
Abstract
In his groundbreaking research, Geoffrey Miller (1999) suggests that artistic and creative displays are male-predominant behaviors and can be considered to be the result of an evolutionary advantage. The outcomes of several surveys conducted on jazz and rock musicians, contemporary painters, English writers (Miller, 1999), and scientists (Kanazawa, 2000) seem to be consistent with the Millerian hypothesis, showing a predominance of men carrying out these activities, with an output peak corresponding to the most fertile male period and a progressive decline in late maturity. One way to evaluate the sex-related hypothesis of artistic and cultural displays, considered as sexual indicators of male fitness, is to focus on sexually dimorphic traits. One of them, within our species, is the 2nd to 4th digit length (2D:4D), which is a marker for prenatal testosterone levels. This study combines the Millerian theories on sexual dimorphism in cultural displays with the digit ratio, using it as an indicator of androgen exposure in utero. If androgenic levels are positively correlated with artistic exhibition, both female and male artists should show low 2D:4D ratios. In this experiment we tested the association between 2D:4D and artistic ability by comparing the digit ratios of 50 artists (25 men and 25 women) to the digit ratios of 50 non-artists (25 men and 25 women). Both male and female artists had significantly lower 2D:4D ratios (indicating high testosterone) than male and female controls. These results support the hypothesis that art may represent a sexually selected, typically masculine behavior that advertises the carrier's good genes within a courtship context.
Introduction
Evolutionists place cultural displays into the broader framework of sexual selection by considering the evolutionary advantage of such behaviors. Artistic ability, together with other human and nonhuman behaviors, has no apparent survival value. However, Darwin (1871) himself pointed out that this holds true for traits associated with male courtship, within a framework of sexual selection. The biological relevance of such behaviors has much to do with mating success and little with adaptation to the environment. Thus, the propagation of these traits is thought to result from the action of sexual selection rather than natural selection (Darwin, 1871). Moreover, according to the Millerian courtship model (Miller, 1999), cultural displays may be considered evolutionary products enhancing the chances of individual reproductive success. From this perspective, females seem to be more selective in choosing their partners than males who, instead, tend to exhibit their environmental fitness-adapted skills. In the resulting sexually dimorphic picture, males clearly tend to prefer physical attributes, while females look rather for social status and resources of their potential partners (Buss, 1989). The outcomes of several surveys conducted on jazz and rock musicians, contemporary painters, English writers (Miller, 1999), and scientists (Kanazawa, 2000) seem to be consistent with the Millerian hypothesis, showing a predominance of men carrying out these activities, with an output peak corresponding to the most fertile male period (at around age 30) and a progressive decline in late maturity. Interestingly, a similar pattern has been observed among male human homicides (Daly and Wilson, 1984), confirming the hypothesis of a biological relationship between the mechanisms of sexual selection and the evolution of male violent competition in order to access resources and females. However, if such criminal acts appear to be attributable to the action of intrasexual selection, which refers to the competition among males for access to mates, the production of art, instead, seems to find a place in the hierarchical structures of intersexual selection; i.e., the female process of choosing a suitable mating partner.
Consequently, masculinity (i.e., a higher testosterone level) is positively correlated to cultural and artistic displays. However, the theory is only half complete since it does not provide any scientific evidence that can address socially- and culturally-dependent explanations. Analyzing sexually dimorphic traits is one way to uncover experimental evidence specifically related to this hypothesis. The sexually dimorphic brain structure claim (Baron-Cohen, 2003) suggests that prenatal exposure to different steroidal levels, such as testosterone, plays a central role in determining different behavioral outcomes. Specifically, whereas women are superior to men with regard to language tasks (Kimura, 1992), tests of social judgment (Halpern, 1992), measures of empathy and cooperation, and pretend play in childhood (Hutt, 1972), men are ahead of women with regard to mathematical reasoning (Mills, Ablard, and Stumpf, 1993) and spatial skills (mostly Euclidean geometric navigation; Linn and Petersen, 1985). In other words, female brains are more developed in terms of “folk psychology” and male brains are more developed in terms of “folk physics.” The former brain type is closely related to “mindreading” capacities and the latter to understanding physical objects. In this perspective, autism and Asperger syndrome (which consist of impairments in detecting the mental states of others, the production of spontaneous pretend play (Baron-Cohen, 1987) and mindreading abilities in general) appear to be extreme forms of the male brain type. In addition, there is a strong male bias in the sex ratio of autism, or AS (within the population of autism the male:female sex ratio is 4:1).
In addition to discriminately choosing sexual partners, many members of other species show a preference for traits that are products of sexual selection. In our species, women tend to view creativity as a very attractive, though not essential, feature (Li, Bailey, Kenrick, and Linsenmeier, 2002). According to the Millerian model, it is therefore possible to consider the performance of creativity as an almost exclusively male phenomenon. Some experiments conducted on samples of both sexes have demonstrated the existence of an exclusively male positive correlation between creativity and romantic priming (Griskevicius, Cialdini, and Kenrick, 2006). In other words, the same psychological input does not seem to affect creative abilities in women, regardless of the scenario proposed (short-term or long-term relationships), which seems to give further support to the hypothesis of a clear role distinction between sexes within a courtship context.
A sexually dimorphic anatomical trait, within the human species, is the ratio of the 2nd to 4th digit length (Baker, 1888; George, 1930). In men, the index finger is generally shorter than the ring finger (2D:4D < 1), which indicates a higher testosterone level, whereas in women the 2nd digit is longer (2D:4D > 1) (Manning et al., 1998). In fact, Manning, Scutt, Wilson, and Lewis-Jones (1998) showed that the 2D:4D ratio was sexually dimorphic in children 2 years of age and older, but they did not measure younger children; however, it is reasonable to assume that 2D:4D is sexually dimorphic from the early stages of intrauterine life, about the end of the first trimester, in correspondence with a different hormone exposure: A lower ratio between the second and fourth finger reflects embryonic exposure to high testosterone levels, and a higher ratio is the result of the exposure to a low testosterone prenatal environment (Breedlove, 2010).
Although this hypothesis was not supported by any experimental tests, the close anatomical similarity of digital proportions with mice has permitted the analysis of data concerning embryonic hormonal indices in the uterus during the development of digital cartilages. Zheng and Cohn (2011), in this experimental study, have provided scientific validation for the use of digit ratios as an index of the type of uterine hormonal environment. They have also shown that, in mice, different digital proportions develop during a narrow window of embryonic growth between the formation of digit condensations and the 17th embryonic day (E17), which do not change in the postnatal period. In addition, treatment of pregnant females with pharmaceutical antiandrogens between the 12th and 15th day led them to give birth to male offspring with higher/feminized 2D:4D ratios.
The levels of hormone exposure in utero seem to determine a wide range of physiological and psychological conditions. In particular, several studies conducted on samples of adults of both sexes, based on X-ray measurement of the 2D:4D ratio, have shown that prenatal hormone exposure is the basis for the development of different athletic abilities (Manning and Taylor, 2001), fertility (Manning et al., 2000), sex-biased diseases (Manning, Baron-Cohen, Wheelwright, and Sanders, 2001), social behaviors (Coates, Gurnell, and Rustichini, 2009), and sexual orientation (Williams et al., 2000). In one study (Sluming and Manning, 2000), prenatal testosterone was shown to be linked to spatial abilities and musicality, with the capacity to produce music even representing a sexually dimorphic trait pointing to higher male fertility. Although our paper addresses a different form of art (i.e., painting), it also presents several similarities with the work of Sluming and Manning (2000) as it investigates the possible correlation of low 2D:4D ratio and visual arts ability.
The objectives of our research were to combine the Millerian theories on sexual dimorphism in cultural displays and the digit ratio method, using it as an accurate indicator of testosterone exposure in utero. According to the Millerian courtship model, artistic exhibitions are a male-predominant behavior displayed to impress female counterparts who, in turn, based on the sexual role distinction between genders, are more attracted by talented partners who are unconsciously advertising their “good genes.” On the other hand, the digit ratio method could represent a possible way to address the alternative, socially- and culturally-dependent explanations of sex differences in social and sexual behaviors. Even if, statistically, the amount of male artists within our species is higher than the amount of female artists, this could be due to historical and cultural constraints. However, if androgenic levels are positively correlated with artistic exhibition, the sexual dimorphism observed among artists within our own species could receive a scientific and hormonal-based explanation. Thus, we predict that both female and male artists should exhibit low 2D:4D ratios compared to non-artists.
Materials and Methods
Our subjects were 50 visual artists (25 men and 25 women) randomly recruited from the Las Cruces's art community in New Mexico. Artists had different professional levels and some of them were involved, apart from visual art, in several artistic activities, such as pottery, sculpture, and photography. All artists were Caucasians, the majority originating from the New England area, and five were Europeans. Similarly, controls (25 men and 25 women) were North American and European Caucasians, randomly recruited both from adult social clubs in the area of Las Cruces (12 men and 15 women) and from Italy (13 men and 10 women). Recruitment was restricted to an age range of 18 to 80. No Hispanics were included in the study, as there are data indicating that Hispanics tend to have a low 2D:4D ratio compared to Caucasians (Loehlin, McFadden, Medland, and Martin, 2006). After being informed about the experimental procedures and providing consent, all participants were asked to fill in a questionnaire. They were asked to indicate their age, sex, and ethnicity and to rate themselves as visual artists on a scale from “non-artist” to “professional” (“non-artist,” “amateur,” “adequate,” “competent,” “accomplished,” and “professional”). Artists were then asked to indicate the approximate number of visual art works sold. All participants filled in the Bem Sex Role Inventory questionnaire (BSRI: Bem, 1974, 1977; Hyde and Phillis, 1979) to calculate the androgynous personality factor using a self-evaluating scale. Photographs of both right and left hands, placed on a flat surface with palms uppermost, were taken from all participants (see Figures 1 and 2), and the 2D:4D ratios were then measured using the NIH Image Processing and Analysis in Java (ImageJ) software. All hand pictures were taken with a camera tripod to ensure the same distance from the hands.

Example of a female artist's hands

Example of a male artist's hands
Results
Measurements of the 2D:4D ratio were recorded twice, by independent judges, and reliability was high, r(98) = 0.92, p = 0.0001.
Mean ages of both visual artists and controls were similar for males (artists: M = 44.6 ± 16.72 [SD] years; controls: M = 45.36 ± 16.66 years; t(24) = 0.16, p = 0.87) but not for females (artists: M = 55.08 ± 16.52 years; controls: M = 34.88 ± 13.83 years; t(24) = 4.69, p = 0.0001). However, since 2D:4D ratios develop early in utero, appear at 2 years of age, and never change across the lifespan (Manning et al., 1998), the age factor does not seem to be relevant in digital proportion analysis.
Descriptive statistics (Ms and SDs) of male artists and male controls were as follows: Male artists had lower 2D:4D ratios than controls (right hand male artists: M = 0.97 ± 0.04; left hand male artists: M = 0.96 ± 0.04; right hand male controls: M = 0.99 ± 0.04; left hand male controls: M = 1.01 ± 0.03). The mean difference between male artists and controls was significant (right hand: t(24) = 2.54, p = .015; left hand: t(24) = 4.28, p = .0001; see Figure 3). Female artists also had lower 2D:4D ratios than controls (right hand female artists: M = 0.96 ± 0.03; left hand female artists: M = 0.97 ± 0.03; right hand female controls: M = 1.00 ± 0.03; left hand female controls: M = 1.00 ± 0.03). The mean difference between female artists and controls was significant (right hand: t(24) = 3.87, p = .0001; left hand: t(24) = 3.77, p = .0001; see Figure 4).

Distributions of 2D:4D ratio in the right and left hands of 25 male artists (Experimental) and 25 male non-artists (Control)
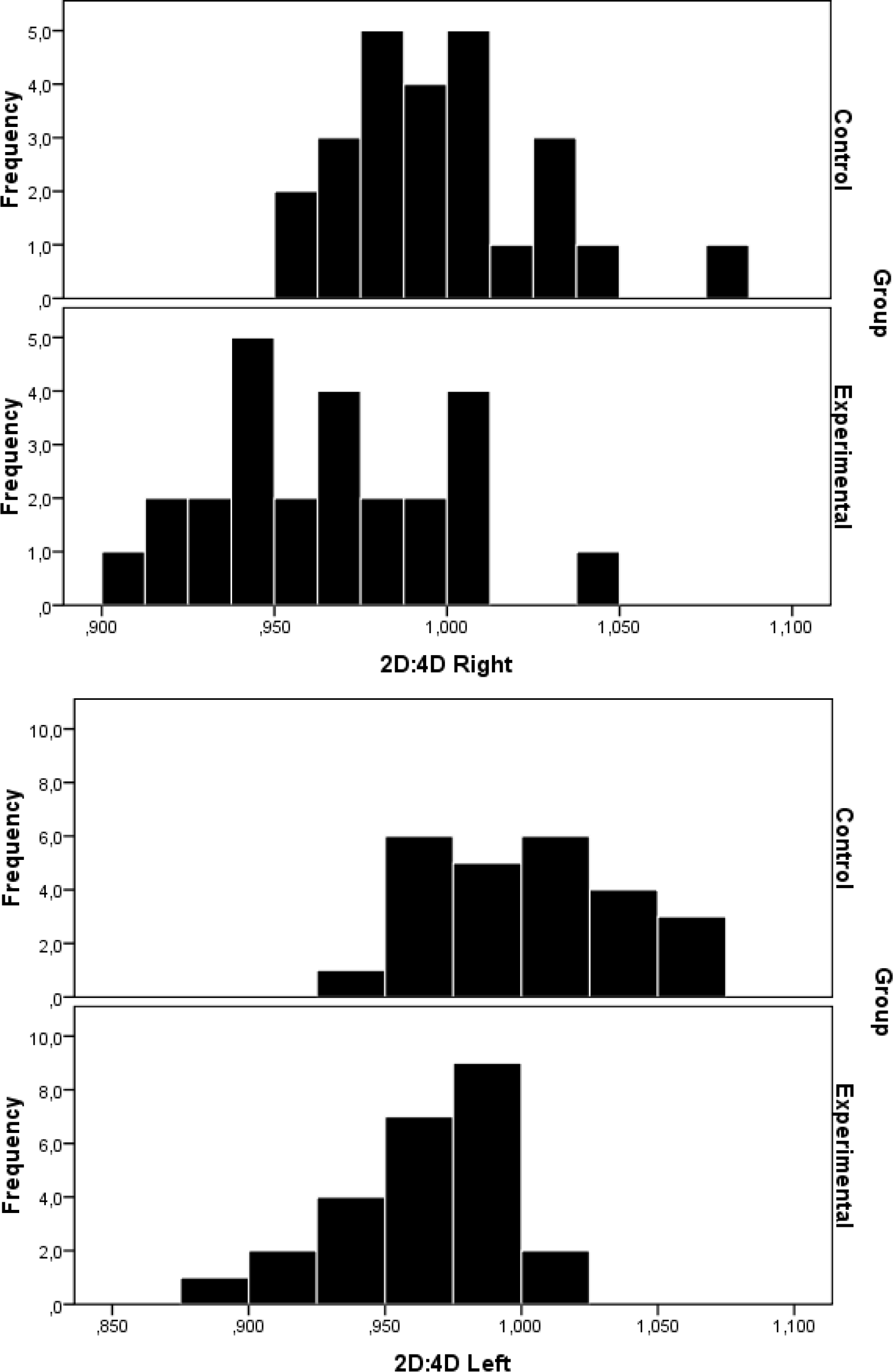
Distributions of 2D:4D ratio in the right and left hands of 25 female artists (Experimental) and 25 female non-artists (Control)
Overall, the mean 2D:4D ratios for right (see Figure 5) and left hand (see Figure 6) were significantly different between artists and non-artists. Specifically, lower 2nd 2D:4D ratios were recorded among the artist sample compared to controls.

Distribution of 2D:4D ratio in the right hand of both groups (50 artists [Experimental] and 50 non-artists [Control])
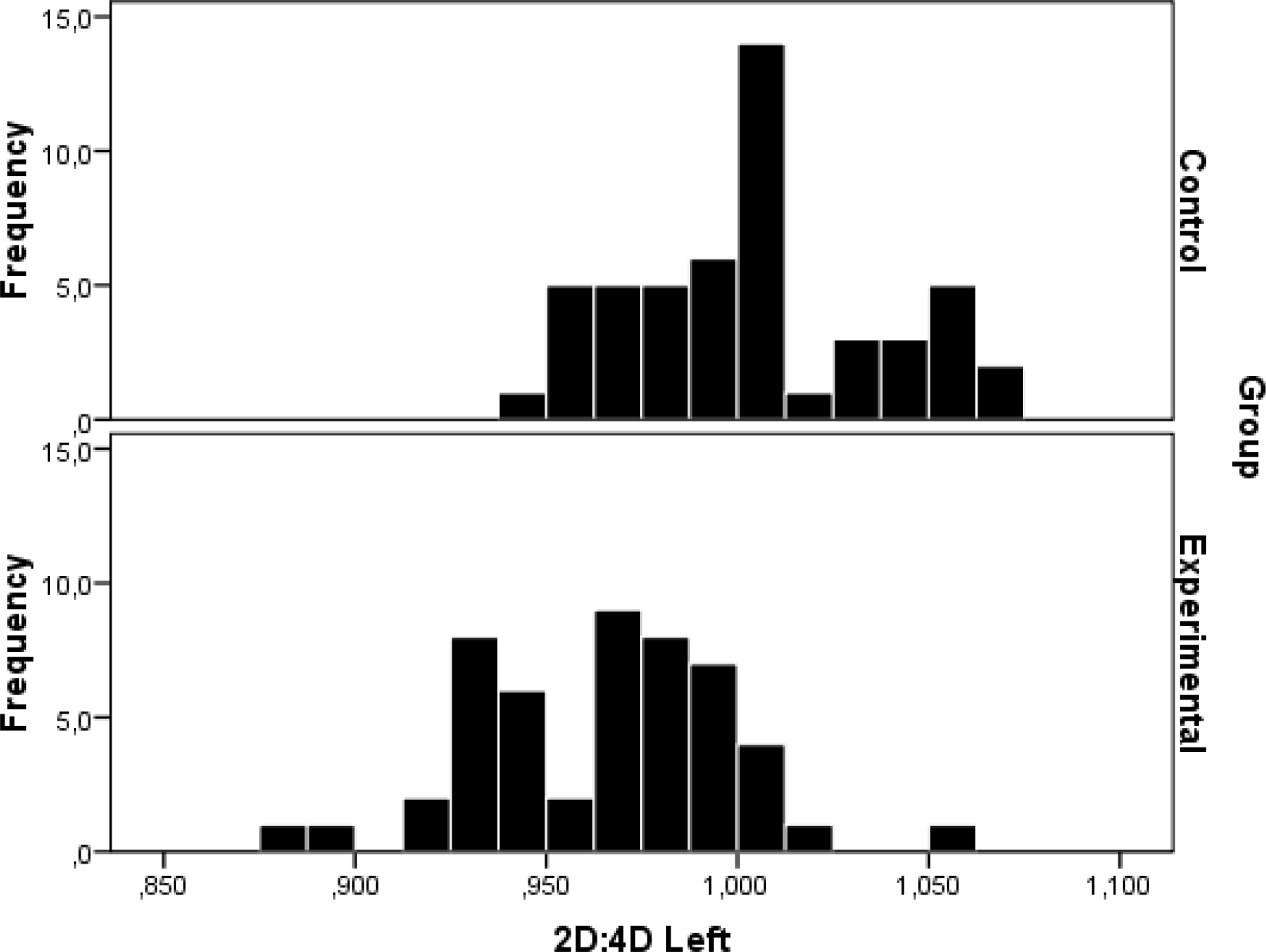
Distribution of 2D:4D ratio in the left hand of both groups (50 artists and 50 non-artists)
Surprisingly, no significant differences between the mean digit ratios of male and female artists were recorded in our sample (right hand: t(24) = .07, p = .945; left hand: t(24) = .08, p = .935). This suggests that the hypothesized higher masculinity exhibiting trait is greater in female rather than in male artists. Also, no significant differences were recorded between male and female controls (right hand: t(24) = .61, p = .54; left hand: t(24) = .96, p = .54).
Moreover, there was no evidence of a significant correlation between artistic ability and masculinity considering the results of the Bem Sex Role Inventory questionnaire. The scores for femininity (FS) and masculinity (MS) were as follows: Among both groups the MS was found to be slightly higher in men (5.05 ± 0.74) than in women (4.94 ± 0.66), although the difference was not significant, t(49) = .80, p = .423; the FS was found to be higher in women (5.10 ± 0.70) than in men (4.70 ± 0.64), and this difference was significant, t(49) = 2.99, p = .003. Comparing the two groups, both the MS and the FS appear to be slightly, but not significantly, higher among artists (MS artists: 5.05 ± 0.69; MS controls: 4.94 ± 0.71; t(49) = .85, p = .399; FS artists: 4.99 ± 0.63; FS controls: 4.81 ± 0.76; t(49) = 1.33, p = .186).
Among the artist sample, there was no evidence of a significant association between an increasing professional level and a decreasing 2D:4D ratio. In fact, given the small sizes of some of the groups composing the artistic level grading scale, it was not possible to associate 2D:4D ratios to either increasing or decreasing artistic ability. However, there was a significant correlation between increasing artistic level and increasing number of visual art works sold, r(48) = .302, p = .05.
Finally, contrary to expectations that an increasing number of visual art works sold means a decreasing 2D:4D ratio, a non-significant correlation was recorded. In fact, applying a multiple regression test, we determined that 2D:4D, sex, and age combined are poor predictors of the number of works sold (right hand: R2 = .104; left hand: R2 = .059).
Discussion
As the results demonstrate, our sample of visual artists had lower 2D:4D ratios compared to controls. However, decreasing 2D:4D ratio was not associated with increasing artistic ability, without relevant sex difference, and no significant relation between visual art works sold and lower 2D:4D ratio was recorded. On the other hand, there was quite a significant correlation between increasing artistic level and increasing number of visual art works sold, which could be ascribed to a prolonged contact with the artistic field.
Our results are consistent with an evolutionary approach to the original role played by artistic behaviors, through the action of sexual selection. In a context of role distinction between sexes, artistic abilities could be considered as typically male traits selected over generations, through female unconscious choice, that lead to increased reproductive opportunities. Although the study did not address the quantitative issue regarding the male predominance in the artistic field, the fact that both female and male artists showed lower 2D:4D ratios than non-artists demonstrates that androgenic concentrations are positively correlated to artistic proclivity. Since high levels of testosterone in men advertise good genes, fitness, fertility, and dominance, heterosexual women, especially when they are ovulating, are evolutionarily predisposed to finding highly androgenic men attractive (Johnston and Franklin, 1993). In this perspective, cultural displays and, more specifically, art may signal a direct benefit to the female's offspring by advertising the male's good genes (Miller, 2000; Zahavi, 1975, 1977). The problems that arise with the theoretical models of genetic quality displays and mate strategies not only deal with the honesty in signaling, but also, and especially, with the socio-cultural bias. The statistically observed sexual dimorphism among artists, musicians, and scientists (Kanazawa, 2000), with a predominance of men carrying out these activities, could be due to the fact that women have historically experienced less access to such fields. Instead, if artistic behaviors are linked to prenatal testosterone, as the present study suggests, this strengthens the hypothesis of interdependence between art, cultural displays in general, and sexual selection.
Moreover, different phases of the menstrual cycle influence the desire for creative and artistic individuals in such a way that female fertility is positively correlated with a preference for creativity and artistic propensity in short-term relationships (Haselton and Miller, 2006). Thus, artistic outputs seem to act, for their carriers, as an indicator of good genes. In addition, male artistic productivity seems to depend on the age of the producer, peaking at young adulthood when sexual competition is greater (Miller, 1999). Productivity, together with percentage of income made from artwork and number of exhibitions, was associated with professional and social success amongst visual artists (Clegg, Nettle, and Miell, 2011). In our study, contrary to expectations, no correlation was found between decreasing 2D:4D ratios (i.e., higher male fertility/genetic quality) and increasing productivity, neither among male nor among female artists. This result could depend on the limited sample size (there were only 25 male and 25 female artists) and further studies, on a larger number of subjects and focusing on the correlation between 2D:4D ratio and productivity, are needed.
More difficult to explain, at least in terms of the “good gene” hypothesis, is the low 2D:4D ratio in female artists. “Masculinization” of female artists could in part explain, on the one hand, their smaller number in the general population compared to male artists; on the other hand, the low percentage of women with masculine traits in the general population could simply represent one end of the normal distribution curve of the corresponding underlying genes.
We already know that testosterone affects the development of brain regions responsible for song in birds. Both male and female birds hear their species-specific song as nestling, but only males of most songbird species sing as adults. Male birds use song to claim territory, compete with other males, and declare dominance. They also use songs to attract females, which suggests that females know the song of their species even if they don't sing. Why do they not sing? Do they lack the muscular or nervous system capabilities necessary to sing? Or do they simply lack the hormonal stimulus for developing the behavior? To answer these questions, female songbirds were injected with testosterone in the spring. In response, females developed their species-specific songs and sang the same way as males did (Nottebohm, 1980). In addition, and compounded to the previously mentioned data, during the nonbreeding season, the male brain areas that are involved in birdsong are similar in size to those of a female brain. This gives greater evidence to cultural display as our hypothesized exclusively-male courtship behavior. Also, Sluming and Manning (2000) found a positive association between prenatal testosterone and spatial abilities and musicality. Analogously, the present study suggests a close link between low 2D:4D ratio (hence high prenatal testosterone) and painting ability. In fact, analogous to Sluming and Manning, who examined the percentage of women present in the seats close to musicians during concerts, we could further test our hypothesis by, for example, examining inspection times per work of art of women and men in art galleries.
The assumption for considering art as a male-predominant behavior, by highlighting its relation to sexual selection, is viewing it not as an exaptation or a side effect of increased brain size and cranial capacity, but rather, as suggested by Miller (2000), as a costly adaptation able to accomplish important courtship functions. According to one school of thought, the ability to produce and appreciate art is a co-opted trait that, at the beginning, had a different primary adaptive function. From a different perspective, art is considered to be a trait originally selected for in the domain of natural environment-driven competition (i.e., a character able to enhance reproductive success chances). Whether one may lean towards the former hypothesis or towards the latter (even if, as George C. Williams noticed in 1966, every trait may be considered in terms of both adaptation and exaptation), it is hard to believe that artistic capacity would be maintained by natural selection if it were “useless” in terms of biological advantage, especially because of its evident costs (e.g., “wasting” time that could be allocated to the performance of other important tasks, improper use of neural space). Evolutionarily speaking, these costs can be outweighed by gaining reproductive advantages. In this case the male's good genes are advertised by costly, sexually selected behaviors such as artistic abilities that could represent for females honest indicators of the potential partner's genotypic and, hence, phenotypic quality.
Footnotes
Acknowledgements
I am grateful to Dr. Paolo Perciballi for his assistance in the statistical elaboration of the data. Also, I would like to thank the subjects, artists and non-artists, for allowing us to interview them and measure their digits.