
Abstract
Oxidative stress resulted from reactive oxygen or nitrogen species in biological systems has a significant role in the diagnosis/progression of several human diseases. Human diseases associated with oxidative stress include Alzheimer's disease, chronic lung disease, chronic renal failure, cancer, diabetes, and fibrosis. In oxidative stress conditions, carbonylation process can be described as one of the most common modifications in biomolecules that takes place in the presence of carbonyl (C = O) groups which are introduced into molecules by direct metal-catalyzed oxidation of certain amino acids or indirectly by reaction with the oxidation of lipids and sugars. At a molecular cellular level, carbonylation can cause some defective biological consequences or chemical transformations in cells. During this process, specifically, carbonylated proteins can be accumulated in cells and trigger to develop some diseases in human body. The role of the accumulation of carbonylated proteins in the progression of several diseases has also been reported in the literature, such as neurodegenerative diseases, diabetes, obesity, aging, and cancer. Early detection of carbonylation process is, therefore, very critical to monitor these diseases at an early stage. Finding a suitable biomarker or probe is very challenging due to the need for multiple criteria: high fluorescence efficiency, stability, toxicity, and permeability. If they are designed with a good strategy, these probes are highly effective in cell biology applications and they can be used as good diagnostic tools for monitoring oxidative stress-induced carbonylation in relevant diseases. This review highlights the design and use of recent fluorescent probes for visualization of carbonylation in cellular systems and the relationship between oxidative stress and carbonyl species for causing long-term disease complications.
Introduction
Reactive Oxygen Species, Oxidative Stress, and Carbonylation
Free radicals are highly reactive species with one or more unpaired electron(s) in the environment and can be formed when oxygen interacts with certain molecules. These radicals can be produced in cells by losing or accepting a single electron; therefore, they can act as oxidants or reductants. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are known as reactive radical and nonradical species of oxygen and nitrogen, respectively. Reactive oxygen species and RNS can be formed during aerobic cell activities and further play a critical role in aging as well as in age-related diseases1–14 (Figure 1).

Carbonylation of biomolecules (DNA, proteins, and lipids) under stress and its causes. Photos were modified from the original article. 15
These radicals under oxidative stress environment are involved in several acute and chronic pathological processes, such as acute and chronic kidney disease, neurodegenerative diseases (NDs), macular degeneration, biliary diseases, Alzheimer, and cancer.16–19 Cardiovascular risk factors (ie, obesity, diabetes, hypertension, and atherosclerosis) are also associated with the inflammatory pathway mediated by IL-1α, IL-6, IL-8, and increased cellular senescence.20–25 Moreover, in NDs, including Alzheimer disease (AD), brain tissue samples indicate high levels of p16, MMP, and IL-6. The induction of epithelial to mesenchymal transition triggered by RONS promotes cancer metastasis.18,19,26,27 In synthesis, given the close relationship between oxidative stress, inflammation, cancer, and aging, oxidation-based aging has been explained in several ways: aging is known as homeostasis damage due to chronic oxidative stress that affects especially the human regulatory systems, such as nervous, endocrine, and immune systems. The consequent activation of the immune system induces an inflammatory state that creates a circular pathway in which chronic oxidative stress and inflammation feed each other, and consequently, increases the age-related diseases and their corresponding problems. Based on general disease progression and their existing relevant biochemical mechanisms, oxidative stress has an important role in the production of certain chemical compounds. For example, reactive lipid aldehydes that nonenzymatically alkylate cysteine, histidine, or lysine residues in a reaction termed protein carbonylation. Reactive lipid aldehydes and their derivatives are detoxified via a variety of phase I and phase II systems, and when antioxidant defenses are compromised or oxidation levels are elevated, an increase in protein carbonylation was observed. The resulting modification has been shown as a causing factor in a variety of metabolic states and pathophysiology of many diseases including neurodegeneration, muscle wasting, insulin resistance, diabetes, aging, and cancer. Although such modifications usually also result in loss of protein function, protein carbonylation may be regulatory and activate major signaling pathways involved in antioxidant biology and cellular homeostasis.5,6,8,28 In the presence of an oxidative stress environment, ROS or RNS is generated in excess and this excessive ROS/RNS attack biomolecules such as proteins, lipids, and DNA, which can disrupt cell structure and function and cause inflammation responses. Proteins are particularly taking a critical role in oxidation-based inflammation processes which are described as oxidative posttranslational modifications and occur in proteins. They are essential molecules in any cell disruption and are classified into 2 categories in the literature: irreversible oxidation and reversible oxidation; both of which can be selectively induced by ROS and RNS. Carbonylation process has been reported in proteins by many researchers more than the other biomolecules due to its known posttranslational modifications. 15 In this process, irreversible posttranslational modification is considered to trigger protein aggregation in cells and tissues and thus is a well-accepted biomarker of oxidative stress–related disorders.29–31 Three different pathways have been reported in the literature for regular carbonyl production: (i) metal-catalyzed oxidation, (ii) advanced glycation end products, and (iii) reaction with reactive lipid peroxidation products. Carbonyl products from reactive lipid peroxidation reactions are the common source of protein-bound carbonyl forms. However, lipid-bound carbonyls have been less studied and reported compared to protein carbonyls, but their reactions produce protein adducts, immune-modulating, and pro-inflammatory-based molecules. In addition to proteins, carbonylated phosphatidylcholine lipids can be recognized by scavenger receptors leading to the induction of immune response and adhesion of monocytes to endothelial cells.2,7,15,20,30,32,33
Over the past decades, fluorescent probes with different spectral and chemical properties have been designed, reported, and applied in detecting carbonylation levels in cellular systems (Figure 2). Therefore, it is critically important to specifically detect the activity of oxidative stress-associated carbonylation levels in living cells and in vivo to investigate their disease-causing factors.7,21,30,32,34–36 It is, therefore, essential to explore robust and specific detection methods to understand the multiple pathways of biomolecule carbonylation and their biological relevance in cellular systems to understand its long-term effects on disease progression. Identification of diseases at early stages in molecular level will enable us to pursue on finding solutions for early diagnosis of major diseases and help to develop personalized-drug therapies.

Representation of fluorescence labeling of carbonylated lipids, sugars, and proteins with fluorophores.
In this review, recent advances involving small molecule fluorescent probes designed to visualize oxidative stress–related carbonylation systems in cellular environment. The probes here would allow us to detect disease progression in various systems. This review will provide background knowledge for developing future new fluorescent probes which can be used in studying oxidative stress-based diseases.
Carbonyl Detection Methods
There are several biochemical and analytical methods available to identify and detect reactive carbonyls. 7 Their detection methods can be classified and used into 3 categories: (i) using biochemical and immunological techniques such as immunoblotting and ELISA, which provide general information on the modified proteins and carbonylation levels; (ii) using spectrophotometric and chromatographic fluorescent assays to determine the total protein carbonyl content; and (iii) mass spectrometry (MS) for identification of the modified proteins and relative quantification of protein-bound carbonyls. Spectrophotometric and biochemical techniques usually focus on using carbonyl-specific derivatization reagents. Here, a short overview of the most popular recent reagents is discussed with their different strategies to detect carbonyls in cellular systems. Although such labeling steps are not required for MS, they have often been used for several reasons: Modified proteins (peptides), especially in complex biological samples such as cell or plasma proteomes or peptidomes, require specific and highly sensitive methods due to their very low amounts in the assay. In addition, some carbonyl modifications can cause small mass shifts, such as aminoadipic semialdehyde formed from lysine (1 m/z unit), which are challenging to distinguish by the fast-scanning mass analyzers with low resolution. Also, some other carbonyl species are isomeric, such as glutamic semialdehyde (formed from Pro) and hydroxyproline, so isomer derivatives are much more difficult to analyze with their fragmentation patterns. Due to all these potential existing challenges, fluorescence-based carbonyl-tagging systems are preferred in current research. Some of the recent specific-targeted fluorophore tagging in different biological systems are briefly discussed in the following section:
Specific-Targeted Fluorophores
One of the oldest commercially available chemical reagents for monitoring carbonylated species (eg, ketones and aldehydes) is 2,4-dinitrophenylhydrazine (DNPH)—it is basically used for the detection of carbonylated proteins with spectrophotometric methods.37,38 Anti-DNP antibodies are widely used as a sensitive immunodetection method by using western blots or ELISA. Immunocytochemical detection and fluorescence microscopy are used to evaluate spatial distribution of carbonylated proteins. However, standard immunochemistry protocols are slow, expensive, and ending up failing due to cross-reactivity problem of the antibodies (Ab). Furthermore, DNPH-based immunochemistry essays are relatively specific and easy to process for the detection of only protein-bound carbonyls, not applicable for other carbonylated biomolecules. Fedorava and his coworkers reported a fluorophore, coumarin-hydrazide, a fluorescent chemical probe for time- and cost-efficient labeling of cellular carbonyls followed by fluorescence microscopy to evaluate their intracellular formation both in time and space.15,39 They also confirmed its specificity in time- and dose-dependent experiments using human primary fibroblasts and compared their results with conventional DNPH-based immunochemistry (Figure 3).

Left: Fluorescence labeling of carbonylated lipids and proteins in cells using coumarin-hydrazide. Right: Confocal fluorescence microscopy showing the subcellular localization of oxPL (A; control and PQ-treated) and carbonylated biomolecules (B; control and PQ-treated) alone or in co-staining experiments (C) in PQ-treated primary fibroblasts. Fixed cells were immunostained with E06 antibodies (green; A) and CHH (blue; B). For better representation of co-staining (C) E06 stain is shown in green and CHH stain red colors. Merged images (yellow) demonstrate the co-localization of oxPL with carbonyl stain. Images are representatives of 3 independent experiments performed in triplicates. Photos were modified from the original article.39
They analyzed the detection of CHH-derivatized carbonylated phospholipids (oxPL) by MS and reported their oxPL subcellular distribution. This approach was specifically focused on detecting oxidized phospholipids (oxPL) on natural Ab E06 which are on oxidized phosphatidylcholines (oxPC). They observed a drastic fluorescence increase with the fluorophore-labeled Ab E06, which showed high amounts of oxPL in primary fibroblasts after PQ treatment. Then they confirmed oxPL co-localization with carbonylated species, control, and PQ-treated cells. Confocal microscopy was used to monitor the treated cells with the fluorophore (Figure 3. right photo with label C).
In another work, Bane and her coworkers approached carbonylation labeling by designing several coumarin molecules which react with reactive carbonyls in the cellular systems. In their labeling system, the product hydrazone exhibited a wavelength shift in the emission maximum accompanied by an increase in emission intensity. 40 Spectral characteristics of the fluorophore enabled the identification of the fluorophore-conjugated cellular biomolecules in the presence of unreacted dye and eliminated the need for removal of excess dye from the environment. Moreover, they also found that this fluorophore is nontoxic and was thus suitable for live cell imaging. They also reported another coumarin fluorophore, benzocoumarin hydrazine (BzCH), which was particularly applicable for live cell analysis. In their spectroscopic analysis, the emission maximum of BzCH had a drastic red shift upon hydrazone formation (from ∼430 to ∼550 nm), which was due to fluorophore disaggregation. The hydrazone product from this reaction produced a large stokes shift (∼195 nm) (Figure 4), which made the biomolecule carbonylation detectable in cell imaging. 41 In 2021, same group developed a novel fluorogenic assay using another coumarin sensor, TFCH, that interacted with biomolecule carbonylation and efficiently detected modest forms of renal injury with much greater sensitivity than standard assays for nephrotoxins. They successfully demonstrated that this coumarin derivative could be deployed in live kidney cells and in renal tissues. 42

Benzocoumarin hydrazine (BzCH) was used as a fluorogenic sensor, showing a large stokes shift on the absorbance for detecting carbonyls in isolated biomolecules and in live cells. 41 Photos were taken from the original article with permission.
In 2020, Bowden and his group analyzed carbonylation process with a different approach using a one-pot 2 steps reaction—they used carbazate groups on commercial cellulose membrane (CM) to specifically target carbonylated proteins by hemodialysis. 43 They also discussed the possibility to use CM application as an alternative potential dialysis membrane in hemodialysis method. On the same year, Zheng et al reported a review on fluorescent probes for monitoring various carbonyl species such as formaldehyde and carbon monoxide and showed some cellular imaging applications 44 (Figure 5).

(Left) Amine-based probes to detect formaldehyde. (Right) Highly selective amine turn-off probe (NaFP) turned-on after endogenous formaldehyde detection in live cells. Photos were modified from the original article. 44
In similar research, detection of major carbonylated groups such as aldehydes and ketones was also studied by Grune's group. They designed a new fluorometric assay with rhodamine B hydrazide (RBH), which analyzed specifically ultrasensitive quantification of total protein carbonyls. 45 Their spectral characteristics for the RBH-based essay indicated that a protein carbonyl-RBH hydrazone product exhibited a turn-on fluorescence of (ex/em 560/585 nm) and was greatly enhanced by guanidine-HCl (Figure 6).

Fluorometric assay with rhodamine B hydrazide and its turn-on hydrazone product. Photo was modified from the original article. 45
Same group reported another interesting detection method of ntrDNPH assay (commercially available molecule) which determined carbonyl groups on cell wall polysaccharides, and thus this approach might allow scientists to investigate cell walls acting as antioxidant defense in plants, fungi, bacteria, and lichens 37 (Figure 7).
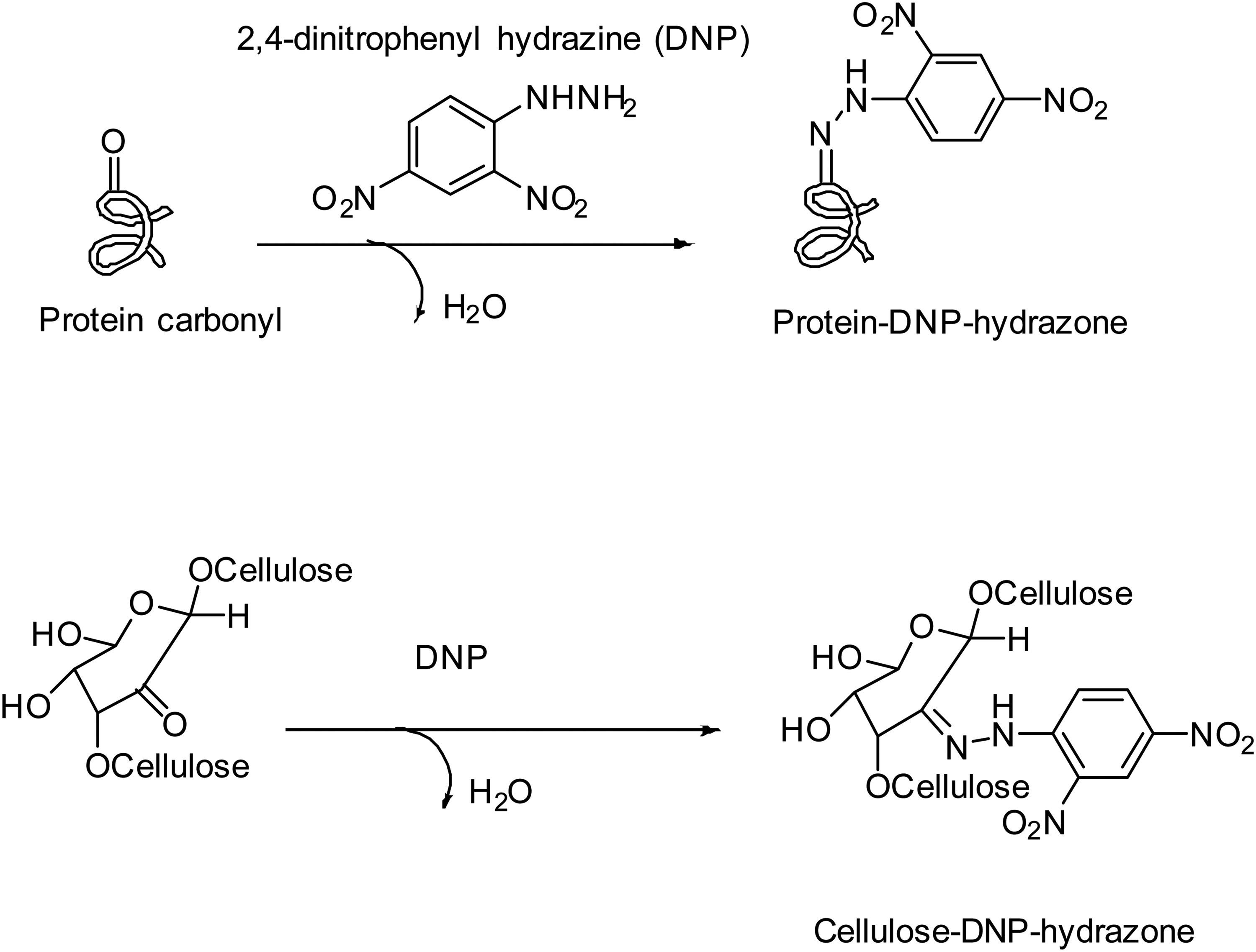
DNP for detection of cellulose carbonyls. Photo was modified from the original article. 37
Another novel fluorescence-based application was reported by Rustad et al in 2019, the group developed a simpler, faster, and cheaper method to determine carbonylated protein level in muscle foods, including both soluble and insoluble protein fractions by using a fluorophore, 7-(diethylamino) coumarin-3-carbohydrazide (CHH). 46 This novel technique labeled both soluble and insoluble carbonylated proteins with CHH and analyzed carbonyl content by fluorescence microscopy assay. This approach was a promising candidate for carbonyl detection method that could also be used as a potential conventional method like ELISA. In terms of hydrazide derivatives, Ros and his group developed a protein carbonyl detection method based on fluorescent BODIPY, Cy3, and Cy5 hydrazides.47,48 One of the advantages of this method was Western blot and immunodetection methods were not necessary to use. Method itself was quick and gave high accuracy rates. A combination of Cy3 and Cy5 hydrazides provided multiplexing analyses in a single 2-dimensional gel. Derivatization with BODIPY hydrazide allowed easy matching of the spots of interest. This method was another effective monitoring system for detecting protein carbonylation in protein samples referred to metal-catalyzed oxidation “in vitro” (Figure 8). 48

Left photo: Use of fluorescent BODIPY hydrazides for detection of oxidized proteins. BODIPY-Hz derivatization was used for accurate matching of the spots and identification. Right photo: Extracts obtained from hydrogen peroxide-treated cultures were derivatized with Cy5-Hz and BODIPY-Hz and analyzed by 2D-gel electrophoresis. Both samples (20 μg each) were mixed and analyzed on the same 2D-gel system. The images shown correspond to: A, Cy5 channel; B, BODIPY channel; C, overlay from images A and B. In D, matching between BODIPY signal and protein stain is shown. Photo was taken from the original article with permission. 48
In another work, Ahn et al also designed an alternative method for monitoring the presence of carbonyl groups based on the derivatization with fluorescein isothiocyanate. 49 With this method, the fluorescently labeled proteins (thiosemicarbazones or hydrazones) were further separated with lithium dodecyl sulfate-polyacrylamide gels by using fluorescence. The main disadvantage of this method was the instability of fluorescently labeled proteins at room temperature. In a relevant work, Daneshvar and his group reported a fluoresceinamine and sodium cyanoborohydride molecule for detecting glutamic and aminoadipic semialdehydes by the reductive amination method. 50 However, the resulting products easily degraded to nonfluorescent decarboxylated derivatives by acid hydrolysis. In 2020, Dilek and her group reported a design of a new fluorescent molecule, 2Hzin5NP, which analyzed oxidative stress–induced carbonylation in different live renal cancer cell lines. 51 These synthetic probe designs were very helpful to determine disease prognosis studies via bioorthogonal chemistry which was also reviewed recently by Wu et al, Devaraj, and Li.44,52–60 Recently, James and coworkers published a review article discussing the development of new probes that were used for targeting of organ related processes in vitro and in vivo. 61 This review covered different strategies from how to design a promising molecule to how to use them for labeling in biological systems and also highlighted the main pitfalls and limitations of these different systems in the imaging field.
Understanding the relationship between oxidative stress and carbonyl species for causing long-term disease complications
Several fluorescent probes have been designed and used to understand the relationship between oxidative stress with carbonyl production as summarized above. Increasing oxidative stress is linked with protein carbonylation which is associated with a variety of metabolic diseases, syndromes, and conditions, including aging, diabetes, heart disease, and neuropathies21,22,24,27,35,62–69 (Figure 9). One of the recent studies focused on adipose oxidative stress that was counted as a major contributor to insulin resistance and cellular dysfunction and was regulated in an age- and depot-specific manner. 7 Adipose oxidative stress varied with age in both humans and experimental mice and changes between depots. Hauck et al reported that depot and age-regulated accumulation of ROS was related to adipocyte biology and had a critical fundamental role in signaling, fat cell metabolism, and dysfunction and it potentially will have significance in clinical cases such as type 2 diabetes and obesity. 59 In this work, the accumulation of ROS in the visceral adipose depot of obese insulin-resistant mice was analyzed with insulin-sensitive mice and humans and they proved that inflammation triggered oxidative stress (Figure 10). Whether this observation showed that inflammatory cells themselves were the source of ROS or that cytokine-dependent downregulation of adipocyte antioxidants, but these results were not still conclusive to confirm it. To understand the basis of underlying problems of these diseases, increasing oxidative stress levels in chronic renal failure could also be counted as evidence of increased lipid, carbohydrate, and protein oxidation products in plasma and cell membranes.
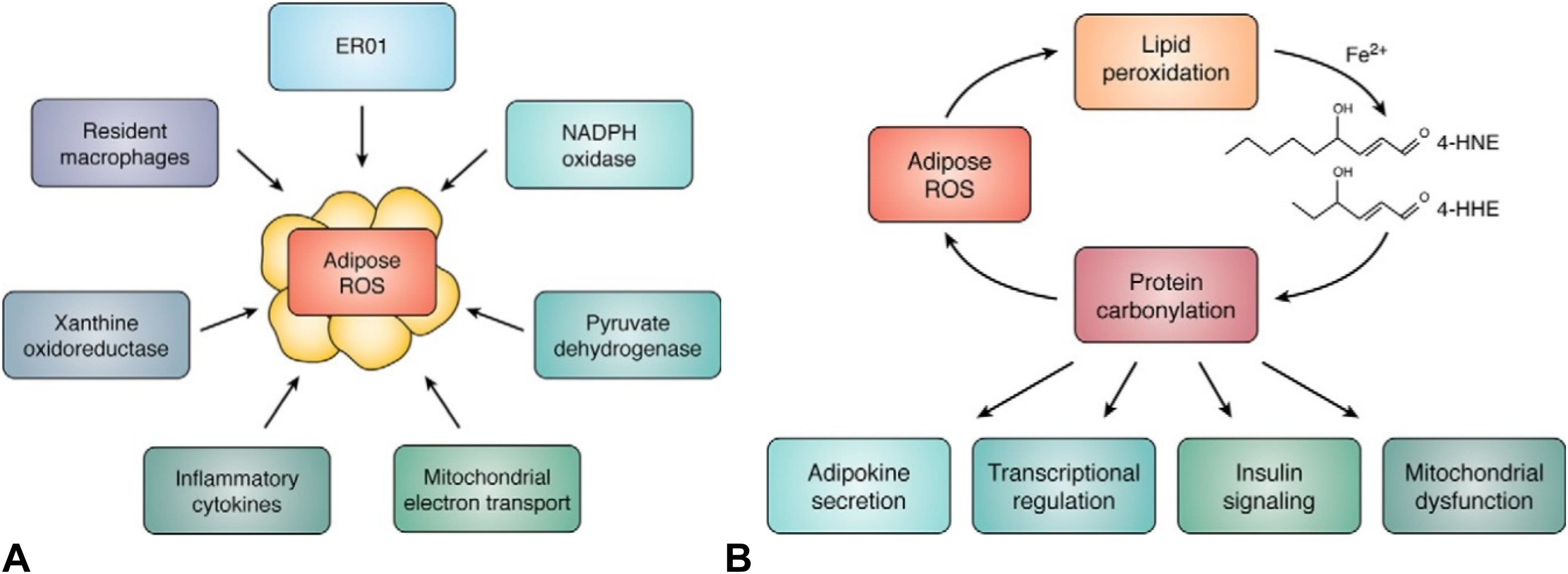
A, Enzymatic system chart for oxidative stress. (B) Reactive oxygen species (ROS) carbonylation cycle: Increased adipocyte ROS production cause lipid hydroperoxides and additional reactive lipid electrophiles such as 4-HNE and 4-HHE. Reactive lipid aldehydes in turn covalently modify proteins of the adipocyte in the mitochondrion and nucleus to regulate major metabolic processes. Photo was taken from the original article with permission. 7
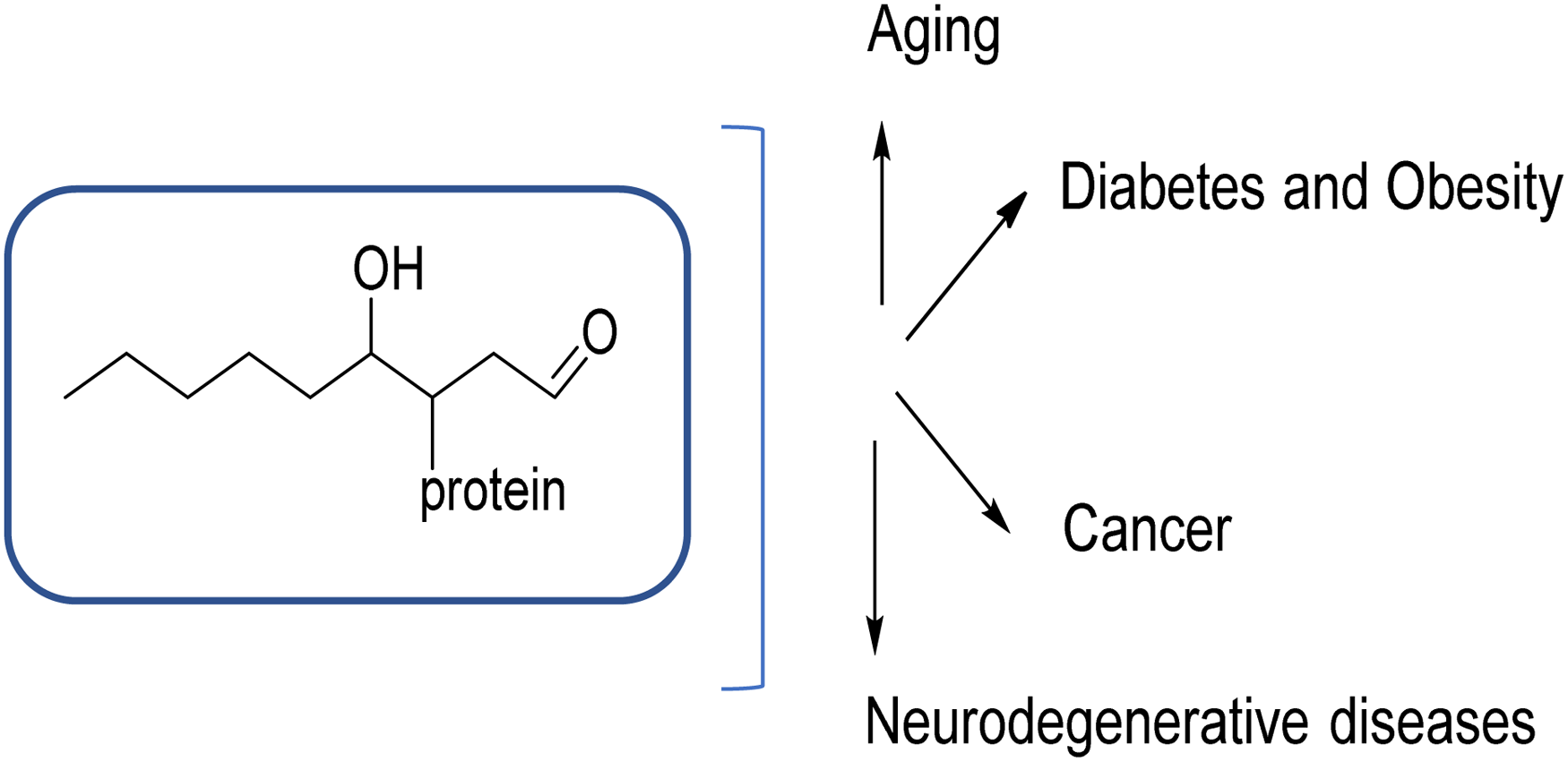
Protein carbonylation occurs in a variety of metabolic diseases. Photo was modified from the original article.
Reactive Oxygen Species–Dependent Cases
Recent studies have indicated that oxidative stress in nonenzymatic biochemistry leading to irreversible protein modifications, thus ROS may directly alter proteins with the eventual formation of oxidized amino acids. In addition to proteins, reactive carbonyl compounds formed by the oxidation of carbohydrates and lipids might indirectly lead to advanced glycation or lipoxidation of proteins. 16 For example, chronic uremia was observed with increased modification of protein caused by reactive carbonyl compounds derived from both carbohydrates and lipids and carbonylation-based oxidative stress end products, glycoxidation, or lipoxidation products, or any glycation end products, were produced from the reactive carbonyl compounds64,70 (Figure 11).

Structure of reactive carbonyl compounds derived from carbohydrates, ascorbate, lipids, and amino acids. Photo was remodified from the original articles with permission (Miyata et al., 1999 and 2000). 16
In another study, Murata and his coworkers analyzed the relevance of oxidative stress in carbonylation with ND model. Their findings showed that the characterization of specific carbonylated proteins indicated that oxidative damage could cause HSPs to misfunction in neurodegenerative and inflammation-related cancer diseases. 20 They also mentioned that in NDs, DNA damage might not only play a role in the induction of apoptosis but also might inhibit cellular division via telomere shortening. With a similar perspective, Chen et al recently published an article on the role of oxidative stress in Parkinson's disease (PD). 71 Chang and Chen also worked on the PD-causative mutations of SNCA, PRKN, PINK1, DJ-1, LRRK2, FBXO7, and ATP13A2 and found that these mutations have a significant role in oxidative stress in the pathogenesis of PD. 4 According to their study, the molecules involved in oxidative stress, such as DJ-1, coenzyme Q10, uric acid, 8-hydroxy-2′-deoxyguanosin, homocysteine, retinoic acid/carotenes, vitamin E, glutathione peroxidase, superoxide dismutase, xanthine oxidase, and products of lipid peroxidation, could be candidate biomarkers for PD.
Aging, Inflammation, and Cancer
Balducci group studied oxidative stress by studying from the point of aging and cancer. 72 They evaluated the multiple aspects that distinguish the aging process. They ended up evaluating the aging theory with a different process: according to their reviews, there was no single biomarker to date that has been developed able to identify the stage of this process. At the same time, clinical clusters have been added to find possible effective biological markers. It, therefore, finding a powerful biomarker to understand the aging process depended on several factors in human's life. Indeed, the pathways, described as potential biomarkers of aging, were strongly linked to inflammation and cancer. In another chronic experimental approach, Pecorelli et al described oxidative stress-relevant diseases by focusing on the aspect of chronic, low-level subclinical inflammatory response. 25 They proposed that the term “OxInflammation” could be described and used as a permanent pro-oxidative feature. According to their findings, nothing clinically detectable was observed in an inflammatory process that could cause the chronical problem to systemic/local damage, so all of them are regulated between inflammatory and oxidative stress mediators. To understand the deep relationship of inflammation-induced protein carbonylation with cholangiocarcinoma (CCA), Murata and coworkers ran experiments via Fenton reaction to determine the carbonylated serotransferrin-associated iron accumulation effects during oxidative stress, and they found that carbonylation of HSP70.1 with antioxidative property and A1AT with protease inhibitory capacity might lead to CCA progression. 20 Saha group also studied the relationship between nutrition, oxidative stress, and cancer initiation and examined the impact of nutrient-mediated regulation of antioxidant capability against cancer therapy. 19 In their review, they discussed in detail that cancer initiation might be triggered by the nutrition-mediated increase in ROS levels, which could stimulate cancer initiation by leading further DNA damage, and pro-oncogenic signaling (Figure 12).

(Left) Nutrients mediate oxidative stress at the metabolic tissue level. (Right) Nutrients act as cancer suppression at the molecular level. Photo was taken from the original article with permission. 19
Recent studies supported the concept that oxidative damage could occur to proteins during oxidative stress. Aging was very selective, and the proteins with specific sensitivity to oxidation could regulate cellular signaling events, including apoptosis. Magi et al published their results on the effects of oxidative stress on photodynamic therapy (PDT) evoking response. This process was likely to produce drastic changes in mitochondria and ER chaperons, which indicated that oxidative stress induced by PDT evoked a response affecting mitochondria, which allowed them to gather more information on the mechanism. 73 In another study, Rossner et al investigated plasma protein carbonyl levels in women. This group found that women with higher levels of plasma protein carbonyl and urinary 15F(2t)-isoprostane had an 80% increase in breast cancer risk compared to women with levels below the median for both markers of oxidative stress. 74 In summary, their results suggested that increased plasma protein carbonyl levels might be associated with high breast cancer risk. Other groups also studied protein carbonyl levels in tissues. For example, Aryal and his coworkers reported that 3 specific proteins of interest for carbonylation in human breast tissues; filamin A, heat shock protein 90β (HSP90β), and bifunctional glutamate/proline-tRNA ligase (EPRS), which were selectively carbonylated in tumor tissue compared to matched adjacent healthy tissue. 75 Correspondingly, they observed an increase in filamin A and EPRS protein carbonylation compared to noncancerous MCF-12A. They also confirmed low level of total super oxide dismutase activity and high autophagy. However, they did not see any increase in HSP90β protein carbonylation. According to these results, finding the selectively carbonylated proteins and defining their roles in cancer progression might be the key factor for the development of targeted therapeutic experimentations.
In a recent study, Zamyatyin et al also described and confirmed several crucial processes which were responsible for the regulation of cellular aging, including the role of carbonylation and its effects in perturbing proper protein folding, and its potential value as an aging biomarker. 63 They particularly discussed the proteins which could be used as aging biomarkers, and the recent techniques (such as MS and ELISA) could provide precise identifications of protein modifications. However, the specific proteins are the most sensitive to oxidation in different pathologies and oxidation caused an increase in carbonylation which took critical roles in various diseases.
From a different perspective, one of the critical approaches to monitor oxidative stress–induced carbonylation processes was to use a novel fluorescent probe to tag carbonylation in live cancer cells. 51 Dilek group published a relevant work that described the turnoff and fluorescence strategy (Figure 13) to understand the relationship of oxidative stress–induced carbonylation levels with cancer signaling pathways in renal cancer lines (Figure 14).

(Left) Synthesis and design of hydrazine-based fluorescent probe to label carbonyl species in live cells. (Right) (A) Absorption spectra of 2Hzin5NP and its hydrazone products in methanol. From left to right, 2Hzin5NP (blue), aliphatic hydrazone; 2 (red), aromatic hydrazone; 3 (green). (B) Emission spectra of compounds in methanol. 2Hzin5NP (blue, λex = 354 nm, λem = 469 nm), 2 (red, λex = 397 nm, λem = 502 nm), and 3 (green, λex = 418 nm, λem = 517 nm). (C) Solutions of 2Hzin5NP, its aromatic hydrazone in methanol under room light (left) and long wavelength fluorescent light (right). (D) Solutions of 2Hzin5NP, its aliphatic hydrazone in methanol under room light (left) and long wavelength fluorescent light (right). 51

Detection of serum starvation–induced carbonylation in HDF, A498, and ACHN cells. Cells were incubated with and without 10% FBS in DMEM for 16 h. Although A498 cells were labeled with 20 µM, ACHN and HDF cells were labeled with 15 µM 2Hzin5NP for 30 min. (A) Representative images from 3 independent experiments were captured using Zeiss LSM 800 confocal microscope at 40× objective. 405 nm and 488 nm diode lasers were used for excitation and LP 435 and 518 filters were used for the emission. Scale bar is equal to 10 µm. (B) Quantitative analysis of carbonylation levels in HDF, A498, and ACHN cells using Varioskan Multimode Plate Reader. Cell lysates obtained by 6 freeze-thaw cycles in lysis buffer were analyzed by measuring fluorescence intensity (RFU) at 396 nm excitation and 506 nm emission. The autofluorescence intensity of control groups was, respectively, subtracted from all experimental groups. Each data point represented average of at least 3 independent experiments. The percentage of RFU obtained for each 2Hzin5NP labeled cell line in the serum-free conditions was set to 100%. (C) Detection of carbonylation levels in HDF, A498, and ACHN cells in response to serum starvation. Each data point corresponded to average RFU from 3 independent experiments. *P ≤ .05, **P ≤ .01, ***P ≤ .001. 51
Neurodegenerative and Other Chronic Diseases
According to Milzani group, they specifically discussed protein carbonylation-related human diseases which included AD, chronic lung disease, chronic renal failure, diabetes, and sepsis in their article. 36 They evaluated recent progress in the identification of carbonylated proteins which could act as new diagnostic (possibly presymptomatic) biomarkers for oxidative damage and produced fundamental information to determine the possible effective antioxidant therapy. They basically examined select diseases in which elevated levels of PCO (polycystic ovarian) and concluded as these elevations correlate well with the progression or severity of the disease. Herwig and coworkers also reported protein carbonylation-based clinical study. They assessed whether carbonyl proteins as end products of metabolic processes might act as a marker for oxidative stress in the cerebrospinal fluid (CSF) of patients with NDs. 76 Their clinical trial set results showed that carbonyl proteins might serve as an indicator for oxidative stress in the CSF of relapsing as well as progressive (multiple sclerosis) MS patients and in patients with NDs. In relevance to MS, Irani group focused on only MS patients and took CSF samples to analyze carbonylated protein levels inside and found that carbonylation proteins were elevated compared to control samples. 77 They confirmed that in their experimental autoimmune encephalomyelitis, carbonylated protein levels in CSF correlated closely with those found in inflamed spinal cord tissues.
Butterfield and coworkers covered and compared the study of carbonylated proteins by using immunological techniques with proteomic analytical methods. 27 Analyzing carbonylated proteins using 2-dimensional gel electrophoresis, followed by immunoblotting and MS was producing rapid processing than the conventional methods. They also used proteomics approaches for the identification of carbonylated proteins in various diseases in humans, animal, and cell models and have provided critical information to cell biologists by describing the effects of modifications by carbonyl species on protein function at a cellular level. To improve rapid detection methods for functional protein networks, Butterfield and coworkers developed a proteomics perspective to identify specifically oxidized proteins in AD by detecting carbonylated proteins.
In addition to neurological diseases, carbonylation relationship with other chronic metabolic diseases has been also reported by numerous research groups.7,16,22,35,78–82 Bernlohr et al studied the metabolic changes associated with obesity and the development of insulin resistance. 7 They focused on the oxidative stress level interaction with the inflammation status at both the tissue and organismal level. Obesity-related oxidative stress has been found as a contributing factor to the increase of insulin resistance in the adipocyte and myocyte. They also found that oxidative stress–triggered protein carbonylation in the metabolic defects had a determining role in mitochondrial dysfunction. In another oxidative stress–based disease case, Bane group recently published an article on an efficient fluorogenic diagnostic assay that was used for the detection of biomolecule carbonylation for renal injuries. 42 Their assay would help to identify and analyze diverse pathologies, which would not only allow clinicians to make early diagnoses but also help them to understand the basis of these pathological results.
Conclusions and Future Perspectives
Biomolecule carbonyls (lipids, DNA, or proteins) are usually known as important biomarkers which can be used in the early diagnosis of pathologies associated with the overproduction of ROS during oxidative stress; therefore, robust, and specific methods to quantify the protein carbonyls in complex biological samples are much needed. To find the best method or biomarkers/probes for investigating the role of carbonylation in oxidative stress/ROS-based pathogenesis and/or progression of human diseases is challenging. Although there is considerable evidence of the involvement of oxidative stress leading to biomolecule carbonylation in many diseases, there are still many questions that need to be explored about how oxidative stress is involved mechanistically and chemically in several diseases. For example, one of the challenges in oxidative stress research today is to identify the specific oxidized protein diseases. During oxidative stress and damage, it is possible to lose the specific protein functionality and this could lead to inconclusive imaging results if the cells are labeled with the probes. The relationship between protein oxidation and corresponding diseases is therefore still unclear.
The imaging studies of oxidative stress concept and its dynamics have the potential to contribute to the understanding of important pathways involved in the regulation of cellular functions and metabolism. Especially, new upcoming imaging technologies in terms of developing selective fluorescent probe systems, bioconjugation, and genome engineering tools will make innovative approaches toward a broader understanding of the role(s) of oxidative stress and protein carbonylation in human metabolic diseases.
The current rapid progress in the detection of carbonyls in biomolecules should open a new insight to the imaging research world. Especially small fluorescent molecules having suitable photophysical and chemical properties can be designed as fluorophores for use them in human diseases that are associated with oxidative stress so that these robust assays may provide key information to the next step of clinical studies. For example, the detection of selective and rapid performing small molecule–based imaging technologies to target selectively protein carbonyls in tissue environment or in vivo might be the potential future imaging approach to understand the underlying metabolic signaling pathways during oxidative stress.
The other possible challenges for the future in this area are to elucidate the chemical mechanisms that lead to disease-associated oxidative stress, to develop effective drug therapies. However, before developing any drug therapies, determining the effective fluorescent imaging agents to diagnose on selective targeting of the target biomolecules that triggers or associates with oxidative stress would be the key element in this process. The completion of these studies will open a new insight in diagnosis and therapy of human diseases associated with oxidative stress.
Caveats
The probes/fluorophores and protocols here are depending on the kinetics of the probes with the target molecule and fluorescence efficiency of the probe in the cells. Several presented probes are much slower than the others due to their specific functional groups. For example, hydrazide-containing reagents are very slow to react with aldehydes and ketones at physiological pH, 83 but these reaction rates are increased by orders of magnitude when they are in cellular environment. On the other hand, some of the designed probes also have fast kinetics with the target biomolecule but show fluorescence quenching when they go into cells. Sometimes either no spectral (absorption or emission) shift is observed on the spectra or no fluorescence increases when the fluorophore binds to the target molecule, so this may cause a serious imaging problem when the probes are used in cells. In addition to spectral properties, solubility, and chemical characteristics of the fluorophores in aqueous environment may also lead to some problems in imaging because some fluorophores do not fluoresce when they are in very polar solutions. If these fluorophores are used in vivo or in living cell conditions, fluorescence intensity is an essential parameter that can be used to determine the concentration of the target in the fluorescence-based detection method. If the fluorescence quantum yield upon ligand binding is low or if there is too much background fluorescence appears due to existing excess unbound fluorophore, this situation can complicate the calculations of ligand-bound system and cause some errors in imaging.
Footnotes
Abbreviations
Acknowledgements
The authors would like to thank University of the District of Columbia for providing me all the institutional support.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.