumab (Herceptin) was obtained from F. Hoffmann-La Roche Ltd (Basel, Switzerland). The antibodies for Western blot against EGFR, phospho (p)-EGFR (Tyr1068), HER2, p-HER2 (Tyr1248), HER3, p-HER3 (Tyr1289), AKT, p-AKT (Ser473), glyceraldehyde-3-phosphate dehydrogenase, and the corresponding secondary antibodies were purchased from Cell Signaling Technology (Boston, MA, USA). Phycoerythrin (PE)-conjugated anti-EGFR and anti-HER3 antibodies and fluorescein isothiocyanate-conjugated goat antihuman immunoglobulin (Ig) G (IgG) were from eBioscience (San Diego, CA, USA). The PE-conjugated anti-HER2 antibody was from BD, and HER3 was from Sino Biological Inc (Beijing, China). LMAb3, a novel anti-HER3 mAb (IgG1 κ) was prepared in our laboratory. The reagents for electrophoresis and hybridization nitrocellulose filter membranes were obtained from Bio-Rad (Hercules, CA, USA). The BCA protein assay and enhanced chemiluminescent (ECL) reagents were from Thermo Scientific (Waltham, MA, USA). The cell culture medium Dulbecco’s modified Eagle medium (DMEM) and fetal bovine serum (FBS) were purchased from HyClone (Logan, Utah, USA). Human NRG1-β1/HRG1-β1 were purchased from R&D (Minneapolis, MN, USA), CCK-8 were purchased from Dojindo (Kumamoto, Japan), and agarose (cell culture grade) were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cell Culture
The human breast cancer cell line MCF-7 was purchased from (American Type Culture Collection, ATCC No. HTB-22). The cells were cultivated in DMEM supplemented with 100 units/mL penicillin and streptomycin and 10% FBS.
The acquired trastuzumab-resistant ovarian cancer cell line, SKOV3-T, was developed by continuous culturing of SKOV3 cells in the presence of 20 μg/mL trastuzumab. The surviving cells were pooled together and tested for dose response to trastuzumab as described earlier. 46 The SKOV3-T cells were then maintained in the presence of 10 μg/mL trastuzumab. All the cells were incubated in a humidified incubator (Thermo Scientific, Waltham, MA, USA) at 37°C with 5% CO2.
Proliferation Assay
The cells were loaded in a 96-well plate at a density of 2 × 103 cells/well with or without diluted antibody and supplemented with 10 ng/mL of heregulin (HRG) for 72 hours. Cells incubated with human IgG were used as negative control. Next, 10 μL of the CCK-8 solution was added to 100 μL of medium in each well for 1 to 4 hours at 37°C. The absorbance was then measured at 450 nm using a Tecan Spectrophotometer (Tecan SPECTRAFluor, Tecan, Männedorf, Switzerland).
Biacore Method
The antigen was the standard HER3 protein, and the solvent (sample buffer) was set as the negative control. The background was deducted according to the V-baseline to obtain the kinetic curves and obtained the Kon and Koff constants.
Flow Cytometry
The cells were collected, incubated with specific primary antibodies against the membrane markers, and then stained with PE-conjugated secondary antibodies. For each sample, approximately 15 000 cells were analyzed using a BD-FACStar instrument. Data analysis (the percentage and intensity of stained cells) was performed on a FACS Calibur flow cytometer using the BD CellQuest program.
Enzyme-Linked Immunosorbent Assay
Enzyme-linked immunosorbent assay (ELISA) plates were coated with 2μg/mL HER3 at 4°C overnight. Following blocking with 1.5% BSA in phosphate-buffered saline (PBS) at 37°C for 1 hour, 100 μL of diluted LMAb3 was added. This was then incubated at 37°C for 1 hour, followed by the addition and treatment with 100 μL of horseradish peroxidase (HRP)-conjugated antihuman polyclonal antibody for 45 minutes at room temperature. The peroxidase reaction was developed with tetramethylbenzidine substrate (eBioscience [San Diego, CA, USA]), and the absorbance was measured at 450 nm with an ELISA reader.
Agar Clone Formation Assay
The hydrogel was prepared by dissolving the agarose (0.6% and 1.2% w/t) in aqueous solvent at 90°C, and gelation was allowed to occur by cooling to room temperature. The cells were harvested using 0.25% trypsin, counted, and seeded in the agarose scaffolds. Briefly, the lower agarose layer (1.2% agarose in 2× DMEM with 10% FBS and 10 ng/mL of HRG) was allowed to gel at 4°C for 20 minutes in a 6-well culture plate. The cells were resuspended at a density of 1000 cells/mL in 0.6% agarose supplemented with/without LMAb3 (upper agarose layer). The plate was incubated at 37°C and 5% CO2. As a control, the cell aggregates were cultured in the same experimental conditions. The culture media were changed every 3 days. The assays were performed at ian interval of 14 days.
Western Blot
The cells were washed with cold PBS before they were lysed in the lysis buffer (20 mmol/L Tris at pH 7.0, 1% Triton-X 100, 0.5% NP-40, 250 mmol/L NaCl, 3 mmol/L EDTA, 3 mmol/L ethylene glycol tetraacetic acid, 2 mmol/L
In antibody treating assay, the cells were cultured in a 6-well plate and serum-starved overnight. Next, the cells were incubated with/without diluted LMAb3 or trastuzumab for 4 hours. The cells were then stimulated with 10 ng/mL of HRG for 20 minutes and subsequently collected for western blot analysis.
In Vivo Immunotherapy Assay
Twenty-four 5-week-old female BALB/c athymic and nu/nu (nude) mice were inoculated on the fat pads with 1 × 106/0.1 mL SKOV3-T cells on day 0. On day 7, the mice bearing palpable tumors were randomly divided into 3 groups with 8 mice per group (n = 8) and treated intravenously with 5 mg/kg LMAb3 (about 100 μg/mouse), trastuzumab, or natural saline (negative control) once a week for 4 weeks. The body weights, survival, and tumor volumes were checked twice a week, starting from day 7. The tumor volume was evaluated according to the following equation: tumor volume (mm3) =1/2 × (length) × (width). 2 Pairwise differences between the groups were compared.
We followed the guidelines for the welfare and use of animals in cancer research. Use and treatment of the mice were in strict agreement with the international guidelines for the care and use of laboratory animals. Further, the protocols were approved by the Animal Ethics Committee of the Institute of Basic Medical Sciences.
Statistical Analyses
The data are expressed as the mean ± standard deviation. Pairwise differences between the groups were compared. Significant differences were analyzed by analysis of variance, and P values <.05 were considered statistically significant.
Results
LMAb3 Inhibits the Growth of HER3-Positive MCF-7 Cells
Our ELISA results showed that LMAb3 binds HER3 in a dose-dependent manner (Figure 1A). Furthermore, using the biacore method, the binding affinity of LMAb3 to HER3 was calculated to be 2.46 × 10−10 (Figure 1B).

LMAb3 binds specifically to human epidermal growth factor receptor 3 (HER3) with high affinity. A, Enzyme-linked immunosorbent assay (ELISA) assays show that LMAb3 binds to soluble HER3 in a dose-dependent manner. B, Determination of the affinity constant of LMAb3 binding to soluble HER3, by the Biacore method. A high affinity of 2.46 × 10−10 (Kon = 5.66×105 M−1 s−1 and Koff = 1.39×10−4 s−1) was observed.
Next, we also identified the inhibitory activity of LMAb3 in the breast cancer cell line MCF-7. First, the MCF-7 cells were identified to be membrane HER3, as well as HER1 and HER2 positive using standard PE-conjugated anti-HER antibodies (Figure 2A). As shown in Figure 2B, the cell proliferation was repressed specifically by LMAb3 in a dose-dependent manner. For cells treated with 100 μg/mL LMAb3, the survival rate was ∼80% in contrast to the control IgG-treated cells. However, the survival rate decreased to ∼50% for cells treated with 103 μg/mL LMAb3. Furthermore, the agar clone-forming ability of the MCF-7 cells was also inhibited by LMAb3. At 100 μg/mL antibody concentration, the average clone number was less than 30, in contrast to ∼60 (P < .05) for the nontreated samples (Figure 2C).
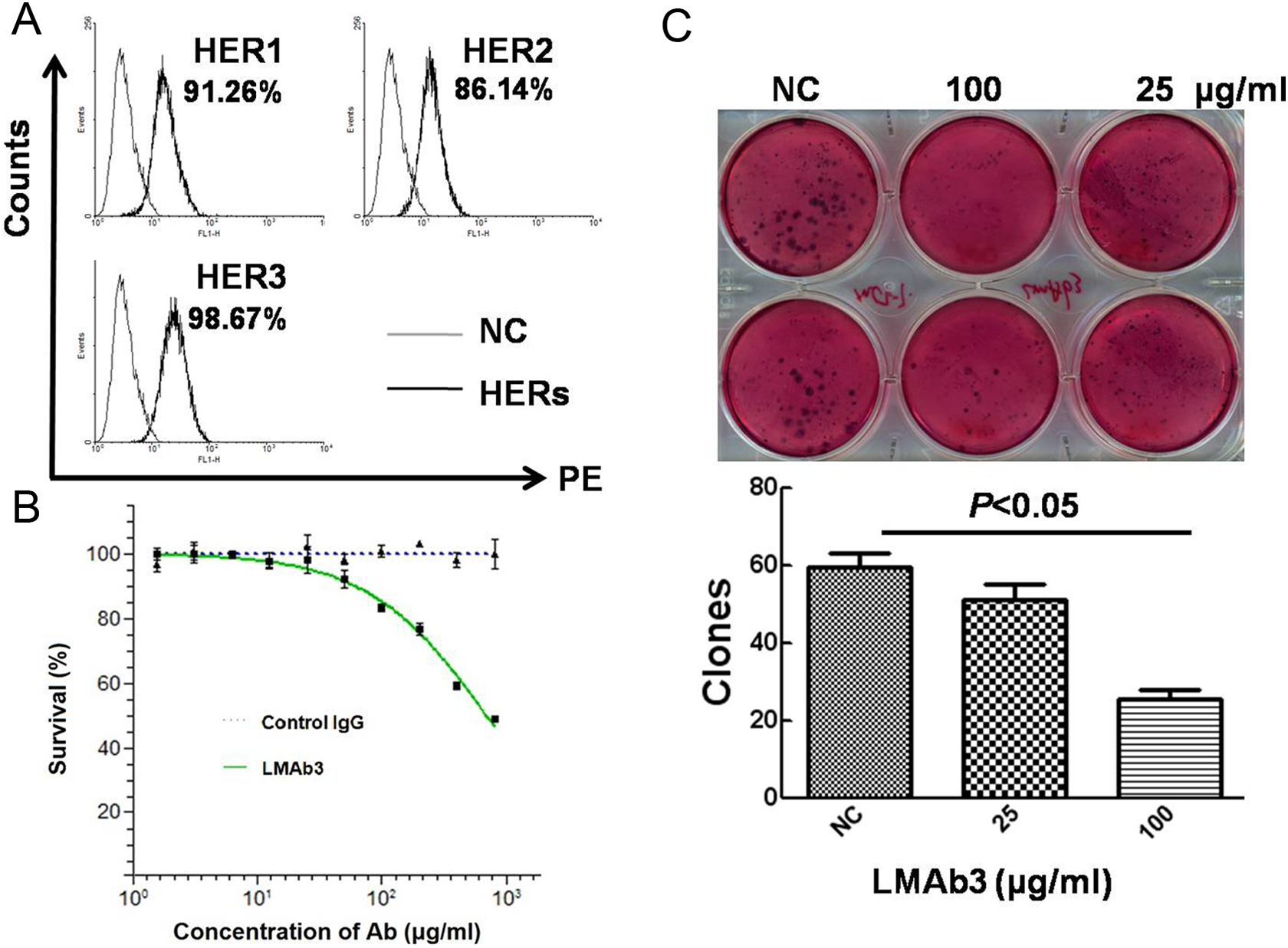
LMAb3 inhibits the growth of MCF-7 cells. A, The expression of membrane HER1, HER2, and HER3 in MCF-7 cells. B, LMAb3 inhibits the proliferation of MCF-7 cells in a dose-dependent manner. C, LMAb3 inhibits the clone formation of MCF-7 cells, indicating its anticancer activity in vitro; 100 μg/mL LMAb3 could inhibit ∼60% clone formation (P < .05), and the monoclones were smaller than those in the negative control samples.
The possible mechanism by which LMAb3 inhibits the cell growth was also investigated by Western blot. In the antibody-treated MCF-7 cells, the HRG (a HER3-specific cell factor)-driven HER family activation (p-HER1, p-HER2, and p-HER3) was inhibited in proportion with the increasing LMAb3 concentrations (Figure 3A). Moreover, the phosphorylation of AKT, the key signaling molecule in the HER family-related pathway, was also inhibited, indicating that the inhibition of the AKT cascade might be one of the antitumor mechanisms adopted by LMAb3 (Figure 3B).
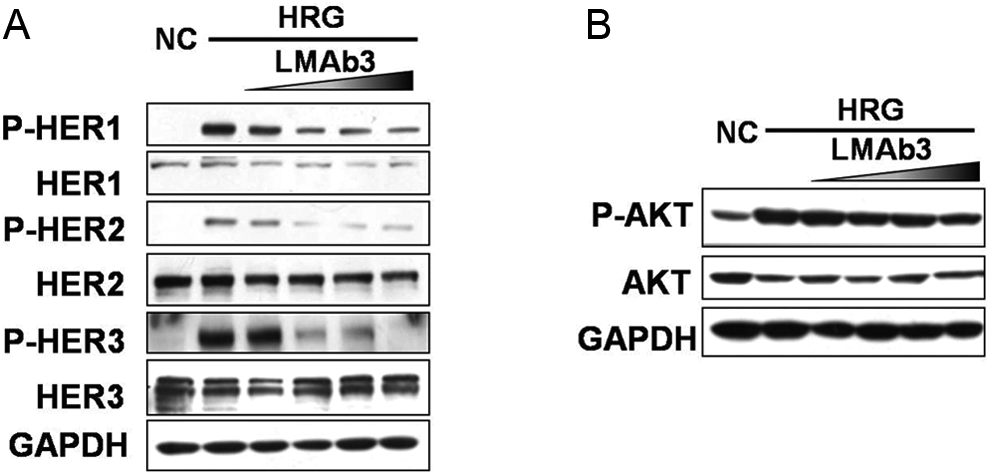
LMAb3 inhibits activation of the human epidermal growth factor receptor (HER) family (A) and hinders the downstream protein kinase B (AKT) signal transduction (B) in human epidermal growth factor receptor 3 (HER3)-positive MCF-7 cells. The concentrations of LMAb3 used were 0.05, 0.5, 5, and 50 μg/mL. According to the Western blot results, the HER family members human epidermal growth factor receptor 1 (HER1), human epidermal growth factor receptor 2 (HER2), and human epidermal growth factor receptor 3 (HER3) and the related key molecule AKT are presumably activated by the HER3 ligand heregulin (HRG), and LMAb3 blocks the HRG-driven stimulation in a dose-dependent manner, thus retarding the cell growth and clone formation.
LMAb3 Inhibits the Growth of SKOV3-T Cells
The human ovarian cancer SKOV3 cells, which overexpress HER2, were cultured continuously for 8 months in the presence of 20 μg/mL trastuzumab, resulting in the acquired trastuzumab-resistant SKOV3-T cells. Our flow cytometry data revealed that the SKOV3-T cells had decreased membrane HER2 and higher HER3 levels compared with the parental SKOV3 cells (Figure 4A). We also analyzed the expression of the HER proteins by Western blot assays. As shown in Figure 4B, the SKOV3-T cells displayed dramatically decreased HER2 and increased HER3 levels in the whole cell lysate. The downregulation of HER2 might be one of the possible reasons behind the trastuzumab resistance acquired by SKOV3-T cells.

The acquired trastuzumab-resistant cell line SKOV3-T overexpresses human epidermal growth factor receptor 3 (HER3). A, Flow cytometry analysis of human epidermal growth factor receptor 1 (HER1), human epidermal growth factor receptor 2 (HER2), and HER3 expressions in the SKOV3 and SKOV3-T cells. The cells were cultured for 8 months in the presence of 20 μg/mL trastuzumab to obtain the SKOV3-T cells. S-T: SKOV3-T cells. B, Western blot analysis of the human epidermal growth factor receptor (HER) proteins in the SKOV3 and SKOV3-T cells. The cells were trypsinized, counted, and 5 × 105 cells/sample were incubated with the standard phycoerythrin (PE)-conjugated anti-HER antibodies. In contrast to the parent SKOV3 cells, SKOV3-T have significantly reduced HER2 and enhanced HER3 expression levels, in which lower HER2 expression might directly induce the acquired resistance to trastuzumab (Herceptin).
Since the trastuzumab-resistant SKOV3-T cells overexpress HER3, we investigated whether LMAb3 can inhibit their proliferation. According to our results, LMAb3 could inhibit the clone formation ability of the SKOV3-T cells. At 5 μg/mL LMAb3 concentration, the mean clone number was ∼30, which further decreased to ∼20 at 50 μg/mL concentration in contrast to >40 clones in the nontreated group (P < .05 and P < .01, respectively; Figure 5A). In vivo experiments, using a xenograft model with SKOV3-T injection, showed that LMAb3 treatment could inhibit the tumor growth. In contrast to the control group with a mean tumor volume of >650 mm3 at the end of the assay, the mice administrated with LMAb3 had a mean tumor volume of ∼500 mm3 (P < .05; Figure 5B).

LMAb3 inhibits the proliferation/growth of SKOV3-T cells. A, Agar cloning assay of SKOV3-T cells treated with LMAb3 or left untreated; 5 μg/mL or 50 μg/mL LMAb3 prohibited the clone formation to two-third or less than one-half of the negative control sample, respectively. B, In vivo carcinogenic identification in xenograft nude mice. The mice were injected with SKOV3-T cells in the fat pads on day 0 and administrated with 5 mg/kg LMAb3, Herceptin, or control immunoglobulin G (IgG) isotype on days 7, 14, 21, and 28. The LMAb3-treated mice exhibited slower tumor growth rate. NC indicates negative control cells (incubated with 50 μg/mL of human IgG).
LMAb3 Inhibits the HER3-Related Signal Cascades in SKOV3-T Cells
The cell signaling assay results showed that LMAb3 could block the HRG-induced activation of the HER family members. This indicated the existence of membrane HER3 and its potential signaling activity (Figure 6A). However, the levels of the HRG-driven activated HERs (p-HER1, p-HER2, and p-HER3) as well as the key signal molecule p-AKT decreased with increasing doses of LMAb3. This suggested that the inhibition of HER3 and its downstream signal cascades, such as the PI3K-AKT pathway, might be one of the antitumor mechanisms of LMAb3 (Figure 6B and C).

LMAb3 inhibits activation of the human epidermal growth factor receptor (HER) family and blocks the downstream signal transduction in SKOV3-T cells. A, Western blot analysis shows that the human epidermal growth factor receptor 3 (HER3)-specific ligand, heregulin (HRG), induces the activation of human epidermal growth factor receptor 1 (HER1), human epidermal growth factor receptor 2 (HER2), and HER3. B, Activation of the HER family members by HRG is inhibited by preincubation with LMAb3. The bands for p-HER1, p-HER2, and p-HER3 are weaker in the antibody-treated versus untreated samples. C, LMAb3 inhibits the downstream activation of protein kinase B (AKT) in a dose-dependent manner. The LMAb3 concentrations used were 0.05, 0.5, 5, and 50 μg/mL. That LMAb3 is able to treat the SKVO3-T cells follows from the observation that the HRG-induced HER family activation and HER-related downstream signal cascade were efficiently blocked. NC indicates negative control SKOV3-T cells.
Discussion
Clinical cases of antibody resistance are on the rise. Many patients either do not respond to antibody treatment at all (de novo resistance) or relapse after receiving the antibody treatment (acquired resistance). Trastuzumab resistance in patients with breast cancer is one such example of acquired resistance. These cases show low recurrence-free survival, cancer-related survival, and/or overall survival and high risk of local and distant recurrence. Many studies have indicated that induction of the HER3 pathway is one of the reasons underlying this kind of resistance. 47,48 For example, a subset of patients with colorectal cancer, who exhibited either de novo or acquired resistance to a cetuximab-based therapy, had HER2 amplification or high levels of circulating HRG (which activates the HER3 signaling). 49 Furthermore, it was shown that dual targeting of EGFR and HER3 can overcome acquired resistance to cetuximab and erlotinib, thereby suggesting the importance of HER3 in the development of resistance to EGFR-targeted therapy. 50 Besides, the HER3 pathway is also known to contribute to trastuzumab resistance. 9,51 –53
The primary ligands that bind HER3 are the members of the neuregulin (NRG) family, including NRG1/HRG. 54 They bind to the extracellular domain of HER3, which permits its heterodimerization with other family members like HER2. Subsequently, the HER2/HER3 complex elicits the most biologically active and potent stimulation of the downstream PI3K/AKT pathway, 1,55,56 which is important in tumor regulation, including survival, translation, nutrient sensing, cell cycle control, and metabolism.
Anti-HER3 drugs, such as antibodies and TKIs, are now becoming available for the treatment of cancers. Several anti-HER3 mAbs exhibit satisfactory antitumor efficacy in vivo. 57,58 They often exert their functions via various mechanisms that include inhibiting the heterodimerization, blocking the HRG ligand-binding to HER3, and/or downregulating the HER3 level in the membrane. Furthermore, some mAbs are able to engage immune effector cells, which lead to antibody-dependent cellular cytotoxicity. Anti-HER3 antibodies, including AMG 888 (in phase II clinical trials) 59 –61 and MM-121 (phase I), possess antitumor activity against trastuzumab-resistant breast cancer cells. 62 –65
Previously, we established a trastuzumab-resistant ovarian cancer cell line, SKOV3-T. Using the biochip technology, we analyzed the differences in messenger RNA expressions between SKOV3 and SKOV3-T. Interestingly, HER3 presented itself as a significantly altered membrane marker in this analysis. 46 In the SKOV3-T cells, downregulation of the membrane HER2 indicated an “epitope escape,” which induced trastuzumab resistance directly, whereas the HER3 pathway helped to maintain the cell growth/proliferation. We thus inferred that an anti-HER3 mAb might be effective in treating the resistant cell line.
In this study, we developed a functional anti-HER3 mAb, LMAb3, using computer-aided design methods, 66,67 based on the 3-dimensional structure of HER3. LMAb3 binds to coated HER3 specifically with high affinity, and LMAb3 inhibited the proliferation in a dose-dependent manner, revealing the potential antitumor activity of LMAb3 in this cancer. The potential mechanism might involve blocking the activation of the HER3/AKT signaling pathway, as well as the HER1 and HER2 phosphorylation, which is required for heterodimerization with HER3. Initially, we attempted to establish Herceptin-resistant cell lines of gynecological cancers using both SKOV3 (ovarian cancer) and BT474 (breast cancer) as the parent cell lines. However, we successfully established only the SKOV3-T cell line where higher level of HER3 was detected than the parent cells. LMAb3 was also able to inhibit the in vitro cell proliferation as well as the in vivo tumor growth, and the underlying mechanism possibly blocks the HRG-stimulated HER3/HER1/HER2 activation and specifically inhibits the downstream AKT phosphorylation. It is worth mentioning here that we also investigated whether LMAb3 can restore the sensitivity of the SKOV3-T cells to trastuzumab. However, we failed to obtain positive results (data not shown). We inferred that the “epitope escape” of HER2 is the major reason or at least one of the major reasons for trastuzumab resistance in this study, while the high HER3 levels are beneficial for maintaining the growth/proliferation of the resistant cancer cells.
Conclusion
We achieved satisfactory results using LMAb3 to treat trastuzumab-resistant ovarian cancer with high levels of HER3. We thus infer that in clinical trials with resistant phenomena similar to that in this study (acquired or de novo), better curative effect might be achieved by combined antibody therapies, such as trastuzumab plus antibodies against other individual biomarkers such as HER3.
Footnotes
Authors’ Note
Xinying Li, Yanting Duan, and Chunxia Qiao contributed equally to this work. LX carried out the agar cloning, Biacore, and immunotherapy assays in vivo. DY cultured the MCF-7 cells, performed fluorescence-activated cell sorter assays, and assisted in the in vivo carcinogenic assays. QC performed the Western blots and purified LMAb3. ZT cultured the SKOV3-T cells. YM screened the monoclones. GJ performed the ELISA assays. FJ performed statistical analyses of the data. SB helped to draft the manuscript. LM designed the experiments. LY prepared the manuscript. All the authors read and approved the final manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article. This work was supported by the National Natural Sciences Foundation of China grant (Nos. 81272528 and 31370938), the Beijing Natural Science Foundation (No. 5152022), and the National High Technology Research and Development Program (863 program, No. 2012AA02A302).