
Abstract
Human reproductive and sexual behaviors are heritable and may represent integral life history traits that are likely partially subserved by the dopamine system. Two dopamine receptor polymorphisms, DRD4 48bp VNTR and DRD2 TaqI A, were examined in relation to the Sexual-Orientation Inventory (SOI), age at first sexual intercourse, desired age of marriage, and desired age to have children in 195 (45% male) individuals from a general student population. As DRD4 7R alleles have been associated with migratory behavior, we also examined whether those with more 7R alleles had a greater frequency of multi-racial ancestries. Minor alleles of both polymorphisms (7R and A1 respectively) are believed to decrease the function of their respective receptors. Individuals with DRD4 7R alleles were more likely to have had sexual intercourse and to desire children earlier in life. In addition, DRD4 7R+ individuals were more likely to report multi-racial ancestries. Individuals with DRD2 A1 alleles were more likely to not want children and not want to marry. These results suggest that polymorphisms in the DRD4 and DRD2 genes are meaningfully associated with variation in reproductive and sexual behaviors. These results are provisionally interpreted as consistent with other findings suggesting that DRD4 7R and DRD2 A1 alleles are adaptive for lower offspring investment strategies in dynamic social environments.
Introduction
Among humans, genetic studies have shown significant heritable components to the age at which people first have sexual intercourse (Hunt and Rowe, 2002; Guo and Tong, 2006), begin trying to conceive (Rodgers, Kohler, Kyvik, and Christensen, 2001), and have their first child (Kohler, Rodgers and Christensen, 1999; Mealey and Segal, 1993), along with how many children they desire, expect and actually have (Kohler and Christensen, 2000; Kohler, et al., 1999; Rodgers et al., 2001). Additionally, the propensity to marry (Johnson, McGue, Kruger, and Bouchard, 2004), histories of maintaining lasting pair bonds (Kohler and Christensen, 2000; Trumbetta and Gottesman, 2000) and propensities for multiple mating (Trumbetta and Gottesman, 2000) have heritable components. Recent findings show that reproductive outcomes become more heritable with increasing female reproductive choice, suggesting that the heritable variation acts not primarily via modifications of physiological ability to reproduce, but instead via behavioral influences (Kohler, Rodgers, Miller, Skytthe, and Christensen, 2006).
The dopamine system is a good candidate system to modulate such behaviors because it is involved in reward and motivation, and has been implicated in sexual (Dominguez and Hull, 2005; Melis and Argiolas, 1995; Melis et al., 2006; Succu et al., 2007) and pair bonding behaviors (Aragona et al., 2006; Curtis and Wang, 2005; Fisher, Aron, and Brown, 2005; Gingrich, Liu, Cascio, Wang, and Insel, 2000; Smeltzer, Curtis, Aragona, and Wang, 2006). Two particularly promising genes that might be behaviorally associated with reproductive behavior are those of the dopamine receptors D4 and D2 (DRD4 and DRD2 respectively). Some empirical evidence supports this, albeit preliminarily. Polymorphisms of the DRD4 gene have been associated with sexual desire, arousal and function (Zion et al., 2006). Long alleles of the DRD4 48 bp VNTR polymorphism have been associated with increased sexual novelty (Hamer, 2002). DRD4 48 bp VNTR 3R+ genotypes have been found related to age at first sexual intercourse in one study (Guo and Tong, 2006) although DRD4 48 bp VNTR was unrelated to age at first sex in another study (Miller et al., 1999). In the DRD2 gene, the intron5/exon6 2-haplotype has been associated with earlier age at first sexual intercourse among U.S. couples (Miller et al., 1999; however MacMurray, Madrid, Bottini, Muhleman, and Comings, 2002) and decreased number of children among Hispanic women (Legro et al., 1994). In a university population, the DRD2 TaqI A A1 allele has been associated with having more children in those under 35 years old, but fewer children in those over 35 (Comings, 2000). In a group of adult male Mayan Indians, A1 carriers had an earlier age at the birth of their first child and increased total fertility (MacMurray et al., 2002).
The preceding findings are suggestive of genetic influences on reproductive behavior via these dopamine receptor genes, but are clearly far from definitive. In the current study we explored how DRD4 48 bp VNTR and DRD2 TaqI A genetic polymorphisms relate to sexual and reproductive behaviors. Specifically, we examined how restriction in sexual behavior as measured by the Sexual-Orientation Inventory (SOI), age at first sexual intercourse, desired age at first child, and desired age at first marriage associated with these genetic polymorphisms. Based upon evidence that minor alleles of these polymorphisms are associated with greater reactivity to dopaminergic rewards (Hutchison, McGeary, Smolen, Bryan, and Swift, 2002b; Hutchison, LaChance, Niaura, Bryan, and Smolen, 2002a; McGeary et al., 2006; Noble, 2003), we predicted that possession of the DRD4 48 bp 7 repeat (7R) allele and DRD2 TaqI A A1 allele would be associated with less restricted reproductive behavior, or more “r-selected” life history strategies.
Interestingly, 7R alleles of DRD4 48bp VNTR are present at higher frequencies in populations that have migrated farther (Chen, Burton, Greenberger, and Dmitrieva, 1999). The genetic structure of this same “migratory” 7R allele suggests that it originated and was positively selected for between 40 and 50 thousand years ago (Wang et al., 2004). Therefore, we tested a novel hypothesis that the geographical patterns of DRD4 distribution may be due to differences in reproductive behaviors. Those exhibiting more migratory propensities may be more likely to explore novel geographic and social environments, putting them in contact with more potential mates of other races. As such, we predicted that 7R alleles would be associated with more multi-racial ancestries.
Materials and Methods
Participants
Between February and April 2005 a total of 195 subjects were recruited for participation from the Human Subject Research Pool at the State University of New York at Binghamton as part of a larger series of studies examining the associations of genetic polymorphisms of the dopamine system with behavior (Eisenberg et al., 2007). No screening measures were placed on who could participate. Because population stratification is a potential problem in genetic association studies (Hutchison et al., 2002b; Hutchison, Stallings, McGeary, and Bryan, 2004), race was closely examined by asking participants to identify the ancestry of all four of their grandparents as has been recommended by Shields et al. (2005). Participants were allowed to select as few or as many ancestry groups to describe each grandparent from the following categories: European, African American, East Asian, South Asian, Middle Eastern, Native North American, Native South American, Pacific Islander, African and an open ended “other” category.
Candidate Genes and Genotyping
DRD4 48 bp VNTR
The DRD4 48-bp VNTR polymorphism is in exon 3 of the gene coding for the dopamine D4 receptor. The VNTR polymorphism varies between 2 and 11 repeats of a similar 48 bp coding region sequence, with a tri-modal distribution of 2, 4 and 7 repeat alleles (2R, 4R and 7R) in most, but not all, populations (Ding et al., 2002). Although the functional significance of the DRD4 VNTR polymorphism has not been definitively characterized, long alleles (typically 7R as opposed to 4R) have been generally found to be functionally less reactive in in-vitro expression experiments (Asghari et al., 1995; Czermak, Lehofer, Liebmann, and Traynor, 2006; Schoots and Van Tol, 2003; Van Craenenbroeck et al., 2005; Van Tol et al., 1992), with some heterogeneity (Asghari et al., 1994; Cho, Beorn, Van Tol, Caron, and Kim, 2006; Jovanovic, Guan and Van Tol, 1999; Oak, Lavine and Van Tol, 2001; Watts et al., 1999). Additionally, in vivo human pharmacological studies are also generally consistent with the notion that 7R alleles are associated with less responsive D4 receptors than 4R alleles (Brody et al., 2006; Hamarman, Fosella, Ulger, Brimacombe, and Dermody, 2004; Hutchison et al., 2003; Hutchison et al., 2006; McGough et al., 2006).
DNA was collected with QuickExtract buccal swabs and extracted with BuccalAmp solution as directed by the manufacturer (Epicenter). Subjects were instructed to rinse their mouths out with water before swabbing. The DRD4 48bp VNTR locus was genotyped using an adaptation of a previous protocol (Boór et al., 2002).
The DRD4 48bp VNTR PCR reaction contained 1x Q-Solution (Qiagen), 1x Buffer (Qiagen), 1 μM Primer 1 (5′ GCGACTACGTGGTCTACTCG 3′), 1 μM Primer 2 (5′ AGGACCCTCATGGCCTTG 3′), 200 μM dATP, dTTP, dCTP and 100 μM dITP and dGTP, 0.3 units HotStar Taq (Qiagen), 1 μl of DNA template, in a total volume of 10 μl. Thermocycler conditions were: 15 min at 95° to activate the enzyme and denature the DNA, 40 cycles of 1 minute denaturation at 94°, 1 minute annealing at 55°, 1.5 minute extension at 72°, followed by one cycle of 10 minute extension at 72°. A 4R PCR product is 475 bp. Samples were visualized under UV on 1.4 – 2.0% ethidium bromide agarose gels with a 100 bp ladder. Some gels were placed in a 1x boric acid bath for 10–15 minutes to increase contrast and allow better visualization of 9R alleles. With different concentrations of DNA template we observed allelic dropout of both short and long alleles. Because of these problems of allelic dropout and a previous report of only 84% correspondence between DRD4/48bp genotype techniques (Hamarman et al., 2004), all samples were run at 1/1, 1/40 and 1/80 dilutions for confirmation. If a minor allele was consistently observed at any dilution, the subject was considered to have that allele. DRD4 48bp genotyping had a 99% success rate with one subject who yielded no PCR product and another whose results varied between runs.
DRD2 TaqI A
The DRD2 TaqI A site is a single-nucleotide polymorphism with a major A2, and minor A1 allele. The A1+ genotype (heterozygous or homozygous A1) has been most strongly associated with substance abuse, particularly alcoholism, albeit with some ambiguity (Noble, 2003). The A1+ genotype has also been related to pathological gambling, novelty seeking, and, the related psychological construct, sensation seeking (Noble, 2003; Ratsma, van der Stelt, Schoffelmeer, Westerveld, and Gunning, 2001). Although it is unclear why, the A1 allele is associated with decreases in dopamine D2 binding and glucose metabolic rates in many brain regions (Blum et al., 2000; Noble, 2003; Thompson et al., 1997). The DRD2 TaqI A locus is 9.4 kb downstream from the coding region for the DRD2 gene, is not in any known regulatory region, and has no known mechanism to influence DRD2 expression. The DRD2 2-haplotype of the intron5/exon6 is in linkage disequilibrium with the DRD2 TaqI A locus, but the nature of the linkage remains to be elucidated (MacMurray et al., 2002). The DRD2 exon 6 polymorphism is silent (ALFRED-The Allele Frequency Database) and the intron 5 polymorphism is not known to be functional, so their linkages to TaqI A does not explain the functional associations of TaqI A. The TaqI A polymorphism is also in a nearby kinase gene, the Ankyrin Repeat and Kinase Domain Containing 1 (ANKK1) gene, where it causes a Glutamate →Lysine substitution (Dubertret et al., 2004; Neville, et al., 2004). The results of the amino acid substitution are not known, but could impact interactions of ANKK1 proteins with other proteins including the dopamine D2 receptor (Neville, Johnstone and Walton, 2004). No other polymorphism has been revealed to be in linkage disequilibrium with TaqI A that could clearly account for its functional associations (Dubertret et al., 2004; Neville, et al., 2004). DRD2 TaqI A was typed with a PCR/RFLP method (based on GSFL Research Group, 2003).
The DRD2 TaqI A PCR reaction contained 0.5 μM forward (5′ CAC GGC TGG CCA AGT TGT CTA 3′), 0.5 μM reverse primers (5′ CACCTTCCTGAGTGTCATCAA3′), 200 μM dNTP, 2.5 mM MgCl2, 0.25 units HotStar Taq (Qiagen), 1X Buffer (Qiagen), 2.5 μl DNA template in a total volume of 10 μl. Cycle conditions were: 15 min denaturation at 95°C, 40 cycles of 30 s denaturation at 94°, 30 s annealing at 55°, 1 min extension at 72° and one final extension of 7 minutes at 72°. The PCR product was digested with TaqI enzyme (New England Biolabs) overnight at 65° and visualized under UV on a 1.4% ethidium bromide agarose gel with 100 bp ladder. The 300 bp PCR product is not cut by the restriction enzyme in A1 alleles, and A2 alleles yield a 125 and 175 bp fragment. DRD2 TaqI A was typed successfully in 100% of the subjects.
Phenotypic Measures: Sociosexual Orientation Inventory (SOI)
The SOI measures restriction of sexual and pair-bonding behaviors. It has been validated on student populations around the world (Schmitt, 2005; Simpson and Gangestad, 1991). Those who score lower on the SOI generally engage in sex later in relationships that are “characterized by reliably greater expressed love, dependency, commitment and investment (Simpson and Gangestad, 1991, p. 876).” The SOI generally corresponds with k versus r-selected reproductive strategies (high versus low parental care/investment strategies; Gangestad and Simpson, 1990; Simpson and Gangestad, 1992; Simpson, Gangestad, Christensen, and Leck, 1999). The SOI was slightly altered to differentiate hetero and homosexual activity and sexual identity, but because the sample contained only one subject self-identified as bisexual and four self-identified as homosexual, they were not distinguished from those self-identified as heterosexual in the further analyses.
Additional survey items
Subjects were asked their desired age to marry, have their first child, or whether they did not want to marry or have children. They were also asked their age at first sexual intercourse, or if they were virgins.
Procedures
All procedures were approved by the Human Subjects Research Review Committee at the State University of New York at Binghamton and all subjects gave informed consent. Participants attended group assessment sessions (with a maximum number of 10 subjects per group) and were initially provided with oral instructions followed by the DNA sample collection. All data were collected via personal computers (Macintosh G4s) in a computer lab using the database FileMaker (FileMaker, Inc.). Subjects were seated at least every other seat to assure anonymity. In addition to the oral instructions, all measures were accompanied by on-screen written instructions and experimenters (DTAE and/or MM) were available throughout the sessions for questions. The sessions lasted approximately one hour.
Data Analysis
All data was examined for outlying data points, distribution normality and missing values. To assure that missing responses were not systematically biased by genotype, missing versus non-missing data for each self-report scale was analyzed across both DRD4 VNTR and DRD2 TaqI A genotypes via 2 × 2 contingency tables using chi-square or Fisher's exact statistics. Hardy-Weinberg (HW) equilibria were tested with the HWE program (Brzustowski, 2007). The DRD4 HW equilibrium was tested with the Markov Chain algorithm and the DRD2 HW equilibrium was tested with Fisher's exact test.
Based on previous association studies, those with at least one seven repeat allele at the DRD4 48bp VNTR locus were designated 7R+, while those without seven repeat alleles were designated 7R-. In terms of DRD2 TaqI A, individuals with at least one A1 allele were designated as A1+ and those who were homozygous for the A2 allele were designated A1-. The principal analyses used 2 (7R+/7R+) × 2 (A1+/A1-)× 2 (Male/Female) factorial analyses of variance (ANOVAs). Chi-square or Fisher's exact tests were used to analyze categorical data. Significant main effects were evaluated to see if the minor alleles have the assumed dominant effect or an alternative additive, recessive or overdominant effect. Population stratification was addressed by re-examining the significant results within the largest single racial group (i.e., a group with minimized racial admixture); in this case, it was the European ancestry group. Rather than emphasizing statistical significance based on F-ratio or □2 value, due to the inherently diminished power by reduced sample size, comparisons were made between effect sizes in the findings from the overall sample and the subsample. Since this study employs multiple phenotypic scales, the potential exists for type I errors. For several reasons we have not employed a correction of our significance criteria for multiple testing (Bland, 2000; Perneger, 1998). The diversity of scales employed in this study should not decrease the sensitivity of analysis. In addition, because of the exploratory nature of our analysis (Perneger, 1998), and because some phenotypic variables are correlated (see Table 5), corrections for multiple tests are too conservative (Bland, 2000). Nonetheless, it is important for the reader to bear this caution in mind as they interpret the results. A two-sided significance criterion of p ≤ .05 was used throughout.
Results
Sample Background
The sample is described demographically in Table 1. It had a roughly equal sex ratio, a narrow age range (as expected in a college population) and was predominately of European descent. Because of the small spread in age, age was generally not included as a covariate. Genotype and allele frequencies are given for DRD4 48bp VNTR and DRD2 TaqI A in Tables 2 and 3 respectively. Genotype frequencies were comparable to other samples of mixed populations with predominately European descent. Both loci were in Hardy-Weinberg Equilibrium (DRD4 VNTR: Markov Chain algorithm, p = 0.38; DRD2: Fisher's exact, p = 1.0).
Sample characteristics, (n = 195). Subjects were undergraduate students recruited through a psychology department's human subject pool.

DRD4 48 bp VNTR allele frequencies, genotypes and genotype classifications. Allele frequencies are unremarkable for a mixed U.S. population. Genotypes were in Hardy-Weinberg Equilibrium (Markov Chain algorithm, p = 0.38)

The 7R+ genotype classification includes those with at least one seven repeat allele and 7R- reflects those without any seven repeat alleles. DRD4 status could not be ascertained from two participants.
DRD2 TaqI A allele frequencies, genotypes and genotype classifications. Allele frequencies are unremarkable for a mixed U.S. population. Genotypes were in Hardy-Weinberg Equilibrium (Fisher's exact, p = 1.0).

The A1+ genotype includes both A1/A1 and A1/A2 while A1- denotes the A2/A2 genotype.
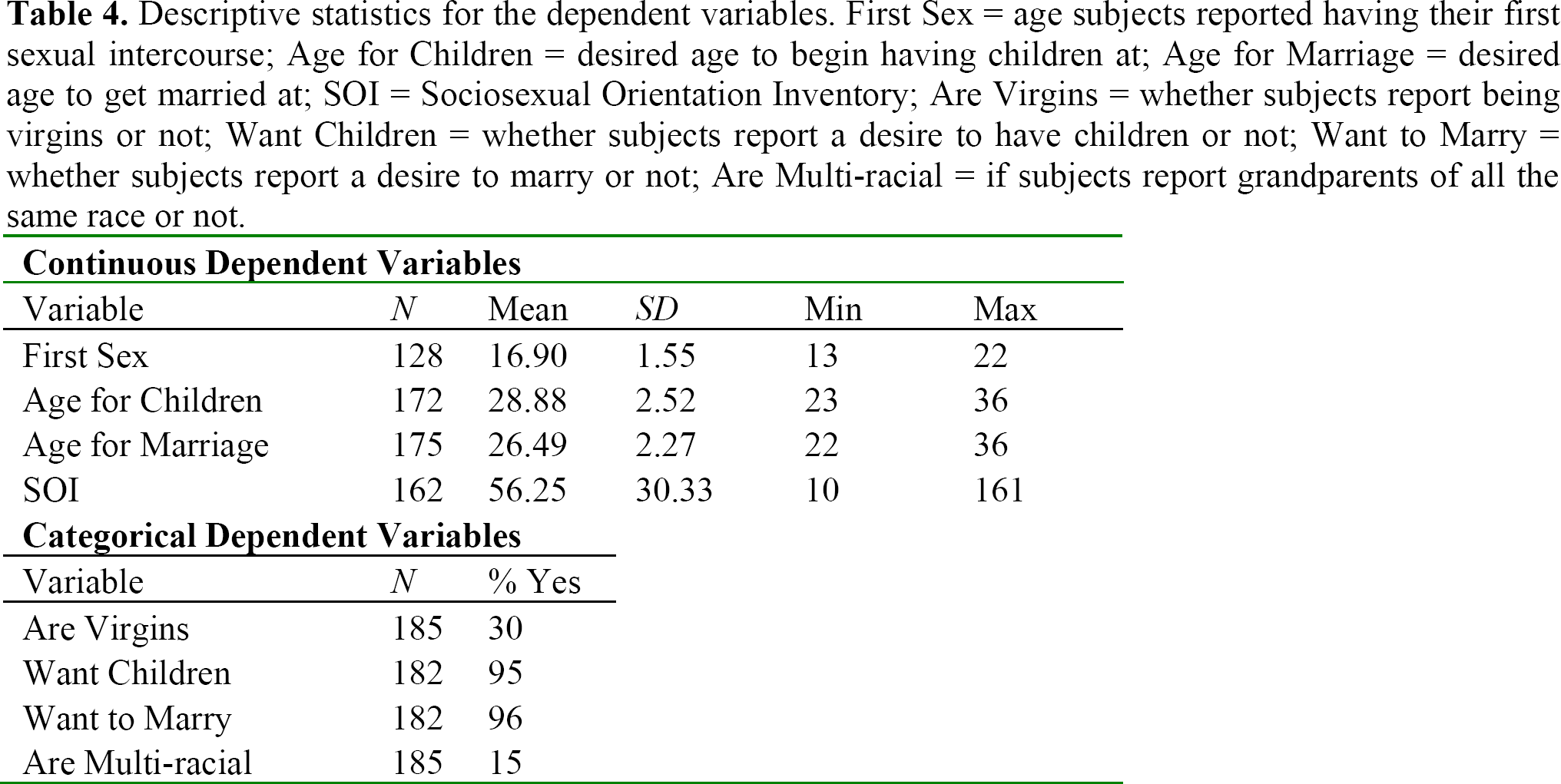
Dependent variables are described in Table 4 and each continuous variable is given with its inter-correlations with the other variables in Table 5. Unsurprisingly, the desired age for marriage and children were highly correlated and less restrictive socio-sexual profiles were associated with earlier age at first sexual intercourse.
Descriptive statistics for the dependent variables. First Sex = age subjects reported having their first sexual intercourse; Age for Children = desired age to begin having children at; Age for Marriage = desired age to get married at; SOI = Sociosexual Orientation Inventory; Are Virgins = whether subjects report being virgins or not; Want Children = whether subjects report a desire to have children or not; Want to Marry = whether subjects report a desire to marry or not; Are Multi-racial = if subjects report grandparents of all the same race or not.
Pearson's product-moment correlations between continuous dependent variables. SOI = Sociosexual Orientation Inventory

p<.01
p<.05
Missing Values Analysis
Since more missing data by one genotype than another might alter subsequent analyses, all scales were examined for biased missing data. Missing values were due to subjects skipping one or more items on self-report scales or incorrectly entering subject ID numbers. The only scale that showed biased missing responses was the SOI, in which 7R- subjects were less likely to answer than 7R+ (80.5% and 91.7% respectively; ηp.2 = 3.858, df = 1, p = .050, N = 193) and A1- individuals were less likely to complete the scale than A1+ individuals (78.1% and 88.8% completed respectively; ηp.2 = 4.117, df = 1, p = .042, N = 195). Post-hoc, binary logistic regression revealed no significant DRD4 by DRD2 interaction effect on SOI missing data (results not shown). Since it is striking that missing data was biased by both DRD4 and DRD2 on the SOI, we further examined missing values for each item of the SOI individually. The only individual item to show bias was item 4, which asks subjects “How often do you fantasize about having sex with someone other than your current dating partner?” A1- subjects were more likely to omit item 4 than A1+ subjects (ηp2 = 5.950, df = 1, p = .015, N = 195).
Children and Marriage
Individuals with 7R+ genotypes reported desire for children slightly earlier than 7R- subjects (28.26 years of age versus 29.18 respectively; F [1,162] = 5.184, p = .024, ηp2 = .031). The effect size was over twice as large with nearly the same level of significance in the European-only analysis (F [1, 61] = 5.198, p = .026, ηp2 = .079). As shown in Table 6, the DRD4 association with desired age at first child tentatively reflects an additive effect of 7R alleles. Marriage age, which is highly correlated with age at first child (Table 5), was only marginally associated with the 7R+ genotypes (F [1,165] = 3.054, p = .082) and in the same direction as the association with desired age at first child.
Desired age at first child by DRD4 genotype. 7R+ genotypes reported desire for children slightly earlier than 7R- subjects (28.26 years of age versus 29.18 respectively; F [1,162] = 5.184, p = .024, ηp2 = .031). The effect appears to be additive in that increasing numbers of 7R alleles are associated with decreasing desired age of first child.

Significantly more A1+ participants reported that they did not want to have children than A1- (Fisher's exact, p = .002, CC = .230). Indeed, all ten subjects who reported not wanting to have children were heterozygous at the DRD2 locus. Similarly, all seven subjects who reported not wanting to get married were DRD2 heterozygous (A1+ versus A1- Fisher's exact, p = .014, CC = .190). In European-only analyses, A1+ subjects still had less desire to have children and marry with larger Contingency Coefficients (CCs; children, Fisher's exact, p = .016, CC = .282; marriage, Fisher's exact, p = .089, CC = .219). Since all who did not want children were heterozygous and all homozygous subjects wanted children, an overdominance inheritance pattern is suggested. However, with only 17 A1 homozygotes (Table 2), the certainty of overdominance is low. There were no other significant effects of gender, genotype or interactions thereof on desire for children or marriage (ps > .086).
Sexual Behaviors
As expected, males reported less sexual restriction on the SOI than females (males = 67.19, females = 48.91; F [1,153] = 13.968, p < .001, ηp2 = .084). Contrary to predictions, there were no genotype-SOI associations (not shown). However, the results on the SOI should be viewed in light of the significant bias in missing data on this scale with respect to both genetic loci. There were fewer virgins than expected in 7R+ subjects (16.1%) relative to 7R- subjects (36.2%; ηp.2 = 7.505, df = 1, p = .006, CC = .198). Since age (despite its narrow range) could have an obvious relationship to virginity status, it was entered as a covariate in a binary logistic regression model, where the association between DRD4 and virginity remained (p = .008). The CC of DRD4 on virginity status was slightly smaller among Europeans than the pooled population (7R+ v. 7R-: Fisher's exact, p = .352, CC = .143). Similar to desired age at first child, when analyzing virgin status the 7R allele tentatively has an additive effect (Table 7). There were no other significant effects of gender, genotype or interactions thereof on SOI, age at first sex, or virginity status (ps > .159).
Virginity status by DRD4 genotype There were fewer virgins than expected in 7R+ subjects (16.1%) relative to 7R- subjects (36.2%; ηp.2 = 7.505, df = 1, p = .006, CC = .198) and the effect appears to be additive such that increasing numbers of 7R alleles is associated with a decreased likelihood of being a virgin.

Reproduction and Migration
Consistent with the hypothesis that DRD4 48bp would be related to multi-racial ancestry, we found that 7R alleles were over-represented in those subjects with biracial and tri-racial ancestries (Table 8; Fisher's exact, p = .002). Other methods described in the appendix were used to eliminate some possible biases and were also supportive of the multi-racial coupling hypothesis.
DRD4 and multi-racial ancestry. 7R alleles were over-represented in those subjects with biracial and tri-racial ancestries (Fisher's exact, p = .002). Number of races is judged by self-identified ancestry of each of four grandparents. Percentages of row totals are given in parenthesizes.

Discussion
The current study sought to examine the relevance of polymorphisms of the dopamine D2 and D4 receptor genes to variation in human reproductive and sexual behavior. The broad hypothesis was that minor alleles of these genes would be associated with less restricted reproductive behavior on an array of indices. The results generally supported that notion, although the findings were mixed with regard to the two genes and specific indices examined. DRD4 VNTR 7R+ status was associated with interest in reproducing at a slightly earlier age, earlier initiation of sexual activity, and multi-racial ancestry, whereas DRD2 TaqI A A1+ status was associated with reporting no interest in marriage or having children. Notably, these findings were robust when only individuals with European ancestry were included in the analysis, suggesting little influence of population stratification. Contrary to predictions, no associations were evident between genotypes and performance on the SOI. Of interest, the differential associations observed between the two genes and behaviors, and the absence of epistatic interactions, suggests that these polymorphisms have different influences. However, these findings should all be viewed in light of the exploratory nature of the study and our decision not to correct for multiple statistical tests (see methods).
In terms of their consilience with the small existing literature on these polymorphisms and reproductive behavior, the DRD4 VNTR findings both converge with and diverge from previously reported findings. The results are consistent with past associations of the DRD4 gene with sexual desire, arousal and function (Zion et al., 2006). However, Miller et al. found no relation between DRD4 48 bp and age at first sex in a group of married couples (Miller et al., 1999), while Gou and Tong found that 3+ genotypes (but not 7R+) were associated with earlier first sex among a general United States population sample (Guo and Tong, 2006). The effect of DRD4 may be different in the subpopulation of college students than the more general populations represented among the other two studies (Guo and Tong, 2006; Miller et al., 1999).
With regard to the results relating DRD4 48bp VNTR status to multi-racial ancestry, Chen et al. previously found that populations that have migrated further over the past 1,000 to 30,000 years have a higher frequency of DRD4 7R alleles (1999). Consistent with this, we found that minor alleles of the DRD4 48bp VNTR polymorphism are associated with more multi-racial ancestry. However, the mechanisms to explain the migratory patterns observed by Chen et al. remain unclear. The reason 7R DRD4 48bp VNTR alleles are over-represented in more migratory populations might not be because 7R+ individuals migrate more than the general population, but instead that they are more successful after migration. Speculatively, this success could come from appreciation for novel aspects of the environment, including mates.
In terms of the DRD2 TaqI A findings, the results are inconsistent with two past studies that found A1+ subjects have more children at an earlier age (Comings, 2000; MacMurray et al., 2002). In one of those studies A1+ subjects less than 35 years of age had more children, but those over 35 had fewer (Comings, 2000), suggesting a complex nature to the association of DRD2 with reproductive desires and outcomes. These contradictory results might be resolved by considering the distinction between desire for children and actually having children. It is possible that A1+ individuals who express less interest in having children nonetheless practice behaviors that make them more likely to have children early. However, at this point, such an interpretation must be speculative. Thus, these data may be consistent with the notion of the A1 allele being associated with a low offspring investment reproductive strategy, but the definitive relationship is not clear at this point.
As noted above, contrary to our original prediction, the SOI scores did not differ by genotypes. This could be because aspects of sexual behavior assessed by the SOI are not relevant to aspects of the two polymorphisms examined, but could also be due to biases introduced by missing data for the SOI. The SOI was the only scale to show biased missing data at both loci. Of note, Item 4, which assumes a current dating partner and was biased by genotype, has also been noted as problematic in past studies which found it to be the most frequently omitted on the SOI (Clark, 2006). Possible explanations for these omission biases include that possession of these minor alleles could be associated with being less reluctant about reporting specific sexual experiences on surveys or being differentially likely to have a dating partner; those who were not dating might have chosen to omit Item 4. The advertisement for this experiment did not mention that participants would be asked questions of a sexual nature, but other studies which are more forthright about this (e.g. Zion et al., 2006) might bias their subject pools.
In addition to missing data with regard to the SOI, there are two additional limitations to the current study that are worthy of discussion. First, the study was relatively small and used a sample that was probably not representative of the general population. Second, the sample was within a relatively circumscribed age range, with virtually all participants being young adults. As a result, it is possible that some data suffered from restricted range, such as virginity status. Given these aspects of the design, these findings should be most prudently interpreted as being specific to this sample and require further validation in larger, more representative samples.
In spite of these qualifications, these findings nonetheless converge with recent evolutionary theoretical approaches to the adaptive nature of DRD4 and DRD2 genetic polymorphisms (Campbell and Eisenberg, 2007; Harpending and Cochran, 2002). Minor alleles of DRD4 and DRD2 are associated with traits that ostensibly decrease fitness, including substance abuse, schizophrenia, movement disorders, migraines (Noble, 2003; Vandenbergh et al., 2000), disorganized infant attachment (Gervai et al., 2005; Lakatos et al., 2002; Van Ijzendoorn and Bakermans-Kranenburg, 2006), and ADHD (Li, Sham, Owen, and He, 2006). However, these minor alleles have been maintained in many populations and, in the case of DRD4 48bp 7R, there is evidence of positive selection (Wang et al., 2004).
It is possible that these risk-conferring alleles are more adaptive in dynamic environments. In a dynamic social environment or changing ecological environment, ideas acquired from family members may be less innovative and adaptive than those ideas acquired from a broader survey of society (Richerson and Boyd, 2005). Perhaps consistent with wider surveying for ideas, DRD4 48bp 7R+ individuals have increased propensities for novelty-seeking (Savitz and Ramesar, 2004; Schinka, Letsch and Crawford, 2002; but Kluger, Siegfried and Ebstein, 2002), cognitive flexibility (Dreisbach et al.), higher activity levels (Langley et al., 2004), better sexual functioning (Zion et al., 2006) and may have faster response times (Langley et al., 2004; Swanson et al., 2000). Especially notable, while externalizing behaviors are normally negatively related to IQ, among 7R+ subjects (but not 7R-) there is no relationship between externalizing behaviors and IQ (DeYoung et al. 2006). 7R+ individuals may be characterized by putting less value on familial influences and being more readily able to take advantage of dynamic social environments. Our evidence of younger desired age at first reproduction, earlier first sexual intercourse and more multi-racial ancestries among 7R+ individuals is generally consistent with a higher mating effort and lower offspring investment strategy suiting a dynamic social environment with less dependable family influences. Inter-racial marriage could provide new networks of kin support, and avenues for learning to take advantage of new environments. Similarly, although with less evidence, possession of a DRD2 A1 allele may adapt individuals better to such a low offspring investment, high mating effort strategy.
In summary, the current study suggests that polymorphisms in the DRD4 48bp VNTR and DRD2 TaqI A genes influence variation in human reproductive and sexual behavior. The findings generally support the notion that minor alleles of these genes are associated with lower offspring investment reproductive strategies, but had mixed overlap with the small number of previous studies in this area. In addition, these results converge with recent evolutionary perspectives on these minor alleles that emphasize both their fitness-enhancing and fitness-decreasing influences.
Footnotes
Acknowledgements
We gratefully acknowledge Joshua Beauchemin for assistance in genotyping, Stephen A. Lisman for assistance in conducting the study, Rita Spathis for lab assistance and advice, Kevin Sheridan, Zsolt Ronai and John McGeary for genotyping advice, David Schmitt for sexual scale suggestions, Justin Garcia for comments on drafts of the manuscript and the subjects for their time and participation. This study was supported in part by Undergraduate Research Awards from the Harpur College Awards Committee of the State University of New York at Binghamton to DTAE, MM, and DD; by a start up grant from the Research Foundation of Binghamton University to JKL; and by Grant T32 AA007459-20 from the National Institute on Alcohol Abuse and Alcoholism.