
Abstract
Objective:
This study aimed to investigate the effect of tetra-arsenic tetra-sulfide on treating multiple myeloma and its potential regulation on suppressor of cytokine signaling 1 methylation-mediated Janus kinase 2/signal transducer and activator of transcription 3 signaling pathway.
Methods:
Tetra-arsenic tetra-sulfide with different concentrations were used to treat U266 cells, and cell viability was measured at 12, 24, and 48 hours with 0 μM tetra-arsenic tetra-sulfide treatment as control by Cell Counting Kit-8 assay. Suppressor of cytokine signaling 1 methylation and expression were determined by methylation-specific polymerase chain reaction, quantitative polymerase chain reaction, and Western blot, respectively, in U266 cells and normal plasma cells and in U266 cells treated by tetra-arsenic tetra-sulfide. Then, rescue experiments were performed by transfecting suppressor of cytokine signaling 1 small interfering RNA into tetra-arsenic tetra-sulfide-treated U266 cells. Besides, phosphor–Janus kinase 2, Janus kinase 2, phospho–signal transducer and activator of transcription 3, and signal transducer and activator of transcription 3 expressions were determined by Western blot.
Results:
Tetra-arsenic tetra-sulfide inhibited U266 cell viability efficiently in a dose- and time-dependent manner. Suppressor of cytokine signaling 1 methylation was higher while suppressor of cytokine signaling 1 expression was lower in U266 cells compared to normal plasma cells; when treated by tetra-arsenic tetra-sulfide, suppressor of cytokine signaling 1 methylation was decreased while suppressor of cytokine signaling 1 expression was increased in U266 cells, along with the reduced phospho–Janus kinase 2 and phospho–signal transducer and activator of transcription 3 expressions. Then, suppressor of cytokine signaling 1 small interfering RNA enhanced the cell viability and phospho–Janus kinase 2 as well as phospho–signal transducer and activator of transcription 3 expressions in both tetra-arsenic tetra-sulfide treatment-free and tetra-arsenic tetra-sulfide-treated U266 cells.
Conclusion:
Tetra-arsenic tetra-sulfide exhibits good killing effect on multiple myeloma cells via repressing suppressor of cytokine signaling 1 methylation and downstream Janus kinase 2/signal transducer and activator of transcription 3 signaling pathway, which might serve as a potential treatment option for multiple myeloma.
Introduction
Multiple myeloma (MM) is a plasma cell neoplasm characterized by clonal proliferation of malignant plasma cells and accompanied with symptoms such as extensive skeletal destruction, infections, anemia, hypercalcemia, and renal failure, which accounts for around 1% of neoplastic diseases and 13% of hematologic malignancies. 1 -3 Despite the therapeutic landscape of MM evolved spectacularly over the past decades, such as the application of autologous stem cell transplantation, advances in supportive care, and the use of novel drugs, most patients eventually relapse, and MM remains a fetal disease. 4 -7 Hence, a deeper understanding of the pathophysiology of this disease and the development of a novel drug with better treatment efficacy are still needed for MM.
Tetra-arsenic tetra-sulfide (As4S4), a major constituent of realgar, has been applied widely for acute promyelocytic leukemia (APL) treatment and present with low toxicity as well as good therapeutic efficacy; moreover, it has shown potential clinical application in chronic myelogenous leukemia (CML) when combined with other agents. 8 -10 For the molecular mechanisms of As4S4, previous studies have shown that As4S4 is able to inhibit the expression of multiple cancerogenic factors (such as SET protein, ribosomal protein P2, and prohibitin) to induce the accumulation of cellular reactive oxygen species (ROS) and upregulate the expression of the p53 tumor suppressor, which promotes mitochondria- and endoplasmic reticulum stress–mediated cell apoptosis in various hematological malignancies such as acute lymphoblastic leukemia, APL, and CML. 11 -14 Notably, our preliminary experiment found that As4S4 had a repressive effect on suppressor of cytokine signaling 1 (SOCS1) methylation. And some studies have disclosed that SOCS1 hypermethylation is observed in MM patients, meanwhile an in vitro experiment shows the effect of SOCS1 demethylation on promoting MM cell apoptosis, indicating that targeting SOCS1 methylation may be a potential approach for treating MM. 15,16 Furthermore, Janus kinase 2/signal transducer and activator of transcription 3 (JAK2/STAT3) signaling pathway is a downstream signaling pathway regulated by SOCS1, and 2 previous studies have revealed that As4S4 may exert negative regulation on JAK2/STAT3 signaling pathway. 17,18 Therefore, considering As4S4 has good antitumor effect in hematological malignancies, and As4S4 has been found to regulate the methylation and the downstream signaling pathway of SOCS1; furthermore, targeting SOCS1 methylation may facilitate MM cell apoptosis, and we speculated that As4S4 might have killing effect in MM cells through targeting SOCS1 methylation and JAK2/STAT3 signaling pathway; however, related evidence is seldomly reported.
Therefore, the aim of the present study was to investigate the effect of As4S4 on treating MM and its potential regulation on SOCS1 methylation-mediated JAK2/STAT3 signaling pathway.
Methods
Cell Culture
The human MM cell line U266 was purchased from Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures GmbH (Braunschweig, Germany) and was cultured in 90% Roswell Park Memorial Institute (RPMI) 1640 Medium (Gibco, Gaithersburg, Maryland, United States) supplemented with 10% fetal bovine serum (Gibco). Cells were maintained in a humidified atmosphere of 95% air and 5% CO2 at 37°C.
As4S4 Sensitivity Detection
Tetra-arsenic tetra-sulfide (MuseChem, Fairfield, New Jersey, United States) was dissolved in 0.1 M sodium hydroxide to make a stock solution of 40 mM. Stock solutions were diluted in RPMI 1640 medium before using. The U266 cells were treated with 0 μm (served as a control), 5, 10, and 40 μM As4S4 for 12, 24, and 48 hours, respectively, and then cell viability was detected by Cell Counting Kit-8 (CCK-8) assay. A growth curve was drawn by CCK-8 colorimetry. Relative cell viability was calculated by optical density (OD): [OD of treated/OD of control] × 100%. IC50 (the concentration inhibiting 50% of in vitro cell growth) was calculated according to the relative cell viability.
SOCS1 Methylation, Messenger RNA Expression, and Protein Expression in U266 Cells
In U266 cells, the methylation of SOCS1 was detected via methylation-specific polymerase chain reaction (MSP), the messenger RNA (mRNA) expression of SOCS1 was detected by reverse transcription–quantitative polymerase chain reaction (RT-qPCR), and the protein expression of SOCS1 was detected using Western blot. Plasma cells isolated from the bone marrow sample of the healthy donor by CD138-coated magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany) were used as control. And the methylation, mRNA expression and protein expression of SOCS1 were also detected in control cells. Our study was approved by the ethics committee of our hospital (approval no. XJTU1AF2017LSK-138). All patients provided written informed consent prior to enrollment in the study.
Effect of As4S4 on Methylation, Expression of SOCS1, and JAK2/STAT3 Pathway
The U266 cells were incubated with 10 μM As4S4 for 12, 24, and 48 hours. After incubation, MSP was carried out to detect the methylation of SOCS1; RT-qPCR and Western blot were performed to detect the mRNA and protein expressions of SOCS1. In addition, Western blot was also conducted to determine the protein expressions of JAK2, phospho-JAK2 (pJAK2), STAT3, and phospho-STAT3 (pSTAT3).
Transfection and Subsequent Detections
Suppressor of cytokine signaling 1 small interfering RNA (siRNA) was purchased from Invitrogen (Carlsbad, California, United States) and the sequence was 5′-CACTTCCGCACATTCCGTT-3′. The control siRNA was also provided by Invitrogen. The SOCS1 siRNA and control siRNA were transfected into U266 cells using Lipofectamine (Invitrogen) according to the manufacturer’s instructions. The cells transfected with SOCS1 siRNA and control siRNA were named as si-SOCS1 group and negative control (NC) group, respectively. At 48 hours after transfection, the SOCS1 mRNA and protein expressions were detected by RT-qPCR and Western blot; the protein expressions of JAK2, pJAK2, STAT3, and pSTAT3 were determined by Western blot; the cell viability was detected by CCK-8 assay.
At 48 hours after transfection, cells in NC and si-SOCS1 groups were incubated in RPMI 1640 medium containing 10 μM As4S4 (MuseChem) for 24 hours. Then cells were divided into As4S4 group and si-SOCS1 + As4S4 group. After incubation, RT-qPCR and Western blot were carried out to detect the SOCS1 mRNA and protein expressions. And Western blot was performed to detect the JAK2, pJAK2, STAT3, and pSTAT3 protein expression. Cell Counting Kit-8 assay was used to detect the cell viability.
Methylation-Specific Polymerase Chain Reaction
DNA was extracted using a DNA purification kit (Boehringer, Mannheim, Germany), and the DNA concentration was measured with the use of a spectrophotometer. The DNA was denatured with sodium bisulfite using CpGenome DNA Modification Kit (InterGen, Edinburgh, United Kingdom). Polymerase chain reaction (PCR) was performed using EpiTect Whole Bisulfitome Kit (Qiagen, Hilden, Germany). The MSP reaction conditions were: 95°C for 5 minutes, then 95°C for 30 seconds, 60°C for 45 seconds, and 72°C for 1 minute for 40 cycles. The SOCS1 primers were as follows: methylation: 5′-TTCGCGTGTATTTTTAGGTCGGTC-3′ (forward) and 5′-CGACACAACTCCTACAACGACCG-3′ (reverse); unmethylation: 5′-TTATGAGTATTTGTGTGTATTTTTAGGTTGGTT-3′ (forward) and 5′-CACTAACAACACAACTCCTACAACAACCA-3′ (reverse). The PCR product was detected by 1.5% agarose gel electrophoresis.
Reverse Transcription–Quantitative Polymerase Chain Reaction
Total RNA was extracted using RNeasy Protect Mini Kit (Thermo Fisher Scientific, Massachusetts, United States). Then, with the application of PrimeScript RT Master Mix (Takara, Kyoto, Japan), transcription to complementary DNA was performed. Polymerase chain reaction procedure was conducted using TB Green Fast qPCR Mix (Takara, Kyoto, Japan) with a 7900HT Fast Real-Time PCR System (Applied Biosystems, Inc., Massachusetts, United States). The qPCR conditions were as follows: 95°C for 5 minutes, then 94°C for 30 seconds, 60°C for 30 seconds, and 72°C for 1 minute for 40 cycles. And the primers of SOCS1 were as follows: forward: 5′-CAGTCTCCACAGCAGCAGAG-3′, reverse: 5′-GAACGGAATGTGCGGAAGTG-3′. The primers of β-actin were as follows: forward: 5′-TCGTGCGTGACATTAAGGAGAA-3′, reverse: 5′-AGGAAGGAAGGCTGGAAGAGT-3′.
Western Blot
Total protein was extracted with RIPA Lysis Buffer (Sigma, St.Louis, Missouri, United States) and the concentration was determined by Bicinchoninic Acid Kit for Protein Determination (Sigma, St.Louis, Missouri, United States). Subsequently, sodium dodecyl sulphate–polyacrylamide gel electrophoresis was performed, and the protein was transferred to polyvinylidene fluoride membrane (Millipore, Darmstadt, Germany), which was further incubated with primary as well as secondary antibodies, and the antibodies applied in Western blot were listed in Table 1. Then, the bands were visualized by enhanced chemiluminescence substrate: Pierce ECL Plus Western Blotting Substrate (Thermo Fisher Scientific, Massachusetts, United States) and X-ray film (Kodak, New York, United States). And the gray value of Western bolt bands was detected by Image J Software (NIH, Maryland, United States). The SOCS1 protein relative expression was calculated by dividing the gray value of SOCS1 protein by the gray value of β-actin. And the pJAK2 and pSTAT3 protein relative expressions were calculated by dividing the gray value of pJAK2 and pSTAT3 protein by the gray value of JAK2 and STAT3, respectively.
Antibodies Applied in Western Blot.

Abbreviations: β-actin, beta-actin; HRP, horseradish peroxidase; IgG, immunoglobulin G; JAK2, Janus kinase 2; SOCS1, suppressor of cytokine signaling 1; STAT3, signal transducer and activator of transcription 3.
Cell Counting Kit-8 Assay
After the cells were washed by phosphate-buffered saline, Cell Counting Kit-8 (Dojindo, Kumamoto, Japan) as well as serum-free medium were added, and the cells were incubated for 2 hours. Then, the OD values were detected.
Statistical Analysis
Data analysis was performed using SPSS 22.0 (IBM, New York, United States) and GraphPad Prism 7.01 (GraphPad Prism, Inc., San Diego, United States). Data were displayed as mean and standard deviation, and the comparisons among groups were determined by the one-way analysis of variance or unpaired t test. The IC50 was calculated by the probit regression model. P value <.05 was considered statistically significant.*** represented P < .001. ** represented P = .01. * represented P = .05. NS represented no significance.
Results
Effect of As4S4 on Inhibiting Cell Viability in U266 Cells
To explore the effect of As4S4 on inhibiting cell viability, U266 cells were treated with various concentrations of As4S4 (0, 5, 10, and 40 μM) for 12, 24, and 48 hours. Cell Counting Kit-8 assay showed that cell viability all reduced under the treatment of As4S4, and the cell viability decreased along with the increased As4S4 concentration and increased treatment time (Figure 1A). Besides, IC50 value of As4S4 was reduced along with the increased treatment time (Figure 1B). These data indicated that As4S4 was able to inhibit cell viability in a dose-/time-dependent manner in MM cells. Besides, the IC50 value (which represented the concentration inhibiting 50% of in vitro cell growth) was about 10 μM, thus we chose this concentration for the following assays.

Tetra-arsenic tetra-sulfide inhibited cell viability in U266 cells. Relative cell viability of U266 cells treated by As4S4 with different concentrations for 12, 24, and 48 hours (A). IC50 value of As4S4 in killing U266 cells (B). Comparison among groups was determined by one-way analysis of variance. As4S4 indicates tetra-arsenic tetra-sulfide; IC50, the concentration inhibiting 50% of in vitro cell growth; NS, no significance. P value < .05 was considered significant. ***P < .001; **P < .01; *P < .05.
Detection of SOCS1 Methylation in U266 Cells
Methylation-specific polymerase chain reaction assay showed that SOCS1 methylation was elevated in U266 cells compared to control cells (Figure 2A). Moreover, SOCS1 mRNA expression was decreased in U266 cells compared to control cells (P < .01; Figure 2B), and the protein expression of SOCS1 was also reduced in U266 cells compared to control cells (P < .01; Figure 2C and D). These data suggested that SOCS1 methylation was enhanced but SOCS1 expression was inhibited in MM cells.
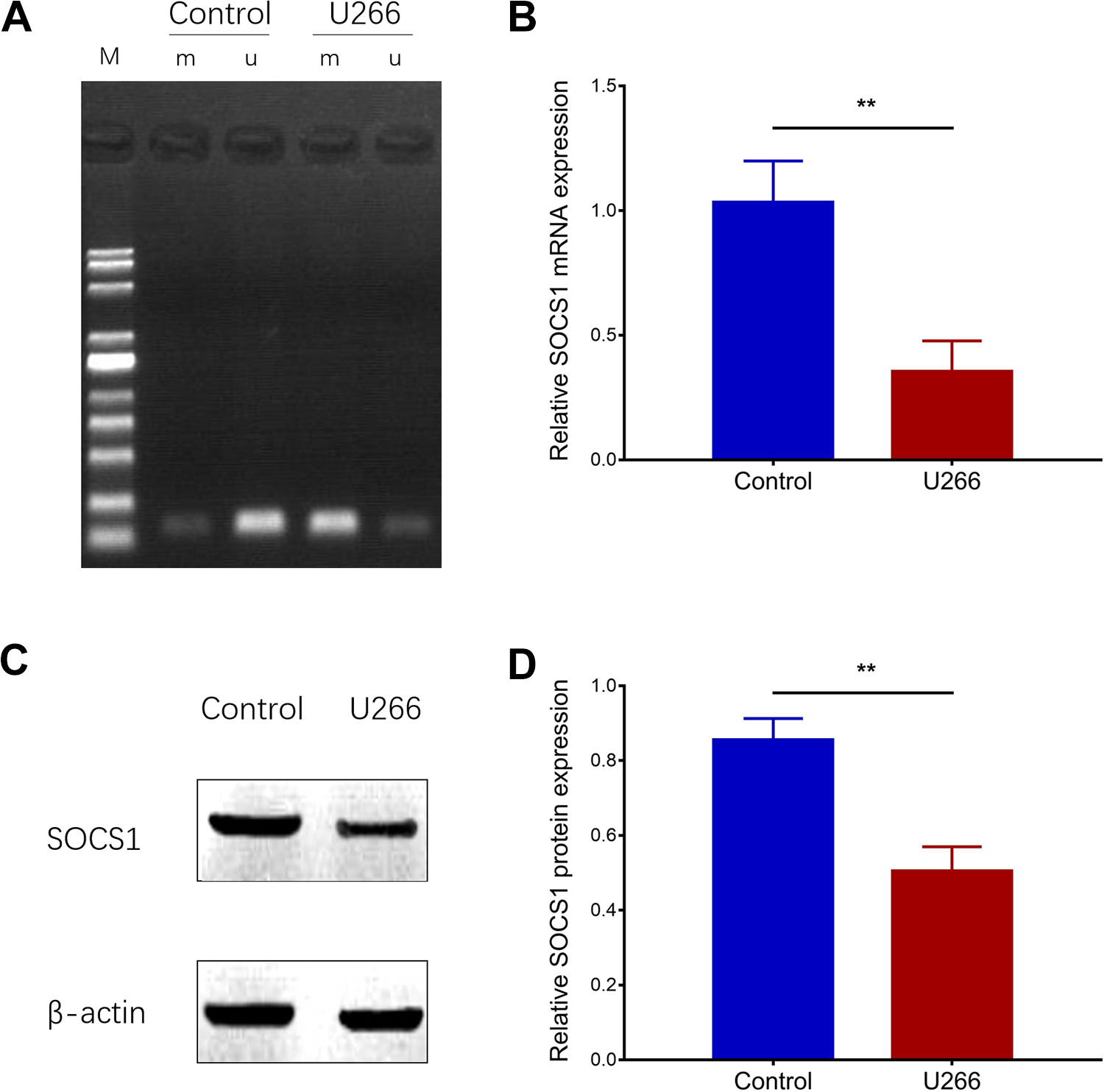
Comparison of SOCS1 methylation between U266 cells and control cells. Suppressor of cytokine signaling 1 methylation in U266 cells and control cells (A). Suppressor of cytokine signaling 1 mRNA expression in U266 cells and control cells (B). Suppressor of cytokine signaling 1 protein expression in U266 cells and controls (C and D). Comparison between groups was determined by unpaired t test. mRNA indicates messenger RNA; SOCS1, suppressor of cytokine signaling 1. P value < .05 was considered significant. **P < .01.
Effect of As4S4 on SOCS1 Methylation and Its Downstream JAK2/STAT3 Signaling Pathway in U266 Cells
To investigate the effect of As4S4 on SOCS1 methylation, U266 cells were incubated with 10 μM As4S4 for 12, 24, and 48 hours. Methylation-specific polymerase chain reaction assay showed that SOCS1 methylation was decreased at 12, 24, and 48 hours compared to 0 hour (Figure 3A), and it decreased along with the increased treatment time. Additionally, SOCS1 mRNA expression as well as protein expressions were increased at 12, 24, and 48 hours compared to 0 hour (all P value < .05), and they were increased along with the increased treatment time (Figure 3B-D). Moreover, protein expressions of pJAK2/JAK2 and pSTAT3/STAT3 were decreased at 12, 24, and 48 hours compared to 0 hour (all P value < .05), and they decreased along with the increased treatment time (Figure 3E and F). These data suggested that AS4S4 inhibited SOCS1 methylation and repressed the activation of JAK2/STAT3 signaling pathway in a time-dependent manner in MM cells.

Suppressor of cytokine signaling 1 methylation in U266 cells after treated by As4S4. Suppressor of cytokine signaling 1 methylation in U266 cells with As4S4 treatment (A). Suppressor of cytokine signaling 1 mRNA expression in U266 cells with As4S4 treatment (B). Suppressor of cytokine signaling 1 protein expression in U266 cells with As4S4 treatment (C and D). Protein expressions of pJAK2/JAK2 and pSTAT3/STAT3 in U266 cells with As4S4 treatment (E and F). Comparison between groups was determined by unpaired t test. As4S4 indicates tetra-arsenic tetra-sulfide; JAK2, Janus kinase 2; mRNA, messenger RNA; STAT3, signal transducer and activator of transcription 3. P value < .05 was considered significant. ***P < .001; **P < .01; *P < .05.
Effect of As4S4 on JAK2/STAT3 Signaling Pathway and Cell Viability in SOCS1 Knockout U266 Cells
After U266 cells were transfected with SOCS1 siRNA and control siRNA at 48 hours, SOCS1 mRNA expression (Figure 4A) as well as protein expressions (Figure 4B and C) were reduced in si-SOCS1 group compared to NC group (all P value < .05). Meanwhile, the protein expressions of pJAK2/JAK2 and pSTAT3/STAT3 were elevated in si-SOCS1 group compared to NC group (Figure 4D and E). Additionally, cell viability was higher in si-SOCS1 group compared to NC group (P < .01; Figure 4F). These results revealed that SOCS1 knockdown promoted the activation of JAK2/STAT3 signaling pathway and enhanced cell viability in U266 cells. Subsequently, when treated with 10 μM AS4S4, SOCS1 mRNA expression (Figure 5A) and protein expression (Figure 5B and C) were lower in si-SOCS1 + AS4S4 group compared to AS4S4 group (all P value < .05). Besides, the protein expressions of pJAK2/JAK2 and pSTAT3/STAT3 were increased in si-SOCS1 + AS4S4 group compared to AS4S4 group (all P value < .05; Figure 5D and E). Furthermore, cell viability was increased in si-SOCS1 + AS4S4 group compared to AS4S4 group (P < .01; Figure 5D). Taken together, our data suggested that AS4S4 inhibited SOCS1 methylation and reduced the activation of JAK2/STAT3 signaling pathway to repress cell viability in MM cells.

Suppressor of cytokine signaling 1 expression, JAK2/STAT3 pathway, and cell viability in SOCS1 knockout U266 cells. Suppressor of cytokine signaling 1 mRNA expression in si-SOCS1 group and NC group (A). Suppressor of cytokine signaling 1 protein expression in si-SOCS1 group and NC group (B and C). JAK2/STAT3 protein expressions in si-SOCS1 group and NC group (D and E). Cell viability in si-SOCS1 group and NC group (F). Comparison between groups was determined by unpaired t test. JAK2/STAT3 indicates Janus kinase 2/signal transducer and activator of transcription 3; mRNA, messenger RNA; NC, negative control; SOCS1, suppressor of cytokine signaling 1. P value < .05 was considered significant. ***P < .001; **P < .01.

Suppressor of cytokine signaling 1 expression, JAK2/STAT3 pathway, and cell viability in SOCS1 knockout U266 cells after As4S4 treatment. Suppressor of cytokine signaling 1 mRNA expression in As4S4 group and si-SOCS1 + As4S4 group (A); SOCS1 protein expression in As4S4 group and si-SOCS1 + As4S4 group (B and C). JAK2/STAT3 protein expressions in As4S4 group and si-SOCS1 + As4S4 group (D and E). Cell viability in As4S4 group and si-SOCS1 + As4S4 group (F). Comparison between groups was determined by unpaired t test. As4S4 indicates tetra-arsenic tetra-sulfide; JAK2/STAT3, Janus kinase 2/signal transducer and activator of transcription 3; mRNA, messenger RNA; SOCS1, suppressor of cytokine signaling 1. P value < .05 was considered significant. ***P < .001; **P < .01.
Discussion
Tetra-arsenic tetra-sulfide is a mineral and widely used drug in traditional Chinese medicine which potently inhibits the expression of a panel of oncogenes and also exerts a synergistic effect with various anticancer agents. 12 These characteristics make As4S4 a hotspot in the research of the cancer field recently. 12 -14 For instance, previous studies have shown that As4S4 inhibits gastric cancer cell proliferation via NFATc3/c-Myc pathway. Also, it inhibits lung carcinoma cell proliferation in a concentration-/time-dependent manner and induces cell apoptosis by activating caspase-3 and caspase-9. 19,20 Furthermore, a study shows that As4S4 reduces cell proliferation through scavenging intracellular ROS and downregulating hypoxia-inducible factor-1α in breast cancer cells. 21 These studies reveal that As4S4 may serve as a promising option in the treatment of various solid tumors.
Although the mechanisms of As4S4 in solid tumors have been frequently explored, the current evidence about the mechanism of As4S4 in hematological malignancies is mainly restricted to APL and CML. 22 -24 For example, a previous study shows that As4S4 inhibits cell proliferation and induces cell apoptosis via blocking the cell cycle in the S and G2/M phases in APL cells. 22 Besides, studies show that As4S4 downregulates SET protein expression and further promotes the proapoptotic factor protein phosphatase 2A (PP2A) expression to enhance cell apoptosis and represses cell proliferation in a dose-/time-dependent manner in APL cells. 11,23 Moreover, As4S4 facilitates CML cell apoptosis through downregulating miR-181 as well as Bcl-2 expressions and promoting apoptotic protein Caspase-3 activity. 24 These studies emphasized the antiproliferative or proapoptotic function of As4S4 in APL as well as CML, and we speculated that As4S4 might also have good antitumor effect in other hematological malignancies, especially MM, while the effect of As4S4 on MM cells was rarely reported. Thus, to investigate whether As4S4 had killing effect on MM cells, we treated U266 cells with different concentrations of As4S4, and we found that As4S4 repressed cell viability in a dose-/time-dependent manner in U266 cells, which may be due to (1) As4S4 might induce MM cell apoptosis via promoting proapoptotic factor (such as PP2A) or inhibiting antiapoptotic factor (such as Bcl-2), thereby reduced MM cell viability; 11,24 (2) As4S4 might regulate some downstream signaling pathways to decrease the cell viability, such as its downstream JAK2/STAT3 signaling pathway, which was displayed in our results. These data indicated that As4S4 possessed good antitumor effect via inhibiting cell proliferation in MM.
Suppressor of cytokine signaling family are reported to be the main physiologic and pathologic regulator in homeostasis, and it contains intracellular cytokine-inducible proteins SOCS1-7. 25 Among them, SOCS1 is an important member, which is a single-exon gene encoding 211 amino acids and has a CpG island. 26 Methylation of genes involves the addition of a methyl group to the CpG dinucleotide, which is observed in many cancers and is regarded to participate in the disease pathologies. 26 For SOCS1, its methylation has been reported to play a critical role in the pathology of some hematological malignancies. 15,27 For example, a study shows that SOCS1 methylation results in the promotion of cell proliferation in AML cells; moreover, another study displays that SOCS1 demethylation induces MM cell apoptosis. 15,27 Therefore, aberrant SOCS1 methylation may be an important cancerogenic factor in the pathology of various hematological malignancies. Moreover, we showed the antitumor effect of As4S4 in MM cells in this present study, and our preliminary experiment found that SOCS1 methylation could be regulated by As4S4. Taken together, we hypothesized that As4S4 might regulate SOCS1 methylation to exert its antitumor effect in MM, whereas the related evidence was limited. To address this problem, our study compared the SOCS1 methylation between U266 cells and controls, and we found that SOCS1 was hypermethylated in U266 cells compared to control cells. In addition, we treated U266 cells with As4S4, and we observed that AS4S4 inhibited SOCS1 methylation in a time-dependent manner in U266 cells, along with the reduced pJAK2/JAK and pSTAT3/STAT3 expressions, and these data might provide novel evidence for the potential application of As4S4 in treating MM.
According to the previous studies, SOCS1 methylation could activate JAK2/STAT3 signaling pathway, and the activation of JAK2/STAT3 signaling pathway stimulates cell proliferation, migration, and invasion in various hematological malignancies such as acute lymphoblastic leukemia, acute myeloid leukemia, and myelodysplastic syndrome. 28 -32 To investigate whether As4S4 inhibited SOCS1 methylation and enhanced theSOCS1 expression, thereby repressed the downstream JAK2/STAT3 signaling pathway and repressed MM cell viability, we conducted the knockdown of SOCS1 in U266 cells, and we found that pJAK2/JAK2 and pSTAT3/STAT3 expressions as well as cell viability were enhanced by SOCS1 knockdown. Furthermore, when treated with As4S4, pJAK2/JAK2, and pSTAT3/STAT3 expressions as well as cell viability remained increased in SOCS1 knockout cells compared to the cells without SOCS1 knockdown. Based on the results and indications we mentioned above, our data revealed that the antitumor effect of As4S4 on MM cells was through suppressing SOCS1 methylation as well as its downstream JAK2/STAT3 signaling pathway. These findings not only provided insight into the underlying mechanisms of As4S4 in MM but also suggested that inhibition on SOCS1 methylation might be a novel treatment approach for MM. Whereas, there were still some limitations in this study: (1) Deeper mechanisms were not investigated; (2) our experiments were conducted in only 1 cell line. Thus, a further study investigating deeper mechanisms and validating these findings in another cell line is needed.
In conclusion, As4S4 exhibits good killing effect on MM cells via repressing SOCS1 methylation and downstream JAK2/STAT3 signaling pathway, which may serve as a potential treatment option for MM.
Footnotes
Authors’ Note
Our study was approved by the Ethics Committee of The First Affiliated Hospital of Xi’an Jiaotong University (approval no. XJTU1AF2017LSK-138). All patients provided written informed consent prior to enrollment in the study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Shaanxi Provincial Key Research and Development Project of China [grant number 2018SF-033].