
Abstract
Human life history (LH) variation stems from developmental phenotypic responses to childhood environments. As with other psychological constructs, LH variation can be assessed by asking questions that are statistically linked to the underlying LH traits. This study introduces the fast-slow scale (FSS), a 35-item questionnaire designed to measure fast–slow LH strategies in adults. The FSS asks questions about trade-offs involving energy, resources, and time, as well as trade-off questions concerning the life domains of sexuality, sociality, and cognitive style. Two validation studies based on community samples totaling 1,491 participants support the internal structural and the external nomological validity of the FSS.
Keywords
Anything that varies in amount or kind can be measured (Thorndike, 1918). However, the use of a questionnaire to measure human life history (LH) strategies has sparked debate. The most fundamental question is whether LH theories, primarily derived from observations across different species, are applicable for investigating individual differences within human populations (e.g., Zietsch & Sidari, 2020 vs. Woodley et al., 2021). A subsequent point of contention revolves around the conceptualization of a unidimensional continuum representing fast-slow LH strategies and its adequacy in explaining differences among individuals or even across species (e.g., Bielby et al., 2007 vs. Del Giudice, 2020). This debate has reached no resolutions other than entrenching positions on both sides. Without trying to repeat and resolve the debate, we take the individual- and phenotypic-difference position and argue that human LH strategies can be measured based on clarifications of two key assumptions (Del Giudice, 2025).
First, although LH theory is primarily derived from genetically-based, between-species observations (Zietsch & Sidari, 2020), the theory can also explain within-species phenotypic variations that follow the same causal relations as those seen between species (Del Giudice, 2025; Galipaud & Kokko, 2020; Kuzawa & Bragg, 2012). LH variation consists of genetic differentiation at the population level, phenotypic plasticity at the individual level, and gene-by-environment interactions (Stearns, 1992). Most LH traits exhibit phenotypic plasticity in response to environmental factors during development (Caswell, 1983) and much of the LH variation represents individuals’ phenotypic responses to the local environments (Metcalfe & Monaghan, 2001; Stearns & Koella, 1986). LH models and measurements concern phenotypic more than genetic variation at the individual rather than population level (Acasuso-Rivero et al., 2019; Duffield et al., 2017; Ricklefs & Wikelski, 2002). As Ricklefs and Wikelski (2002, p. 462) explain, “the reaction norm, or phenotypic response to environmental variation, is usually included in the definition of life history.” Therefore, LH variation can be measured and is no different from that of other phenotypic traits, such as human intelligence or personality, that is routinely measured in psychology (Lu et al., 2021).
Second, the idea of an omnibus construct representing a fast-slow LH continuum may seem at odds with the biological research tradition of investigating individual LH traits (Stearns, 1992). Psychologists investigate the coordination of multiple LH traits or LH-related behaviors (Del Giudice, 2025), which together form an underlying strategy ranging from fast to slow (Del Giudice, 2020). Biologists have focused on specific traits individually, such as age at first reproduction, rather than a comprehensive LH strategy (Stearns, 1992). This apparent inconsistency between the biological and psychological approaches largely stems from biologists’ challenges of identifying coordinated traits that hold significance across species. However, this seeming difference does not mean that an underlying LH strategy does not exist or should not be measured. In fact, Stearns (1992) advocated for a multi-trait LH approach, stating, “the word ‘strategy’ simply means ‘complex adaptation’; it refers to the coordinated evolution of all the life history traits together… It was first necessary to understand the evolution of one trait at a time… This does not mean that life history strategies are unimportant or uninteresting. They have great potential importance” (Stearns, 1992, p. 492). A psychometric approach to measuring LH can unlock this potential and complements biometric research efforts. The Mini-K, developed by Figueredo et al. in 2006, and K-SF-42, published in 2017 (Figueredo et al., 2017), have been widely employed for this purpose, but both can be improved. Additional instruments are necessary not only to improve upon existing ones but also to provide necessary triangulation in construct validation as a unified effort within the discipline. The aim of the present study is to develop and validate such a new instrument.
The Psychological Model for Measuring LH Strategy
Measuring LH strategy as a cognitive or psychological construct involves assessing the processes by which resources and energy are allocated and traded off among different life activities, rather than focusing on the biological or demographic outcomes of such trade-offs (Chang & Lu, 2025). Specifically, a psychological measurement of LH strategies should not include such questions as the timing of first birth or number of offspring. These seemingly definitive LH-trait or life-table variables often misrepresent the underlying trade-off processes. A recent review, aimed at critiquing psychological LH research for being incongruent with biological conclusions, incidentally reveals instances where biological LH trait outcomes present misleading information (Dinh & Gangestad, 2024). For example, being monogamously married (rather than being promiscuous) in foraging societies is found to be correlated with increased offspring quantity (Dinh & Gangestad, 2024; Marlowe, 2001). This finding is deemed misleading because the input of monogamy should contribute to slow LH, whereas increased offspring quantity as the output represents fast LH. However, when we consider the underlying trade-off process or how individuals allocate energy and resources between mating and parenting, it becomes clear that, because of paternal provisioning, as well as added helpers at the nest, monogamously married women having more surviving children represents a slow but not fast LH outcome (Marlowe, 2001).
Thus, our psychological model and measurement of LH strategies aim to capture the LH trade-off process of how individuals allocate and prioritize energy, resources, and time to achieve fitness outcomes that may or may not fully associate with expected biological LH trait outcomes (Del Giudice, 2020; Figueredo et al., 2015). A slow LH strategy, which prioritizes physical and mental development over early reproduction, involves behaviors that channel resources and energy toward long-term growth rather than immediate reproductive outcomes (Figueredo et al., 2015). In contrast, a fast LH strategy, particularly in relation to sociality and time orientation, entails exerting effort and expending energy to secure immediate, zero-sum gains rather than investing in long-term cooperation and mutual benefits. Similarly, a slow LH strategy that emphasizes parenting over mating involves allocating more energy and resources to raising offspring than to seeking additional mates. In general, an LH measurement can tap into the processes by which energetic and material resources are preserved for, versus expended on, pursuing future-oriented activities, which include actions for, as well as a mindset of, accruing resources and delaying gratification, versus seeking immediate rewards and hedonic satisfaction. In all these instances, the adaptive trade-offs may or may not be captured by a single LH trait outcome such as timing of first birth or number of offspring but, with human animals, can aim to be captured by a questionnaire. This is the advantage of the psychometric approach over any biological methods.
By focusing on processes rather than outcomes, a psychological model offers a more nuanced understanding of how the environment, LH strategies, and behavior interact. As illustrated in Figure 1a, LH strategies function both as outputs shaped by early childhood experiences and as inputs that influence future development and behavior. In other words, early experiences or environments contribute to the formation of LH strategies, which can be assessed through questionnaires, while subsequent behaviors are shaped both by the LH strategies and by direct responses to childhood environments. In contrast, biological LH research typically conceptualizes LH traits as direct outcomes of environmental influences. Figure 1b illustrates this distinction by presenting the biological model, where early environments directly determine LH traits. Incidentally, the psychological model shown in Figure 1a has also been adopted in one area of biological research—namely, the pace-of-life syndrome (POLS) research. Like human LH research, POLS research examines within-species variation in personality syndromes and how environments shape the behavioral tendencies of fast and slow strategists (Réale et al., 2010).
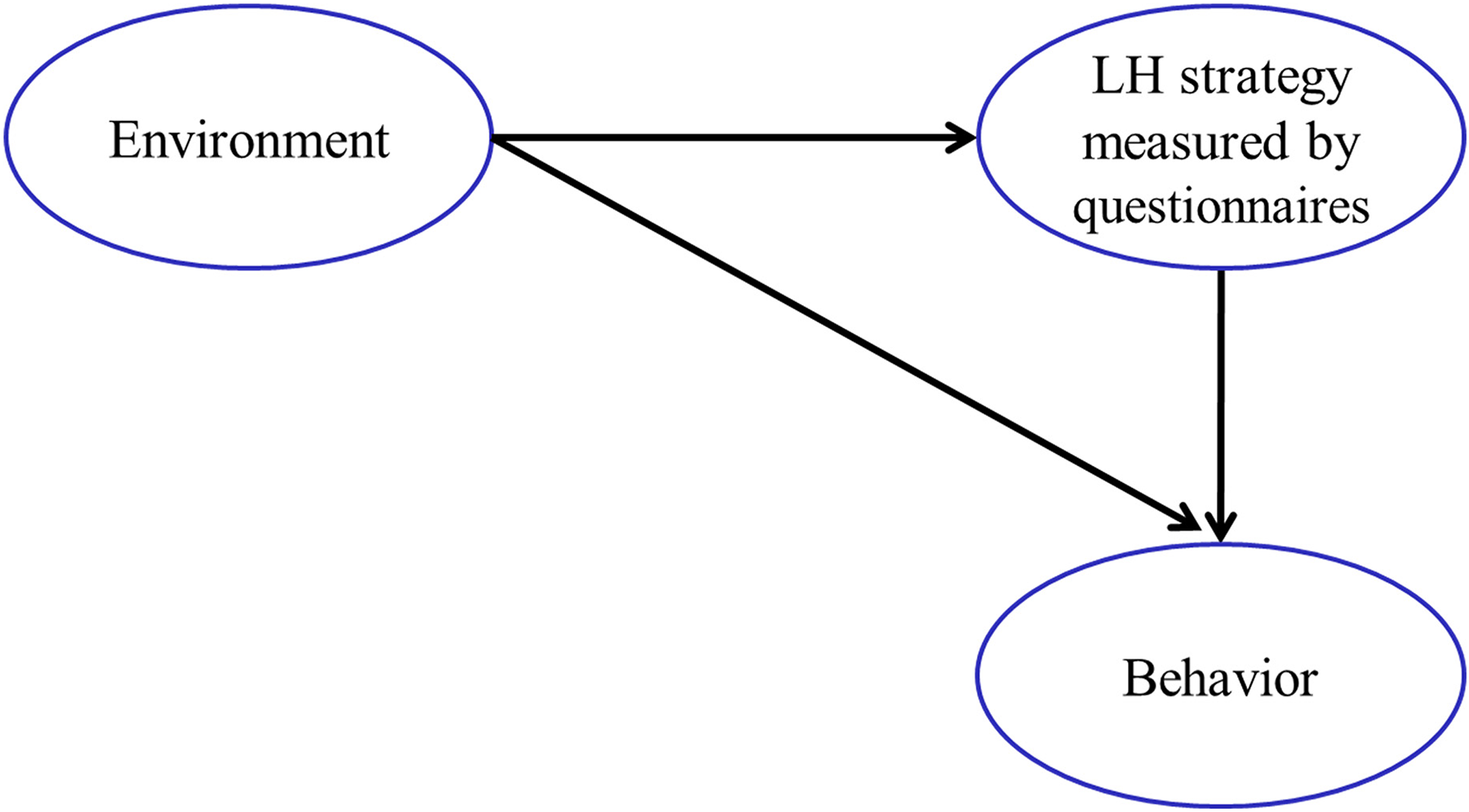
Process-Based Psychological LH Research: LH Strategy Assessed Possibly by a Questionnaire is Shaped by Early Environment and in Turn Influences Development and Behavior.
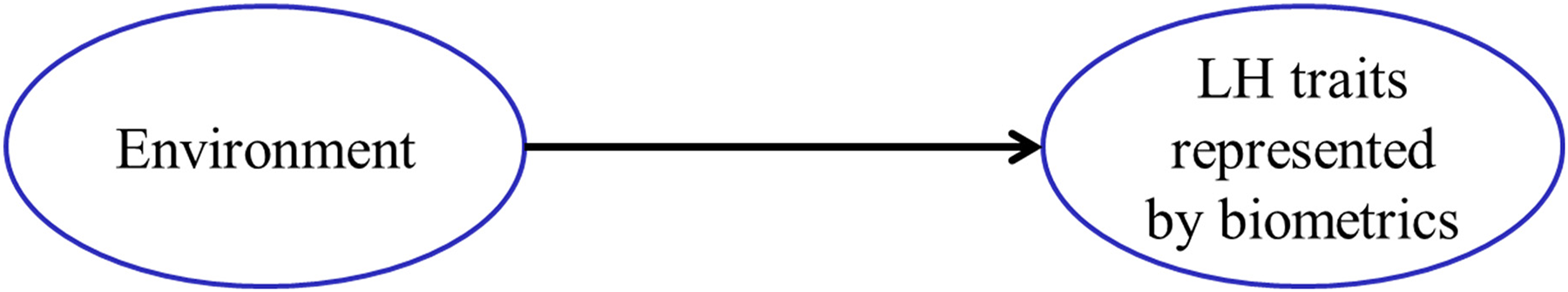
Outcome-based biological LH research: Early environment shapes LH traits as outcomes assessed by biodemographic indicators.
An examination of Figure 1a also highlights another potential limitation inherent in existing LH measurement and research. The limitation or improvement is that an LH strategy measure should not overlap with either environmental causes, on the one hand, or behavioral outcomes on the other hand. However, many existing instruments conflate these otherwise distinct elements, inadvertently measuring both ends of the relationship depicted in Figure 1a. For example, items in the Mini-K (Figueredo et al., 2006) and K-SF-42 (Figueredo et al., 2017) that assess relationships with parents, children, friends, and partners are indistinguishable from indicators of family or childhood environments. Other items in the Mini-K, as well as in instruments like the K-SF-42, assess specific behaviors and attitudes related to romantic relationships, family dynamics, agency, altruism, and religiosity. These items spill over into the behavioral outcome side of the model shown in Figure 1a. Such spill-over flaws partly result from the fact that both the Mini-K and K-SF-42 employ items borrowed from pre-existing instruments that were not originally designed within, or aligned with, an LH framework. In contrast, our measurement uses original items specifically developed to operationalize our conceptual model of LH strategies. We now turn to a discussion of specific operationalizations of LH strategies.
Energy, Resource, and Time
A recent development in human LH research is the reintroduction of bodily energetic conditions in defining LH strategies (Ellis et al., 2025; Volk, 2023); energetic conditions had a strong presence in the early literature (e.g., MacArthur & Wilson, 1967), in the form of resource availability, but were later neglected and are now being reinstated (Lu et al., 2025). Independent of this new development (Ellis et al., 2025), energetic conditions and the related availability of resources, as well as time, directly induce trade-offs between different LH priorities or strategies and, therefore, represent core components of an LH questionnaire. Specifically, internal energy, external resources, and their common denominator—time—are limited or constrained in their acquisition by animals (Ellis et al., 2009; Stearns, 1989). This limitation drives LH trade-off strategies (Gadgil & Bossert, 1970). Among the trade-offs between growth, maintenance (e.g., immune function, oxidative processes), and reproduction (Gadgil & Bossert, 1970), growth is age-dependent and typically loses relevance after maturity. For adults, for whom the LH measurement under discussion is intended, the primary LH trade-off is between maintaining the body, as well as the mind, and reproduction (Stearns, 1989)—or more specifically, mating, as the other component of reproduction, parenting, represents the growth and maintenance of the next generation. Thus, the essence of a questionnaire measuring LH strategies should focus on how human adults allocate energy, resources, and time toward maintaining and conserving the body and mind versus engaging in behaviors aimed at expending the body and mind ultimately to reproduce. Most animals make an effort to conserve energy, resources, and time rather than squandering them excessively (Stearns, 1989; Vander Wall, 1990; Wegmann et al., 2015). The balance between conservation and expenditure in relation to mating corresponds to slower versus faster LH strategies (Pontzer & McGrosky, 2022; van Noordwijk & de Jong, 1986).
The phenotype of faster versus slower LH is shaped by the energy expended on the underlying task (e.g., reproduction vs. maintenance or self-maintenance; Pontzer & McGrosky, 2022). Faced with similar environmental challenges, a slower LH strategy shifts energy investment toward maintenance and longevity, while a faster strategy prioritizes current reproduction at the expense of maintenance and future reproduction (Duffield et al., 2017). Numerous studies on various animals show that, when facing temporary physical setbacks, animals adopt a slower LH strategy by conserving energy and focusing on self maintenance rather than reproduction (e.g., Billman & Belk, 2014; Hudson, 2001; Mougeot et al., 2005). However, when physical conditions are terminally compromised, animals switch to a faster LH strategy, forgoing maintenance and expending energy on reproduction (e.g., Hendry et al., 2016; Jones et al., 2008; Marzal et al., 2008). Although these studies primarily focus on the severity status of physical challenges, the underlying principle is consistent: both between and within individuals, faster and slower LH strategies are shaped by trade-offs between energy allocation toward maintenance and that toward reproduction (Elliott et al., 2014). In human populations, this trade-off can be measured by examining how much individuals prioritize health and longevity versus neglecting the body and mind.
The same principles apply to the other two factors, resources, or material resources, and time, or temporal resource, both of which contribute to the conversion of bodily energy. In addition to the energy conversion dynamics, time adds a separate dimension, directly related to current versus future reproduction, which influences faster versus slower LH strategies. In both animal and human literature, resource acquisition (such as food-seeking) and financial decisions are directly tied to this time dimension. Animals often consume resources immediately after acquiring them, manifesting a fast LH strategy. They may also exhibit a slow LH strategy by saving and hoarding resources for future events like wintering, nesting, or migration, particularly when resources are scarce or competition is intense (Koivisto et al., 2023; Vander Wall, 1990). These conditions define saturated versus unsaturated environments, which independently drive slower versus faster LH strategies (Pianka, 1970). It is also well established in human populations that impulsive versus future-oriented financial decisions align with faster versus slower LH strategies (Gineikiene et al., 2022; Mittal & Griskevicius, 2014). Thus, resources and time, alongside energy expenditure, are key elements in measuring fast-slow LH strategies.
Sexuality, Sociality, and Cognitive Style
Sampling representativeness and relevance are more important than sheer size. In part to reduce overlap with either environmental causes or behavioral outcomes depicted in Figure 1a, the decision on what other components to include in our LH measurement is motivated by trying to limit rather than expanding the potential choices. Existing instruments such as the 199-item Arizona Life History Battery (Figueredo et al., 2006) and the 7-factor K-SF-42 (Figueredo et al., 2017) may have sacrificed the more important relevance for the less important comprehensiveness. Our final selections are narrowed to three broad areas: sexuality, sociality, and foraging or cognitive style, the last of which is notable because it has not been included in existing LH questionnaires. Sexuality and mating-related indicators should represent a significant part of a LH measurement, as they reflect the fundamental trade-off between maintenance and reproduction or, more specifically, mating. It is worth reiterating that these indicators should not replicate demographic LH traits or life table variables commonly used in biological LH research. Here, modern living also introduces additional confounding factors, such as contraception, abortion, cultural and religious influences, and individual circumstances like the availability of space for copulation and child rearing. These factors can significantly affect life table questions, making them misrepresent true LH trade-off priorities. For the sexuality domain, it is much simpler and more informative to employ sociosexual orientation (e.g., casual sex vs. committed relationships), which has already been shown to differentiate between fast and slow LH strategies (Fernandes et al., 2016).
Sociality and foraging or cognitive styles represent evolutionary domains that are both species-general, underlying LH development, and uniquely human, contributing to the slowing of human LH (Chang & Lu, 2025). The enlargement of the human brain is a prominent slow LH trait, linked to other slow LH developments such as premature birth (a paradoxical slow-LH adaptation associated with bipedalism and larger cranial size), an extended childhood (unparalleled in the animal kingdom), postnatal brain development that accounts for three-fourths of adult brain size, biparenting and monogamy, and, representing reciprocal causality, large social groups and complex sociality. Centering on social and cognitive development, these interconnected developmental events have contributed to a tenfold slowing of human LH over the past 2 million years (Smith & Tompkins, 1995). POLS research also shows that sociality is similarly linked to LH development across other species. In high-predation environments, animals exhibit bold, antagonistic, and aggressive behaviors toward both conspecifics and heterospecifics (Réale et al., 2000; Wolf et al., 2007). Such boldness and aggression are associated with fast LH traits, including faster growth and earlier reproduction, with stronger correlations in populations facing greater predation pressure (Biro & Stamps, 2008). By contrast, animals living in riparian habitats or facing low predatory risks show less aggressive, more inclusive, and more affiliative sociality (Riechert & Hall, 2000; Rosenblum & Andrews, 1994).
POLS research also shows that sociality in turn is correlated with foraging and the evolved cognitive style. Foraging is characterized by thorough and exploitative versus superficial and exploratory search style, which is also represented as deliberate versus intuitive cognitive style in humans (Hills, 2006; Hills et al., 2015; Wang et al., 2022, 2023). Superficial exploration is linked to boldness and aggression, reflecting fast LH sociality, while thorough exploration is associated with more socially affiliative behaviors and slow LH strategies (Chang et al., 2019; Kurvers et al., 2010). Superficial exploration is common in high-predation environments, whereas thorough exploration is more typical of safer environments (Both et al., 2005). Exploratory search styles correspond to fast LH traits such as high growth rate, early reproduction, and low parenting investment, whereas the opposite is true for exploitative searches (Biro & Stamps, 2008; Both et al., 2005). These two search styles have been redefined as fundamental trade-offs between exploratory or intuitive and exploitative or deliberate cognition, permeating all cognitive behaviors from foraging and visual search to memory and decision-making (Del Giudice & Crespi, 2018; Hills et al., 2015; Wang et al., 2022; Wang et al., 2023). Moreover, this cognitive trade-off is deeply rooted in adaptive cognitive phenotypes associated with fast and slow LH strategies, with fast strategists favoring exploration to adapt quickly to changing environments, and with slow strategists emphasizing exploitation for efficiency and stability in more constant surroundings (Wang et al., 2023).
Present Study
Within the aforementioned psychological model centered on trade-offs involving energy, resources, and time, and the LH calibration concerning the three domains of sexuality, sociality, and cognitive style, we wrote and re-wrote and translated and back-translated items, pilot-tested them, and, based on item analysis (e.g., item-total correlation) and other psychometric analyses (e.g., factor analysis) conducted on multiple pilot samples, finalized a set of 35 items presented in Table 1 and collectively named the fast-slow scale (FSS). We used the Susan Harter format to present the FSS items. Participants would select one of a pair of statements (e.g., “Some people are more into serious relationships, while other people are more into casual sex”) that best described the participants and would then rate it on a 3-point scale ranging from 1 (a little like me) to 3 (a lot like me). Half of the items featured the slow LH statement first, followed by the fast LH statement (the example above), while the other half featured the fast LH statement first, followed by the slow LH statement (e.g., “Some people have many sexual partners, while other people have a few serious relationships”). We took care to balance the connotation and emotional tone of each pair of fast and slow LH statements. Since slow LH statements tend to appear more socially desirable than fast LH statements, we paid particular attention to wording to ensure both were equally appealing. For analysis, half of the items would be reverse-coded—either in the fast or slow LH direction—so that all items were aligned consistently. We then conducted two studies to validate the FSS, assessing both its internal construct structure and its external nomological relationships. Based on community samples totaling 1,491 Chinese participants, the results provided adequate evidence supporting both the internal construct and external nomological validity. Below, we present these two studies and report their findings.
The 35 Items of the FSS.

Study 1
Sample
A community sample of 701 young adults (314 males; Mage = 25.38, SD = 6.42) was taken from a city in northeastern China. Among them, 437 or 62% had university education, 264 (38%) had high school education. One hundred and 48 (21%) were married, 256 (37%) were unmarried but were then in a relationship, and the rest (42%) reported not being in a relationship. The study procedures and questionnaire content were approved by the Institutional Review Board of the authors’ universities. Participants provided written informed consent.
Measures
Results and Discussion
We first conducted a two-level confirmatory factor analysis (CFA) on the 35 items to test the internal structure of the FSS. At the first level, the 35 items were defined to load onto their respective factors: energy, resource, time, sexuality, sociality, and cognitive style. The factor loadings ranged from 0.36 to 0.83, with an average of 0.65, and were all statistically significant. Although moderate, these values are consistent with those of existing LH measures reported in the literature (e.g., Figueredo et al., 2006) and are considered adequate. At the second level, the six primary factors were linked to a single secondary factor, named the fast-slow LH factor. The factor loadings (energy: 0.84; resource: 0.82; time: 0.96; sexuality: 0.89; sociality: 0.84; cognitive style: 0.75) were adequate. The internal factor structure fit the data well, with satisfactory goodness of fit statistics (CFI = 0.93, TLI = 0.91, RMSEA = 0.047, SRMR = 0.061; χ²/df = 2.53). Overall, the CFA results support the internal structure of the FSS, with adequate factor loadings and goodness of fit statistics.
Next, we computed the correlation between the FSS, which was coded in the direction of slow LH, and Mini-K to establish concurrent validity. The correlation was 0.52, indicating adequate validity.
We also examined the demographics with respect to the FSS. There was no significant gender difference (t = .73, p = .47) with men (M = 3.70, SD = .75) scoring slightly lower than women (M = 3.75, SD = .76) on the FSS coded in the direction of slow LH. There was a significant difference between the two levels of education (t = 13.46, p < .001), with university graduates scoring higher (M = 4.00, SD = .70) than high school graduates (M = 3.27, SD = .61). There were significant differences between the three relationship status groups (F = 55.76, p < .001) with married people scoring the highest (M = 4.24, SD = .73) and with those not in a relationship scoring the lowest (M = 3.52, SD = .79). There was no significant interaction effect involving any of the demographic variables.
Finally, we examined partial nomological or predictive validity by testing the structural relationships involving the FSS and two behavioral outcomes: present orientation and impulsiveness. In this analysis, the FSS was coded in the direction of fast LH. The results are reported in Figure 2, and the zero-order correlations are shown in Table 2. As shown in Table 2, the correlations are adequate and in the expected directions, warranting the testing of the model in Figure 2. Model testing results showed an adequate fit of the model to the data (χ²/df = 3.16, CFI = 0.98, TLI = 0.97, RMSEA = 0.056, SRMR = 0.064). Factor loadings and structural coefficients were all statistically significant, in the expected directions, and represent meaningful magnitude. Specifically, the FSS significantly predicted the two behavioral outcomes (β = 0.46 with present orientation and β = 0.28 with impulsiveness) in the expected directions. Overall, these results support the concurrent, predictive, and partial nomological validity of the FSS.

LH Strategy Measured by the FSS in Relation to Present Orientation and Impulsiveness in Study 1. Note. *** p < .001.
Means, Standard Deviations, and Correlations of Variables Used in Study 1.

Note. All correlations are significant with p < .001.
Study 2
Study 1 partially tested the nomological validity, representing only half of the full model shown in Figure 1a. To address this limitation, we tested the complete model presented in Figure 1a in Study 2. Additionally, Study 2 provides an opportunity to cross-validate the internal factor structure of the FSS.
Method
Sample
A convenience sample was taken from a trade school in a northern city of China. The sample consisted of 790 youths (366 males; Mage = 17.51, SD = 0.73) and their parents (fathers’ Mage = 47.84, SD = 4.59; mothers’ Mage = 47.14, SD = 4.65). The study procedures and questionnaire content were approved by the Institutional Review Board of the authors’ universities. Participants provided written informed consent.
Measures
The same three measures described in Study 1, FSS, Mini-K, and Impulsiveness, were given to the youth participants. Internal consistency reliability estimates were α = 0.87 for FSS, α = 0.90 for Mini-K, and, for the three Impulsiveness subscales, α = 0.83 for Attentional Impulsiveness, α = 0.89 for Motor Impulsiveness, and α = 0.91 for Nonplanning Impulsiveness. In addition, the following measures were given to the youths or their parents.
Results and Discussion
We first validated the FSS by conducting a two-level CFA on the 35 items. The primary factor loadings ranged from 0.44 to 0.67, with an overall average of 0.63. At the second level, the six primary factors are loaded onto a single fast-slow LH factor. The factor loadings (energy: 0.66; resource: 0.93; time: 0.64; sexuality: 0.43; sociality: 0.48; cognitive style: 0.65) are considered adequate, although the CFA results are overall weaker than those obtained in Study 1, possibly because the FSS, designed for adults, may not align as closely with the present sample of youths. The sexuality factor and items had the lowest loadings, likely because most participants in this sample had not yet had sexual experiences. Despite this potential limitation, the internal factor structure fit the data well, with goodness of fit statistics comparable to those of Study 1 (CFI = 0.90, TLI = 0.92, RMSEA = 0.042, SRMR = 0.056; χ²/df = 2.33).
We then examined the nomological relationships involving the FSS by operationalizing the full model of psychological LH research depicted in Figure 1a. The substantiated model and results are presented in Figure 3, and zero-order correlations and other descriptive statistics of all the variables collected in this study are reported in Table 3. In these analyses, the FSS was coded in the direction of fast LH. Model testing results showed an adequate fit to the data (CFI = 0.96, TLI = 0.94, RMSEA = 0.055, SRMR = 0.058; χ²/df = 3.42). Parameter estimates were all statistically significant and in the expected directions. Specifically, environmental harshness and unpredictability positively predicted the FSS. The FSS, in turn, significantly predicted two behavioral outcomes, aggression and impulsiveness, in the expected directions. Finally, we correlated the FSS, coded in the direction of slow LH, with the Mini-K to achieve a satisfactory concurrent validity estimate (r = 0.48). Overall, these results support the nomological and construct validity of the FSS.

LH strategy measured by the FSS is shaped by early environments and influences behavioral outcomes in Study 2. Note. * p < .05, *** p < .001.
Means, Standard Deviations, and Correlations of Variables Used in Study 2.

Note. Most correlations are significant with p < .01.
General Discussion
Overall, the results from the two studies support the structural and nomological validity of the FSS. Structurally, both studies consistently identified the internal structure of the FSS. The primary factor loadings averaged above 0.60, while the goodness-of-fit statistics were mostly adequate. It is noted though that fitting a larger number of factors could improve some of the goodness-of-fit statistics as is often the case. Importantly, all primary factors are loaded onto a single secondary factor representing the unidimensional fast-slow LH strategy continuum. As expected, all secondary factor coefficients were statistically significant, directionally consistent, and of adequate magnitude. While nomological or construct validity requires continuous validation, we obtained sufficient initial evidence.
These findings regarding the internal structure and external relations of the FSS are grounded in a psychological LH theoretical framework. Unlike biological models of LH, our psychological model emphasizes the processes by which behaviors are executed to balance the allocation of energy, resources, and time between adaptive needs, forming fast versus slow LH strategies. A slower LH strategy is associated with a longer time horizon, conserving energy and resources for future reproductive and fitness prospects. Conversely, a faster LH strategy expends energy and resources, favoring present reproduction and short-term fitness gains. The allocation of energy, resources, and time between maintenance (the conserving mode representing slow LH) and reproduction (the expending mode representing fast LH) represents a species-general principle adopted by all animals. In response to external environments and internal states, animals make trade-off decisions between conserving energy (e.g., resting) or conserving resources (e.g., hoarding) and investing in reproduction (Larsen et al., 1997; Vander Wall & Jenkins, 2003). Particularly when physically challenged (e.g., by disease or injury), animals may adopt a conserving mode (slow LH), reallocating resources and energy towards self-maintenance and away from reproduction. Alternatively, they may switch to an expending mode (fast LH), increasing reproductive efforts at the cost of decreased somatic maintenance and future reproduction (Clutton-Brock, 1984; Williams, 1966). These are plastic phenotypes (i.e., strategies), not fixed traits or canalized genotypes (e.g., iteroparity versus semelparity). This scientific insight further supports the validity of our psychometric and individual-difference approach. For both humans and other animals, these two sets of plastic phenotypes or strategies represent and measure individual differences in the extent to which one position towards a slower versus a faster LH.
Another feature of our psychological model, from which the FSS is derived, is that LH strategy is conceptualized to mediate the relationship between early environmental inputs and later developmental outcomes, as depicted in Figure 1a. The LH strategy is shaped by the environment and, in turn, influences development and behavior. Importantly, an LH strategy questionnaire should not include or measure elements from either environmental inputs or behavioral and developmental outputs. Of these two potential overlaps, avoiding overlap with the behavioral outcome side is more challenging. Some of the items from existing LH measures (e.g., Mini-K and K-SF-42) are almost indistinguishable from measures of behavioral outcomes. To mitigate this potential validity issue, we limit our sampling of life activities to three facets: sexuality, sociality, and foraging or cognitive style. These facets represent broad categories rather than specific behaviors, and they are evolutionarily ancient, species-general, inter-correlated, and predictive of other LH traits and variables (Biro & Stamps, 2008; Réale et al., 2000; Wolf et al., 2007), suggesting that they collectively represent broad LH strategies rather than specific behavioral outcomes.
The POLS literature demonstrates that non-human animals may consistently differ in sociality, such as boldness and aggressiveness, and in foraging style, such as fast, superficial, and novelty-seeking searches. These personality syndromes and cognitive styles are associated with early reproduction and reduced parental investment (Biro & Stamps, 2008; Réale et al., 2000; Wolf et al., 2007), suggesting a broad fast (vs. slow) LH strategy. Separately, the fact that POLS studies focus on within-species phenotypic variations supports using a similar approach in human LH research, such as employing a questionnaire to study LH-related individual differences. Finally, the facets of exponential brain and cognitive development, large social groups and complex sociality, as well as bi-parenting, monogamy, and sexual exclusivity represent key components of human LH development (Lu et al., 2022; 2023) and should be included in a questionnaire to measure LH strategies in the human population, as we did with the FSS.
There are limitations to the FSS and the present study. First, the development of the FSS is based on Chinese samples. Future research should validate the FSS across other cultural and ethnic groups to establish ecological validity. Second, part of the validation of the FSS is based on retrospective data rather than the longitudinal data demanded by LH theory. Future validations of the FSS should employ longitudinal designs to examine the relationship between childhood environment and FSS scores in adulthood. Lastly, the present validation study yields moderate parameter estimates, such as factor loadings and structural coefficients, which might cast doubt on construct validity. However, this potential weakness reflects the inherent difficulty of using a psychometric approach to measure LH. The psychometric approach cannot fully capture LH but primarily addresses cognitive or conscious elements (Figueredo et al., 2006), resulting in moderate estimates. Despite this inherent limitation, our estimates are consistent with or slightly stronger than those reported in the literature. For example, validations of the Mini-K show factor loadings ranging from .24 to .63, averaging .44 and .63, with validity estimates ranging from .13 to .53 (Figueredo et al., 2007; Richardson et al., 2017). Similarly, factor loadings ranged from .35 to .62 and validity estimates from −.02 to .16 in another validation study involving the High K Strategy Scale (HKSS: Giosan, 2006) (Copping et al., 2014). Despite these limitations, the theory-based FSS provides a reasonably valid and reliable assessment tool for investigating phenotypic variation in human LH strategies.
Footnotes
Ethical Considerations
This study was approved by the Institutional Review Board of the authors’ universities.
Consent to Participate
Informed consent was obtained from all individual participants included in the study.
Funding
The research was supported by the Hong Kong Research Grants Council Humanities and Social Sciences Prestigious Fellowship Scheme (35000824) and Internal Grants (P0042481 and P0048500) of Department of Applied Social Sciences, The Hong Kong Polytechnic University.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.