
Abstract
Cancer cells are characterized by accelerated proliferation and an outstanding adaptation of their metabolic pathways to meet energy demands. The folate cycle, also known as folate metabolism or one-carbon metabolism, through enzymatic interconversions, provides metabolites necessary for nucleotide synthesis, methylation, and reduction power, helping to maintain the high rate of proliferation; therefore, the study of this metabolic pathway is of great importance in the study of cancer. Moreover, multiple enzymes involved in this cycle have been implicated in different types of cancer, corroborating the cell's adaptations under this pathology. During the last decade, nonalcoholic fatty liver disease has emerged as the leading etiology related to the rise in the incidence and deaths of hepatocellular carcinoma. Specifically, cholesterol accumulation has been a determinant promoter of tumor formation, with solid evidence that an enriched-cholesterol diet plays a crucial role in accelerating the development of an aggressive subtype of hepatocellular carcinoma compared to other models. In this review, we will discuss the most recent findings to understand the contribution of folate metabolism to cancer cells and tumor microenvironment while creating a link between the dynamics given by cholesterol and methylenetetrahydrofolate dehydrogenase 1-like, a key enzyme of the cycle located in the mitochondrial compartment.
Introduction
Normal and pathological cell proliferation is accompanied by an adjustment to the mechanisms of metabolism to fulfill the needs of the cell. 1 In the cancer cell, redirection of energy metabolism enhances cellular capacities for survival under stress conditions and proliferation, placing the metabolic rewiring as a distinctive feature of the transformed cell. 2
The natural history of liver disease initiates with nonalcoholic fatty liver disease (NAFLD), progressing to nonalcoholic steatohepatitis (NASH), fibrosis, cirrhosis, and eventually cancer. 3 The progression of the disease to hepatocellular carcinoma (HCC) is explained by the chronic low-grade inflammatory response observed in liver cancer patients. 4
HCC is the most frequent form of liver cancer, accounting for approximately 90% of cases. 5 HCC ranks sixth in incidence, yet it represents the third leading cause of death worldwide, leaving clear the need for more mechanistic research to understand this complex disease completely. Among the etiologies for developing HCC are hepatitis B and C virus infection (HBV and HCV, respectively), and alcohol consumption have been highlighted. 6 During the last decade, there have been changes in the burden and etiology of liver diseases, placing NASH, the inflammatory state of the NAFLD, as the leading cause of incidence and death of HCC. 3
Liver lipids overload plays an essential role in the generation of NASH and the progression to HCC. Specifically, cholesterol accumulation has been a determinant promoter in the disease.7–9 Recently, it has been reported in murine models that high-fat diets supplemented with cholesterol (HFHC) cause more significant liver damage compared to mice fed high-fat diets (HFD), with higher serum alanine transaminase (ALT) activity, 10 an enzyme implicated in the amino acids interconversion with a higher content in the liver, making it a specific marker for liver damage. 11 Parallel to these studies, dietary cholesterol conditions the development of a subtype of HCC with increased aggressivity and poor prognosis, revealing a mechanism dependent on the overproduction of reactive oxygen species (ROS). 8 It was previously proved that animals fed with cholesterol-enriched diets (such as the Western diet) exhibited a gene signature associated with aggressivity and poor prognosis. 12
Interestingly, we observed that the most prominent members of this signature were genes related to metabolism control, such as solute carrier family 41 member 3 (Slc41a3), fatty acid-binding protein 5 (Fabp5), immunoglobulin superfamily DCC subclass member 4 (Igdcc4), and methylenetetrahydrofolate dehydrogenase 1-like (Mthfd1l), all these overexpressed in the animal subjected to these cholesterol-enriched diets. These data correlated with the signature observed in patients with a more aggressive HCC phenotype. 12
It was interesting to find the Mthfd1l, a significant gene involved in highly aggressive HCC, and that this was associated with cancer evolved under NASH conditions.
This review focuses on folate metabolism and its implication in cancer hallmarks, highlighting the role of methylenetetrahydrofolate dehydrogenase 1-like (MTHFD1L), a key enzyme of the folate cycle which has been gaining attention in cancer in recent years, and it is positioned as a promising therapeutic target.
Folate
Folate is an umbrella term for a wide variety of related compounds. These can be present in food, then called “food or dietary folates,” or in synthetic forms known as folic acid, such as vitamin B9, frequently found in multivitamin supplements. 13
The intestinal microbiota is responsible for the de novo synthesis of folate in humans. They are absorbed through the intestine and incorporated into the host's tissues. Mammalian cells cannot synthesize folates; therefore, dietary intake is the only way to meet the daily needs of these vital molecules, in addition to those obtained in multivitaminics. 14
Folates are fundamental for life. To mention a couple of iconic cases, the participation of N10-formyl-tetrahydrofolate (carbon 2) and N5-N10-methenyl-tetrahydrofolate (carbon 8) in the biosynthesis of purines and the formylation of methionine at the start of translation.15,16 These 2 exceptional examples leave clear the vital relevance of folates.
Structure and Bioavailability of Folates
The common central structure of folates consists of a pteridine ring, which can be reduced or oxidized, allowing H+ to join the molecule due to its lack of electrons keeping each other together, and vice versa when oxidation happens. At physiological pH, folates are composed of (1) a pteridine ring that can be oxidized or reduced, (2) a para-aminobenzoic acid (PABA) that binds to the one-carbon (1C) units, and (3) a negatively charged glutamate tail of a variable length as can be seen in Figure 1.17,18 The classification of this molecule depends on the number of glutamate residues; it has a monoglutamate (a single glutamate residue) or polyglutamate (2 or more glutamate residues). 13
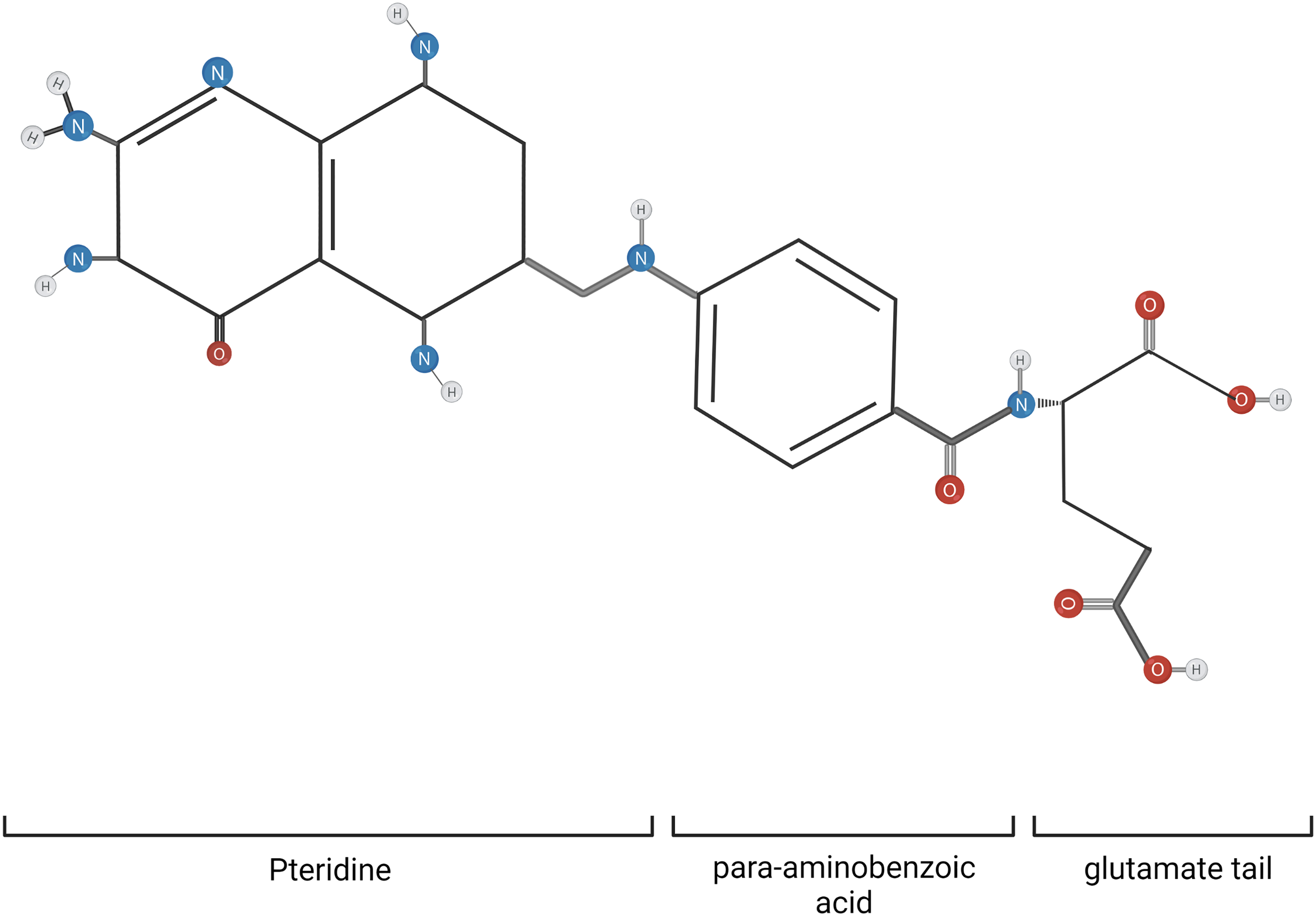
Folate chemical structure. The general structure of folates consists of a pteridine moiety, a para-aminobenzoic acid, and a glutamate tail. They differ in the extent of polyglutamylation and oxidation state. Created with Biorender.com.
The bioavailability of folate is governed by the degree of folate absorption in the intestine. 19 The proximal small intestine is the leading site of folate absorption. Monoglutamate but not polyglutamate forms are transported across the intestine, and dietary folates are mainly polyglutamylated; that is why folate hydrolysis must come first. Intestinal glutamate carboxypeptidase II (GCP II) is responsible for carrying out this activity, and it is ubicated within the brush-border membrane of the jejunum with optimal activity at pH 6.5. 20 On the other hand, supplements such as 5-Methyl-THF salt are monoglutamylated forms and can enter without any transformation. However, dietary folates and folic acids share a common metabolic fate because they are metabolized by L-5-Methyl-THF (5-MTHF). 21
Once in the cell, the addition of glutamate residues (polyglutamylation) is carried out by the action of a folylpolyglutamate synthase (FPGS), which allows folate retention intracellularly. When it is necessary for folate exportation to the external environment, the γ-glutamyl hydrolase (GGH) catalyzes the hydrolysis of the polyglutamylated folate. 22 The hydrolysis of the polyglutamate chain is required since the polyglutamylated forms cannot cross cell membranes; however, upon entering the cell, the polyglutamate chain of this compound is reconstituted. 14 In this way, FPGS and GGH work synergistically to regulate intracellular folate concentration. 22 In addition, polyglutamylated folates are better substrates for intracellular enzymes involved in its metabolism. 23
Folate Transport
Folates are bivalent and highly lipophobic molecules, meaning their simple diffusion through biological membranes is very low, so they must be internalized through mediated processes. 24 As we mentioned before, only monoglutamylated forms are transported. Three primary transporters are recognized: the reduced folate transporter (RFC), the proton-coupled folate transporter (PCFT), and the folate receptor (FOLR). 14
The RFC is an anion transporter member of the solute carrier (SLC) transporters superfamily, specifically the form SLC19A1, and exchanges reduced folate by another anion, such as thiamine phosphate (ThPP). 25 This transporter is ubiquitously expressed in all human tissues, although the highest levels have been detected in the placenta and the liver. In other tissues such as the kidneys, lungs, bone marrow, intestine, brain, and parts of the central nervous system, appreciable levels have been detected 26 ; SLC19A1 is considered the main system of folate transport. 27
On the other hand, the PCFT or SLC46A1 is a transporter optimally operating at low pH. 28 It is preferentially expressed in the acidic microenvironment of the apical brush border membrane of the proximal small intestine and mediates intestinal proton-couple folate absorption.20,28
Finally, high-affinity folate receptors (FOLRs, also called FRs) are folate-binding glycoproteins that mediate folate uptake by endocytosis. 29 There are 4 Folr genes: Folr1-4 (also known as folate receptors α, β, γ, and δ and the classical name folate binding protein).30,31 Once in the endosome, its acidic environment promotes the release of folate from the receptors, which is subsequently transported to the cytoplasm by the PCFT. 32 Folr1 (FR-α) expression is restricted to various epithelial cells; among them, the epithelia of the choroid plexus, the proximal renal tubules, the fallopian tubes, the uterus, and the epididymis, the acinar cells of the mammary, submandibular salivary and bronchial glands, and the trophoblasts of the placenta. 33 Folr2 (FR-β) is expressed during normal myelopoiesis and is present in the placenta, spleen, and thymus, and is also expressed in a subset of macrophages.34–36 Folr3 (FR-γ) has been detected in normal hematopoietic cells in the spleen, bone marrow, and thymus. 37 Folr4 (FR-δ) is constitutively expressed in high amounts by naturally occurring regulatory T lymphocytes (Treg) and is also a biomarker that distinguishes CD4T follicular helper (Tfh) cells from Th1 cells.38,39(p4)
Folate Metabolism
First, in the enterocyte, folates require modifications to be integrated into folate-dependent reactions since they are not in their biologically active form. To achieve this, dihydrofolate reductase (DHFR), using 2 successive reductions of the pteridine ring, first converts monoglutamylated folates and folic acids to dihydrofolate (DHF) and subsequently to the biologically active form, tetrahydrofolate (THF). 40 Later, THF is converted to 5,10-methylene-THF by serine hydroxymethyltransferase (SHMT1) and 5-MTHF by methylenetetrahydrofolate reductase (MTHFR). Finally, 5-MTHF, which comprises about 98% of the plasma folates, 21 exists in the enterocyte through the multidrug resistance-associated protein (MRP) 3, and the contribution of other basolateral transporters is still unclear. 41
Once in the portal blood, 5-MTHF enters the hepatocyte mainly through RFC and FOLR, yet the hepatocyte conserves DHFR activity too. 5-MTHF is a poor substrate of the FPGS, so the first step is its conversion to THF by methionine synthase (MTR). 42 Thus, polyglutamylated-THF accepts and transfers 1C units in a network of biosynthetic and catabolic reactions known as the folate cycle. This cycle comprises the transfer of one-carbon units in different oxidation states between substrates, where such transfers are interconvertible through enzymatic redox reactions. 43 The folate cycle plays a role in integrating various critical nutrients for cell maintenance, referred to as “folate metabolism” or “one-carbon metabolism” due to its participation in other metabolic pathways, such as the methionine cycle and the trans-sulfuration pathway, supporting purine and S-adenosylmethionine (SAM) synthesis, epigenetic regulatory functions, and control of cellular redox status16,18,44,45 (Figure 2).

One-carbon unit metabolism. The compartmentalization of one-carbon metabolism is shown in the figure. The more abundant circulating form of folate is 5-Methyl-THF. Once in the cell, the first reaction is led by MTR to render THF, starting the transfer of 1C unit from one THF position to another by interconvertible redox reactions in the cytosolic and mitochondrial compartments, generating key metabolites for cell growth and epigenetics modification. Activities of enzymes are 10-formyl-THF synthase (1), 5,10-methenyl-THF cyclohydrolase (2), and 5,10-methylene-THF dehydrogenase (3), both in the cytosolic and mitochondrial compartments. Proton coupled folate transporter (PCFT); reduced folate transporter (RFC); folate receptor (FR); tetrahydrofolate (THF); 5-Methyl tetrahydrofolate (5-Methyl-THF); 5,10-Methylene tetrahydrofolate (5,10-Methylene-THF); 5,10-Methenyl tetrahydrofolate (5,10-Methenyl-THF); 10-Formyl tetrahydrofolate (10-Formyl-THF); methylenetetrahydrofolate dehydrogenase, cyclohydrolase, and formyltetrahydrofolate synthetase 1 (MTHFD1); methylenetetrahydrofolate dehydrogenase 1-like (MTHFD1L); methylenetetrahydrofolate dehydrogenase 2/ 2 like (MTHFD2/L); serine hydroxymethyltransferase (SHMT); mitochondrial folate transporter (MFT); methylenetetrahydrofolate reductase (MTHFR); methionine synthase (MTR); S-adenosylmethionine (SAM); Sideroflexin 1 (SFXN1); Solute Carrier Family 25 Member 38 (SLC25A38) nicotinamide adenine dinucleotide (NAD+); reduced nicotinamide adenine dinucleotide (NADH); nicotinamide adenine dinucleotide phosphate (NADP+); reduced nicotinamide adenine dinucleotide phosphate(NADPH); thiamine phosphate (ThPP). Created with BioRender.com.
Cycle Compartmentalization
One-carbon metabolism is highly compartmentalized in eukaryotic cells with THF-dependent enzymes in the mitochondria and cytosol. 46 These cellular compartments are metabolically linked by the transport of serine, glycine, and formate across mitochondrial membranes. Although compartmentalized, the cycle is supported by a mainly unidirectional flow (from mitochondria to cytosol), represented in Figure 2. The mechanism is known due to the tracing of 1C units from serine to formate and methionine, where the units incorporated into the metabolic routes of the cytosol come mainly from the mitochondria 47 ; because of this, the growing scientific attention is focused on the understanding of the functions of the mitochondrial compartment.
The metabolism of serine, glycine, and THF in the mitochondria is initiated by the mitochondrial isoenzyme of serine hydroxymethyltransferase (SHMT2), 48 which transfers a 1C unit of serine to THF to form 5,10-methylene-THF. Then, the 1C unit is transferred from one THF position to another, thus starting the folate metabolism pathway. The consensus is that the formate obtained in the mitochondria will be used in cytosolic reactions. Subsequently, in the cytosol, THF can carry fixed carbon fragments at N-5 (formimino or methyl groups), N-10 (formyl), or N5-N10 (methylene and methenyl group) bridging where each carbon oxidation state may perform different biosynthetic functions. For this reason, the cycle uses enzymes with various capacities summarized in 3 main activities: (1) reversible conversion of THF and formate to 10-Formyl-THF, (2) interconversion of 10-Formyl-THF and 5,10-Methylene-THF, and (3) interconversion of 5,10-Methylene-THF to 5,10-Methenyl-THF 23 (Table 1).
Folate cycle-related enzymes and their main characteristics.

Cytosolic Reactions
In the cytosolic compartment (Figure 2), there is a protein with triple enzymatic capacity, the MTHFD1 (methylenetetrahydrofolate dehydrogenase, cyclohydrolase, and formyltetrahydrofolate synthetase 1), whose activities are 10-Formyl-THF synthase (
The regeneration of methionine from homocysteine also involves the use of 1C units. In this pathway, 5,10-Methylene-THF can be taken up by methylenetetrahydrofolate reductase (MTHFR) to be converted to 5-Methyl-THF. 18 Methionine becomes SAM, the primary donor of methyl groups, an essential chemical modification to regulate gene expression, placing the folate cycle as a remarkable regulator of the state epigenetic. 61 In this pathway, the folate cycle is also involved in the generation of phospholipids, particularly phosphatidylcholine, synthesized through choline. SAM donates the methyl group for the subsequent reactions that will form the phospholipids. 62
The activity cyclohydrolase of MTHFD1 (
Mitochondrial Reactions
Mitochondria initially obtain folates from the cytoplasm via a mitochondrial folate transporter (MFT), 65 and serine and glycine by Sideroflexin 1 (SFXN1) and solute carrier family 25 member 38 (SLC25A38), respectively 25 (Figure 2). As mentioned above, folate must be converted to a polyglutamylated form by the enzyme FPGS, present both in the cytosol and in the mitochondria of mammalian cells, 46 allowing conversion to polyglutamylated forms in the mitochondria.
In addition to the production of one-carbon units from the conversion of serine to glycine, glycine itself is also a potential source of one-carbon units via the glycine cleavage system (GCS) in mitochondria. 66 Once the 1C unit, from either serine or glycine, has been integrated into mitochondria, it is accepted by THF to produce 5,10-Methylene-THF and enters the 1C cycle in the mitochondrial compartment.
In this compartment, as in the cytosol, 10-formyl-THF synthase (
The bifunctional enzymes MTHFD2 and MTHFD2L, encoded by the Mthfd2 and Mthfd2l gene, respectively, carry out the dehydrogenation (
The mitochondrial pathway's final step is to form formate by MTHFD1L, an NADP+-dependent synthetase (
Unlike MTHFD1, located in the cytosolic compartment, MTHFD1L only retains formyl synthetase activity and thus works in conjunction with MTHFD2/2L. 70 Compared to the 2 enzymes discussed above, MTHFD1L is expressed continuously throughout fetal development and adulthood, although it is notably higher during embryogenesis. 47 Due to the increased expression of this protein during embryogenic stages, it has been associated instead with stages of high proliferation; because of that, a great interest has been placed in the association of this protein with cancer, which has high proliferative rates as a fundamental characteristic.1,47
Folate Cycle and Cancer
Cancer cells must reprogram their metabolism to achieve and maintain uncontrolled cell proliferation, providing the energy and building blocks necessary for cell maintenance and biosynthesis. 1 Likewise, metabolic reprogramming is an essential feature of cancer cells, conferring advantages for the hyperproliferative process and counteracts the increment of ROS during the process. 2
FOLRs are overexpressed in many carcinoma cells, facilitating folate transfer into the cells. 71 As mentioned before, folate provides the 1C units necessary for the metabolic reactions of the cellular folate supply cycle. Overexpression may be essential in regulating cellular processes such as cell proliferation, as it has been associated with cancer progression and reduced patient survival.72–74 The accelerated intake of folate from the external environment probably provides the cell with more 1C units that increase some metabolic pathways. FOLR2 is associated with cell proliferation in tumors. This receptor significantly upregulated the expression of p-AKT and mammalian target of rapamycin (mTOR) phosphorylated, one of the most frequently activated signaling pathways in human cancer, and overexpression of FOLR2 associates with modulation of the tumor microenvironment (TME), particularly of the protumoral macrophages (M2).75–77 These findings support a higher incidence of cancer in patients supplementing folic acid.78,79
Several enzymes involved in folate metabolism have been implicated in cancer development. It has been reported that MTHFD1, the trifunctional enzyme plays a central role in the aggressiveness of various tumors such as colorectal tumors, nonsmall cell lung cancer (NSCLC), and HCC.80–82 Also, it has been observed that MTHFD1 translocates to the nucleus in the S-phase of the cell cycle in the breast cancer cell line MCF-7 and the cervical cancer cells HeLa for nuclear thymidylate production, 83 likewise, MTHFD1 binds in different promoter and enhancer regions, where lysin 27 in histone 3 (H3K27) acetylation is enriched, for the regulation of gene transcription. 84
The methylation pathway, also based on the integration of 1C units, is one of the tumor metabolic reprogramming pathways. Folate is required to convert homocysteine to methionine in the methionine pathway. In this process, MTR plays a central role in SAM production, and recently Sullivan et al 85 proved the implication of MTR activity and cancer cell proliferation. On the other hand, inadequate SAM production can lead to decreased methylation of CpG islands in DNA, which affects gene transcription and alters the expression of tumor suppressor genes and proto-oncogenes.86,87
However, it has been described that mitochondrial one-carbon metabolism enzymes play an essential role in cancer aggressiveness. It has been reported that serine catabolism, and subsequent formate production, often occur at a rate higher than the one-carbon demand of biosynthesis, a process referred to as formate overflow, 88 and these data indicate that there is a difference between the cells with active and inactive mitochondrial formate production. This behavior is characteristic of some cancers with an exacerbated oxidative metabolism, identifying formate overflow as a hallmark of such cancers, and cell invasion is a possible selective advantage of this mechanism. 89
In the mitochondria, the NADP+-dependent activity of MTHFD2/2L is overexpressed in various tumors such as breast cancer, colorectal tumors, lung adenocarcinoma, and HCC.52,90–95 MTHFD2/2L activity contributes to NADPH production in an amount comparable to glutamate dehydrogenase (GS). 96 Since NADPH cannot freely diffuse through lipid membranes, this cofactor is translocated to the cytosol by a mechanism dependent on the mitochondrial NADP+-dependent isocitrate dehydrogenase 2 (IDH2) to obtain isocitrate from α-ketoglutarate (α-KG) and then citrate through the reversible activity of aconitase. Citrate then is exported to the cytosol and isomerized to isocitrate to be oxidatively decarboxylated by cytosolic NADP+-dependent IDH1, producing cytosolic NADPH. 97 This mechanism is necessary to counteract cytosolic oxidative stress but also can be used to meet the requirements of Fatty acid synthase (FASN) and 3-hydroxy-3-methylglutaryl-coenzyme A reductase (HMGCoA), both enzymes fundamental for lipogenesis, a determinant process in cancer with high aggressivity.98,99
Last but not least, MTHFD1L also has been associated with poor survival in patients with HCC, colorectal tumors, bladder tumors, and tongue squamous tumors.50,100–103 The activity of this enzyme has been implicated in reducing cellular oxidative stress and nucleotide biosynthesis. 50 Bioinformatics analysis has shown MTHFD1L as a potential diagnostic and prognostic biomarker in 5 different human cancers: urothelial bladder cancer, head and neck cancer, kidney renal papillary cell carcinoma, lung adenocarcinoma, and uterine corpus endometrial carcinoma 71 ; however, it has stirred great interest about HCC in recent years.
Furthermore, due to their role in maintaining hyperproliferative cells, antifolate drugs were the first to be used to treat cancer due to their actions on cell proliferation blockade. 104 This is why understanding the association between the demands subsidized by the metabolism of folate and cancer is placed in the crosshairs of current research.43,50
Folate Metabolism and HCC
Folate-related enzymes have been observed overexpressed HCC. Starting from the cytosolic compartment, overexpression of MTHFD1 in HCC is associated with poor overall survival and shorter recurrence time, correlating with higher serum alpha-fetoprotein (AFP). 82
SHMT2, the enzyme that provides the input of 1C units to folate metabolism in the mitochondrial compartment, is upregulated in HCC. It has been shown that SHMT2 is significantly upregulated in both mRNA and protein in HCC cell lines (Huh-7, HepG2) compared to normal hepatocyte cell lines (THLE) and that inhibition of this protein reduces cell growth and tumorigenicity. 105 SHMT2 expression levels in HCC tissues correlated significantly with tumor grade, and patients with high SHMT2 expression had a worse prognosis than patients with low SHMT2 expression. 106 (p2) However, glycine is also reported as a crucial mitochondrial donor of folate-mediated 1C units in cancer cells. A recent study showed that GCS supports nucleotide biosynthesis in HCC, enhancing tumor growth and mitochondrial metabolism. 107
In addition, MTHFD2 has been associated with advanced TNM stages (Tumor, Nodes, Metastasis), tumor microemboli, metastasis, and recurrence. In addition, it has been observed that this enzyme can translocate to the nucleus and is found at sites of DNA synthesis, which could suggest a role in replication.52,108
In the case of MTHFD1L, particularly in the liver, differentiated cells are known not to have high levels of the enzyme; however, when analyzing embryonic stages of the tissue, hyperproliferative stages upregulate this enzyme, emphasizing the usefulness of this mechanism in cells with high energy demands. 47 This suggests similar scenarios of increased proliferation, such as tumor stages.
It has been reported recently that the overexpression of the Mthfd1l gene is present in various types of cancer, including cancer of the tongue, bladder, colon, osteosarcoma, esophagus squamous cell carcinoma, and HCC. 50 Lee et al 50 reported that overexpression of Mthfd1l in cancer cells depends on folate-derived NADPH, which plays an essential role in defending against oxidative stress in cancer and promoting tumor growth while decreasing patient survival, particularly in HCC. Furthermore, it has been observed that this overexpression tends to develop adverse phenotypic features and poor clinical outcomes observed in patients, even inducing immune infiltrate, and identifying macrophages, tentatively noted as M2 macrophages, which play in favor of the tumor.12,103
Cholesterol-Associated NASH-HCC
As previously stated, NAFLD and its progression to NASH is one of the main risk factors for HCC progression 109 related to more invasive and aggressive phenotypes compared to those where the risk factor includes only hepatitis B/C virus infection or toxins. 110 Hypercaloric diet consumption, such as high-fat-enriched diets, generates NASH. Several lipid species accumulate during the progression from NAFLD to NASH, such as triglycerides, diacylglycerol, free fatty acids, ceramides, and cholesterol, among others. We and others have reported that it is the type rather than the amount of lipids contributing to the transition from steatosis to NASH. 7 Some reports highlight the role of cholesterol in hepatocyte damage, NASH generation, cellular initiation, and HCC progression to a more aggressive feature.3,10,12,111 Liang and collaborators stated that an HFHC diet drives NASH and more numerous NASH-HCC tumors in a murine model, recapitulating some features found in patients with NASH-HCC tumors. 3
Recently, it was reported that Mthfd1l could be a possible marker related to a more aggressive HCC-subtype, 12 and it is stated that the overexpression of Mthfd1l is associated with a worse prognosis in HCC and poor patient survival.12,50 Mthfd1l was closely associated with more angiogenesis judged by the CD34 marker and significant infiltration of M2 macrophages in mice subjected to a high-cholesterol diet; these data were correlated with the histological presentation in human HCC samples and with HCC patients with poor prognosis. 12
These findings suggest that cholesterol consumption could be modifying the epigenome of these transformed cells through the overexpression of Mthfd1l, given its apparent relationship with the methionine cycle. On the other hand, elevated CD206, a marker of M2 macrophages, was found in the mice's HCC tissues, highlighting the importance of the relationship of tumor-associated macrophages with folate metabolism, a topic discussed below.
Retaking formate overflow, MTHFD1L, is the mitochondria's final step to formate synthesis. Through in silico analysis, we show that HCC patients without alcohol or HBV and HCV contribution had less survival when Shmt2, Mthfd2l, and Mthfd1l are overexpressed (Figure 3), 112 and it is reported that mitochondrial formate gene overexpression is related with an increment of cell proliferation. 113 Taking into account and connecting it to our results, we could extrapolate that cholesterol-associated NASH HCC has an active mitochondrial formate production and overflow, which could distinguish a new tumor subtype candidate for therapies against mitochondrial folate enzymes.

Mitochondrial formate synthesis-related enzymes induce poor survival in HCC patients. Mitochondrial formate synthesis-related enzyme overexpression is related to an active formate overflow and poor survival of hepatocellular carcinoma patients without alcohol consumption, hepatitis B, and hepatitis C virus. SHMT2, MTHFD2, MTHFD2L, and MTHFD1L. 112 Hazard ratio (HR).
A more aggressive subtype of tumors due to cholesterol overload opens the way for new nutritional and health proposals for HCC patients. Further research into molecular mechanisms leaves us to ask whether cholesterol per se promotes Mthfd1l overexpression. This questioning makes it clear that there are new intervention routes, either by modulating cholesterol levels for the tumor or by acting directly on the expression of one of the key enzymes: MTHFD1L.
Immune Surveillance in HCC and Folate Pathway Relationship
Leukocyte infiltration, particularly tumor-associated macrophages (TAM) or M2 macrophages encompassing up to 50% of tumor mass in some types of cancer, 114 provides the aggressiveness to transformed cells, playing and exacerbating roles such as proliferation, survival, invasion, migration, angiogenesis, and metastasis. 115 As mentioned, cholesterol overload induces an aggressive phenotype in HCC, triggering F4/80+/CD206+ macrophage infiltration, indicating a positive correlation between inflammation and lipids content. 12 In the same way, in ovarian cancer models, cancer cells take cholesterol from TAM in a cholesterol efflux process through ABCA1 (ATP binding cassette subfamily A member 1)/G1 transporters. 116 The current knowledge concerns whether folate metabolism, the Mthfd1l expression, formate overflow, and cholesterol-infiltrated macrophages in HCC could also play a specific role (Figure 4).
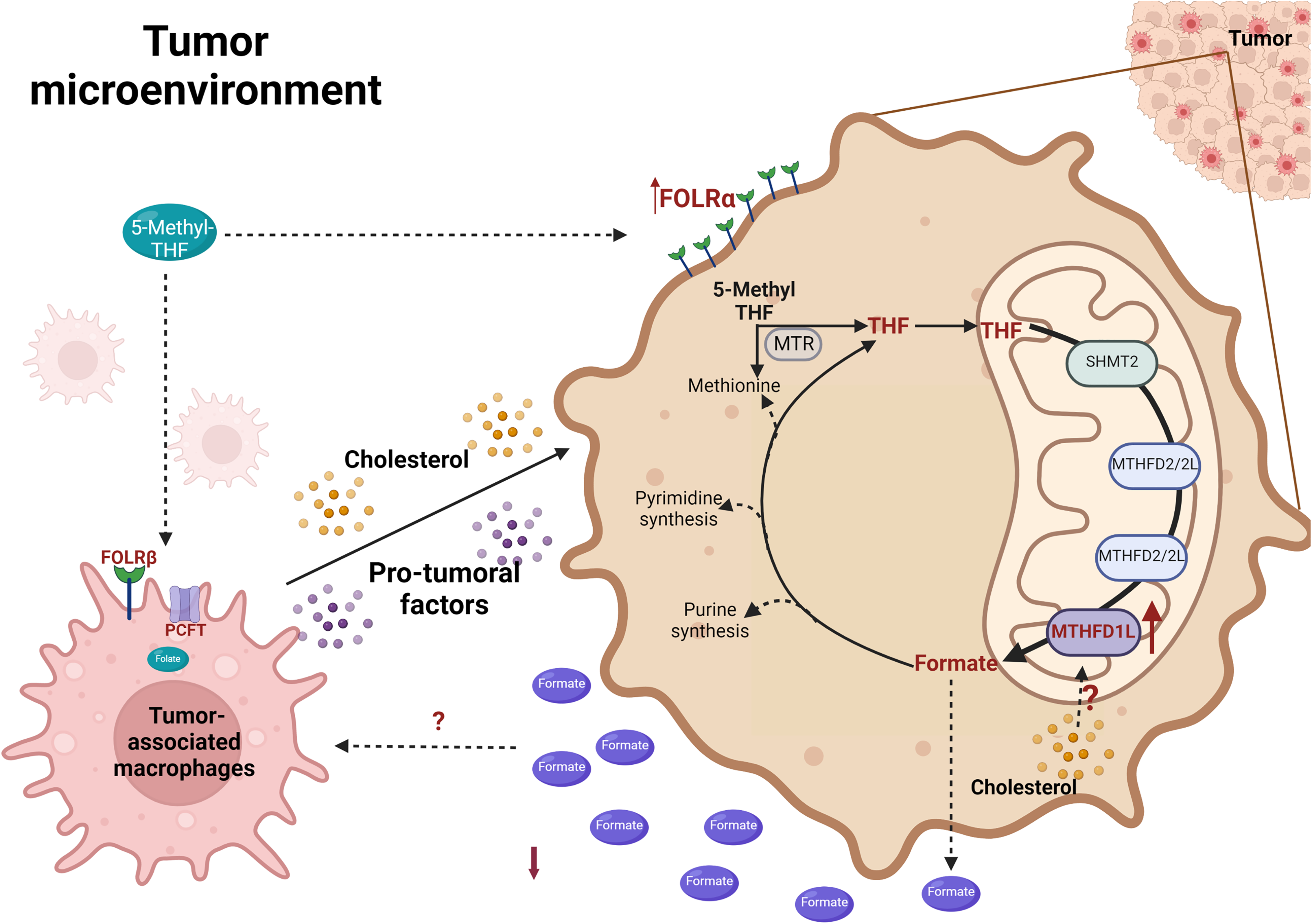
MTHFD1L overexpression modulates the tumor microenvironment—increased cholesterol levels correlated with MTHFD1L overexpression and infiltration of pro-tumoral macrophages. High availability of 5-methyl THF due to an increment of folate receptors in macrophages induces secretion of pro-tumoral factors and, in the transformed cells, induces the overexpression of a mitochondrial enzyme related to the synthesis of formate (SHMT2, MTHFD2/2L, and MTHFD1L). Folate transit and absorption co-exist in tumor cells and the TAM axis to trigger a more aggressive phenotype of HCC. Folate receptor α (FOLR α); Folate receptor β (FOLR β); methylenetetrahydrofolate dehydrogenase 1-like (MTHFD1L); methylenetetrahydrofolate dehydrogenase 28/2 like (MTHFD2/L); methionine synthase (MTR); proton-coupled folate transporter (PCFT) serine hydroxymethyltransferase (SHMT); Created with Biorender.com
The folate cycle has been involved in normal and tampered macrophage metabolism; for example, the FOLR-β is the most overexpressed transporter in these pro-tumoral phagocytes 117 and is considered an M2 and TAM marker. 118 The differential expression has been observed in activated macrophages; for instance, the folate transporter such as RFC is expressed in anti-tumoral macrophages or M1 macrophages, contrary to FOLR-β and PCFT co-expressed in M2 macrophages with major folate catchment capacity, specifically the 5-MTHF metabolite.118,119 It has been studied that 5-MTHF is a modulator of proliferation effects induced by homocysteine in human colon cancer cells such as the Caco cell line, observing cell cycle arrest in G1-G0 and S phases, with a reduction of G2-M phase that it was reflected in cell growth decrement. Similar behavior was found using folate regarding homocysteine treatment. 120 If macrophages carry out 5-MTHF uptake to decrease antiproliferative effects in tumor cells, could this represent a protection mechanism for cancer cells? And is it related to active or inactive mitochondrial formate production? Due to Activin A protein reduction, 5-MTHF uptake in macrophages could mean a witty strategy to delete M1 macrophage polarization and their cytotoxic activity.
On the other hand, folate diminished proinflammatory cytokine profiles such as C-C motif chemokine ligand 2 (CCL2), colony-stimulating factor 1 (CSF1), interleukin 1 alpha (IL1A), interleukin 6 (IL6), interleukin 10 (IL10), tumor necrosis factor α (TNF-α) particularly in M1 macrophages without any changes in M2 phagocytes.119,121 These results are important due to M1/M2 balance changes, tumor immunosurveillance reduced due to immunosuppressive M2 macrophages, and other tumor-infiltrated leukocytes (TIL) interaction with cancer cells activating a high and complexes mechanism. 122 Folate transit and absorption co-exist in tumor cells and the TAM axis to trigger a more aggressive tumor phenotype.
Taking it into account, an interesting point to highlight is that there is a strong relationship between Mthfd1l overexpression and M2 macrophage infiltration in HCC tumors. 103 Our results show that cholesterol-associated NASH-HCC also displays an overexpression of Mthfd1l and M2 macrophage infiltration, so could be a relationship between cholesterol-enriched diets, formate overflow, and M2 macrophage infiltration in HCC tumors?
Other specific enzymes related to folate homeostasis could be involved in the pathobiology of cancer and other immune diseases, such as the MTHFD1L enzyme. In cancer cell lines (786-O, A549, MCF-7, HepG2, and HCT-116), MTHFD2/2L induces immune evasion through the programmed cell death protein ligand 1 (PD-L1) up-regulation to become more tolerant against effector T cells, using c-Myc and O-GlcNAcylation via interferon-gamma (IFN-γ) secretion by T cells represented the first step to induced MTHFD2/2L activation and PD-L1 expression in cancer cells to deny the cytotoxic effect of CD8+ cells. 95 Other studies mention MTHFD2/2L as a metabolic checkpoint with other immune functions such as Treg cell-like phenotypes (FOXP3+ expression) and Th17 metabolism promotion with mTORC1 suppression and epigenetic regulation pathway. 123 Solid tumors frequently express FOXP3, including HCC.
The folate metabolism has been studied in immunity topics but has been little explored in cancer immunosurveillance, specifically in HCC. Therefore, exist a paradox in cancer immunity; leukocytes become manipulated allies instead of destroying it through a variety of mechanism.
Therapeutic Interventions
The importance of 1C metabolism in cancer was first recognized in 1948 when Sydney Farber observed that folate deficiency in the diet of children with acute leukemia reduced the number of leukemic cells; treatment in these patients with the folic acid antagonist, aminopterin, produced a temporary remission of cancer. 104
However, this drug class impacts patients’ health due to harmful side effects since folate metabolism in healthy proliferating cells is critical. Future therapies may target one-carbon metabolism in cancer cells by more selective inhibition of individual enzymes in the one-carbon pathway, 124 such as MTHFD1L. 125
Furthermore, given the significant role that cholesterol plays in developing a more aggressive subtype of HCC, future research may focus on analyzing the effects of statins on this type of tumor.126,127 Statins are small-molecule inhibitors of HMGCoAr used in the treatment of hypercholesterolemia. 128 Basic experimental studies of certain statins, such as atorvastatin, suggest that these inhibitors, which are safe in patients with NAFLD, improve NAFLD/NASH. 129 In this case, Mthfd1l overexpression can be analyzed upon treatment with statins, given the critical role cholesterol plays.
Another therapeutic option could be against these overall immunosuppressor cells and the inhibition of activity and entrance of folate forms to these cells. But why not take advantage of the infiltration of macrophages and other TILs against the tumor? Why don’t we manipulate the TME to our advantage with today's technology? Under these concepts and due to cancer cells expressing a high level of FOLR-α, for example, in ovarian epithelial carcinoma, a powerful strategy could be the use of the antibody-dependent cellular cytotoxicity (ADCC), in which immunoglobulin E (IgE) functions as crosslinking of FOLR-α in cancer cells and fragment crystallizable region ε receptor (FCεR) in macrophages, TNF-α is secreted by monocytes and MCP-1 by cancer cells. This interaction induces macrophage recruitment with subsequent cancer cell death. 130 Other evidence in triple-negative breast cancer (TNBC) shows an increment of FRα and a subsequent folate binding in transformed cells. However, strategically IgA Fc-folate conjugate could trigger polymorphonuclear (PMN) cell interaction with cancer cells due to FcαR1 expression in neutrophils. This interaction is accompanied by subsequent enzyme releases such as myeloperoxidase (MPO), which means a possible targeted therapy. 131 The same case was seen in IL-12-treated Natural killer (NK) cells using folate-conjugated to immunoglobulin G (F-IgG), Fc receptor in NK mediates Cluster of differentiation 69 (CD69) expression, an early classical marker of lymphocyte activation 132 inducing ADCC process in cancer cells including in murine leukemia tumor model. 133 Could this strategy be used in another cell type in the same folate cycle context? This is an issue pending to solve.
The folate cycle, fundamental in both normal and malignant stages, is embattled by various therapeutic targets. However, further studies and analysis of the knowledge of this cycle are essential to finding the way toward new therapeutic interventions.
Conclusion
Various enzymes throughout the folate cycle emphasize the role of folates as participants in an integrating process of cellular metabolism, in addition to alternating between the forms expressed throughout the development stage of the organism (Table 1). As mentioned, 1C units can be distributed in various cellular processes, including cellular biosynthesis, redox state regulation, epigenetic regulation, and genome maintenance. As it is known, cells require the so-called building blocks that include proteins, lipids, and nucleic acids, to maintain accelerated proliferation and to satisfy these needs through an aberrant metabolism, generating an altered, sustained, and beneficial cycle for themselves, where the folate cycle plays an important role.
Likewise, the mutation of any of the enzymes involved in this cycle can have consequences for its functioning, and in turn, for the functioning of the cell itself.
Footnotes
Abbreviations
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Statement
Not applicable.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was partially funded by a grant from the Consejo Nacional de Ciencia y Tecnología (CONACYT) and Universidad Autónoma Metropolitana Iztapalapa.