
Abstract
Background:
There is some evidence that components of the renin-angiotensin system and kallikrein-kinin system are not similarly regulated in both sexes. The aim of this work was to analyze the expression of angiotensin-converting enzyme, angiotensin-converting enzyme 2, angiotensin 1 receptor, angiotensin 2 receptor, beta-1 receptor, and beta-2 receptor during the evolution of myocardial infarction.
Methods:
Thirty-six male and 36 female Wistar rats were used. Myocardial infarction was induced. Six groups of both sexes were formed, (n=6): (a) sham; (b) 48 h myocardial infarction; (c) one week myocardial infarction; (d) two weeks myocardial infarction; (e) three weeks myocardial infarction and (f) four weeks myocardial infarction. The expression was evaluated by real-time polymerase chain reaction on the penumbra of left ventricle.
Results:
The mRNA expression of most biomarkers was lower in females than in males. During acute infarction, an increase of all protein expression was found in female and at two weeks while in the male only biomarker changes occurred at three weeks. In addition, in male biomarkers mRNA expression decreased during chronic infarction while in females it did not.
Conclusions:
The renin-angiotensin system and kallikrein-kinin system biomarkers expression occurs at earlier times in the female than in the male rat. In addition, during chronic myocardial infarction these biomarkers remained unchanged in females while in males they decreased.
Keywords
It is currently known that, after suffering a first episode of myocardial infarction (MI), death is more likely for women (26%) than for men (19%). 1 However, there is not enough information to understand the differences in the pathophysiology of cardiovascular diseases between the sexes. Due to this lack of information, some investigators have shown that the regulation of the renin-angiotensin system (RAS) and the kallikrein-kinin system (KKS) is influenced by sex hormones, as in the study by Owonikoko et al. 2 that showed a reduced number of angiotensin 1 (AT1) receptors in dogs that underwent ovariectomy. Roesch et al. 3 also reported downregulation of AT1 receptors during estrogen replacement therapy. Some studies have proven that, in addition to receptors, enzymes are affected by sex. For example, Zapater et al. 4 suggested that regulation of angiotensin-converting enzyme (ACE) activity may be influenced by estrogens or androgens that contribute to differences in blood pressure between the sexes. However, Gallagher et al. 5 found a decrease in ACE mRNA concentrations and a reduction in ACE activity in the kidney cortex, kidney medulla, and lungs after 21 days of 17β-estradiol replacement therapy in ovariectomized rats. The beta-1 (B1) receptor is an important biomarker in the KKS system, which has not yet been associated with sex during MI and is known to be overexpressed under pathological conditions, and Marceau et al. 6 found this receptor to be upregulated in damaged tissues. In addition, cardioprotective biomarkers, such as the angiotensin 2 (AT2) receptor, have been associated with the kinin pathway7,8 through the promotion of NO release, although it has not been studied during the evolution of infarction in females. Burrell et al. 9 instead found an increase in angiotensin-converting enzyme 2 (ACE2) mRNA after three and 28 days of coronary occlusion in male rats. On the other hand, the beta-2 (B2) receptor has also been considered a cardioprotective biomarker.10,11 B2 receptor mRNA is decreased in kidney tissues after an ovariectomy, 12 an effect that was reversed by estradiol administration, though no changes were found in the heart and uterus. On the other hand chronic hormone replacement therapy alters the expression of the AT1 receptor on rat anterior pituitary. 13 In the same manner, it was found that hormonal replacement increased AT1 receptor expression by 180% in the uterus. 14 Other researchers have found an increase in AT1 and AT2 receptor expression in the pituitary gland with estrogenic treatment. However, it has been reported that there is a decrease in the expression of ACE and AT1 receptors in vascular smooth muscle cells 15 and adrenal cortex. 16 These findings indicate that estrogens modify the expression of RAS components depending on the tissue in ovarectomized rats with estrogen replacement therapy. Therefore it is important to know mRNA expression changes of some RAS and KKS components in cardiac tissue in both sexes during acute and chronic MI.
Methods
Animals
Thirty-six male (300–400 g) and 36 female Wistar rats (250–350 g) were obtained from Cinvestav’s Southern Unit. All animals were placed in individual boxes under light-dark cycles of 12 h with food and water ad libitum. Female rats were not subjected to ovarectomy and grouped by coronary occlusion (CO) time. Six groups of each sex were formed (n=6): (a) sham group; (b) 48 h CO; (c) one week CO; (d) two weeks CO; (e) three weeks CO; (f) four weeks CO. Sham animals were sacrificed one week after simulated occlusion. The experimental protocols were carried out in compliance with the Mexican Federal Regulation for Animal Experimentation and Care.
Reagents
Primers for the receptors AT1, AT2, B1, and B2 and the enzymes ACE and ACE2 were purchased from Sigma-Aldrich Inc. (St Louis, Missouri, USA). RNA was obtained using AxyPrep Multisource Total RNA Miniprep Kit (Axygen, Union City, California, USA), real-time quantitative polymerase chain reaction (RT-qPCR) was performed using the Bio-Rad Reverse Transcription Supermix Kit (Hercules, California, USA) and a Roche Lightcycler Kit 480 DNA SYBR Green Master Mix (10×) for amplification.
MI induction
The left anterior descending coronary artery was permanently occluded in all groups, except for the sham group, following the technique previously standardized by our group. 17
Real-time polymerase chain reaction (RT-PCR) analysis
Analysis was performed to determine the expression levels of the B1, B2, AT1, and AT2 receptors and of ACE and ACE2 in the left ventricle. Total RNA was isolated using a commercial kit (AxyPrep Multisource Total RNA Miniprep; Axygen, Union City, California, USA), and the mRNA was then reverse-transcribed using a kit (Bio-Rad Reverse Transcription Supermix Kit). The negative controls for the RT-PCR, including a reaction with master mix but without ventricle segments, and a reaction in the absence of reverse transcriptase, were performed in parallel. RT-PCR was performed (Roche LightCycler 480 DNA SYBR Green Master Mix) with 10 μmol/l of the sense and antisense primers for the cDNA of interest. Sense and antisense primers were used as shown in Table 1. PCR was routinely performed for 45 cycles, with a hot start at 95°C/5 min, followed by cycles at 95°C/10 s, annealing temperature (Tm, °C)/10 s and 72°C/10 s, and an extension at 72°C/10 min. The relative expression of mRNA was analyzed using the −2ΔΔCt method.

Statistical analysis
All data are expressed as the mean±standard error of the mean (SEM). The data were analyzed using two-way analysis of variance (ANOVA) and Student-Newman-Keuls post-hoc analysis. Values of p<0.05 indicate statistical significance.
Results
The infarction area between males and females was similar over time, and the area was measured from the outer wall of the left ventricle (Table 2). Table 3 shows the results the changes in the mRNA expression of the B1, B2, AT1, and AT2 receptors, as well as of the ACE and ACE2 enzymes, on the penumbra of the left ventricle during the evolution of four weeks of myocardial infarction in male and female Wistar rats. It was observed at 48 h of coronary occlusion that there was a significant increase in the mRNA of these proteins in both males and females. Furthermore, in the males there was an increase in the mRNA of biomarkers after three weeks of coronary occlusion, whereas in the females this increase occurred after 48 h and two weeks of coronary occlusion. Alternately, the females overexpressed the mRNA of these six biomarkers to a lesser degree than the males. Finally, it was observed that the overexpression of the B2 receptor mRNA occurred at all occlusion times in the females, but in males this receptor was overexpressed only at 48 h and three weeks. Tables 4 and 5 summarize the influence of sex and evolution time of MI on the mRNA expression of SRA and KKS components; mRNA is time dependent at 48 h, one, three, and four weeks of CO, while at two weeks the expression is dependent on sex.
Infarcted size area and relationship of left ventricle weight (LVW) with respect to body weight (BW) of male and female Wistar rats.
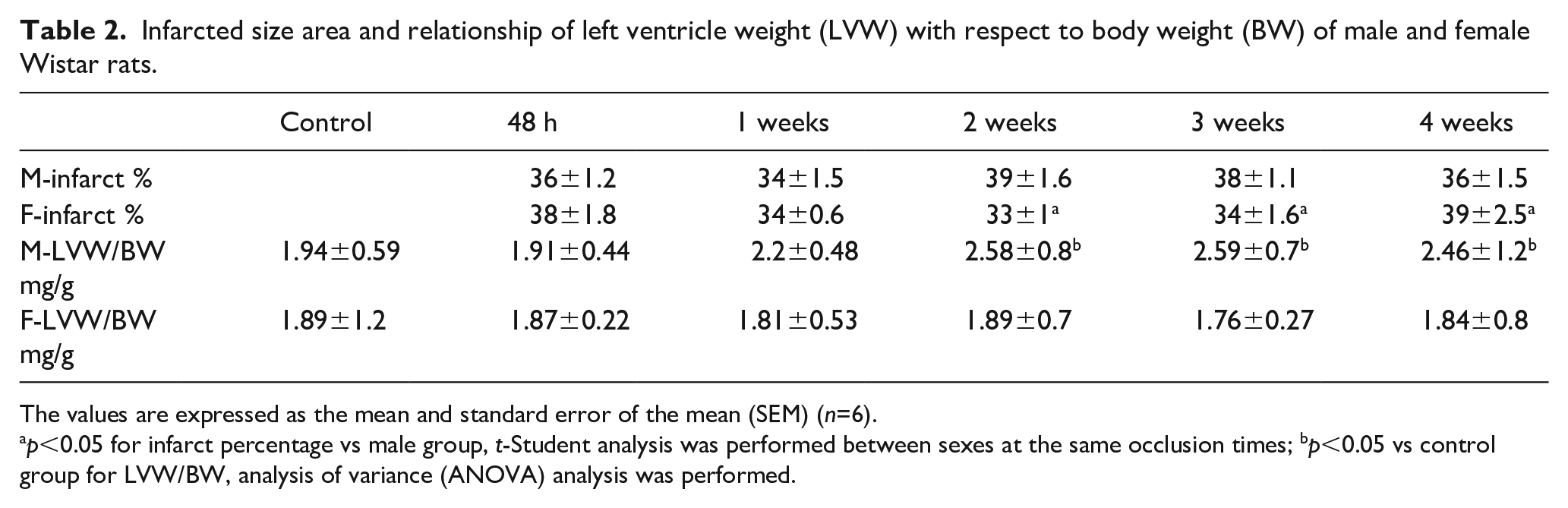
The values are expressed as the mean and standard error of the mean (SEM) (n=6).
p<0.05 for infarct percentage vs male group, t-Student analysis was performed between sexes at the same occlusion times; bp<0.05 vs control group for LVW/BW, analysis of variance (ANOVA) analysis was performed.
mRNA expression of renin-angiotensin system (RAS) and kallikrein-kinin system (KKS) biomarkers in samples from left ventricular penumbra after 48 h, one, two, three, and four weeks (w) of coronary occlusion of male (M) and female (F) Wistar rats.

ACE: angiotensin-converting enzyme; AT1: angiotensin 1; AT2: angiotensin 2; B1: beta-1; B2: beta-2.
The values are expressed as the mean and standard error of the mean (SEM) (n=6).Two-way analysis of variance (ANOVA) was performed to compare experimental groups between sex and time of coronary occlusion.
bp<0.05 vs control group, ap<0.05 vs female.
Changes on the left ventricular mRNA of detrimental biomarkers on male (M) and female (F) Wistar rats during myocardial infarction (MI) evolution.

ACE: angiotensin-converting enzyme; AT1: angiotensin 1; AT2: angiotensin 2; B1: beta-1; B2: beta-2; w: week.
Changes on the left ventricular mRNA of cardioprotective biomarkers on male (M) and female (F) Wistar rats during myocardial infarction (MI) evolution.

ACE: angiotensin-converting enzyme; AT1: angiotensin 1; AT2: angiotensin 2; B1: beta-1; B2: beta-2; w: week.
Discussion
This article aims to evaluate changes in mRNA expression of components of the renin-angiotensin system (ACE, ACE2, AT1, and AT2) and the Kallikrein-Kinin system (B1 and B2) in the left ventricle of male and female Wistar rats at 48 h, one, two, three, and four weeks after coronary occlusion to identify gender differences. Coronary occlusion times of this study are due to a previous study by our team 17 where the differences between sex of vascular reactivity to AT2 were monitored at 2,4, 24, 48 h and one, two, three, and four weeks post-infarction where we found female reactivity returns to normal after one week of coronary occlusion while the reactivity of male increases in a time-dependent manner. Our results were attributed to female compensatory mechanisms that are related to the RAS and the KKS. As we know AT2 plays a critical role in the evolution of MI by an AT1 receptor increase, which generates hypertrophy and left ventricular dysfunction. 22 This reasoning is why it is important to study the changes in mRNA expression of this receptor. Milik et al. and Zhu et al.23,24 have reported the increase in mRNA of AT1 receptors after MI and, in agreement with our study, AT1 mRNA expression was upregulated at three weeks in males and at 48 h and two weeks in female rats. Relative to ACE, this is a key modulator of MI 25 through a detrimental effect. Passier et al. 26 found an increase in ACE mRNA expression at four, seven, and 90 days. While Zhu et al. 27 observed that ACE mRNA increased at three weeks after coronary artery ligation in male rats. This is in agreement with our results, which showed an increase at seven days and three weeks in male rats. In addition, ACE mRNA expression in female rats was increased at two weeks of CO, which has not been previously reported. In relation to kinins, Zhu et al. 28 showed that these substances were released from the myocardium during ischemia. Even though the studies of Chahine and Tschöpe29,30 showed that the B1 receptor has a cardioprotective effect, others disagree, such as Potier et al. 31 who described how the B1 receptor role in MI remains uncertain. Tschöpe et al. 32 had previously reported an upregulation of B1 receptor expression during the acute phase of MI in male rats, while we found the same increase in both sexes, in the chronic phase at three and two weeks in male and female rats, respectively. We also found that the increase in the expression of the B1 receptor and ACE were present at the same CO times in both sexes, which could be related to the results of Ignjacev et al. 33 who reported that ACE can upregulate B1 and B2 receptor expression. In addition, the RAS and KKS have regulatory biomarkers that counteract the effects of AT1/ACE/B1 as AT2 receptors, which have antiproliferative effects.34,35 During MI, Nio et al. reported an increase in mRNA of the AT2 receptor at 24 h and seven days after CO in male rats. 36 In contrast to their results, we found a decrease at the same times in males, while in females the decrease was at seven days. There are no reports on the mRNA expression of the AT2 receptor during the chronic phase of MI, but our results show an increase in both sexes, which could be beneficial. Some reports have shown differences in this receptor expression due to sex, such as Hilliard et al., who reported that the expression of AT2 is higher in females than in males, 36 modulating blood pressure by natriuresis through a vascular effect. In addition, Okumura et al. 37 reported that the decrease in AT2 receptor mRNA may be involved in the estrogen response improving vascular remodeling depending on age. Another cardioprotective biomarker is ACE2, with this study showing an overexpression of this enzyme at the acute and chronic phase for both sexes, in accordance with Burrell et al. and Yamamoto et al., who demonstrated that ACE and ACE2 mRNA expression increased during the acute and chronic phases9,38 and speculated that this increase might counteract the effect of mechanical injury and cardiac remodeling. In addition, Grobe et al. 39 proposed that ACE2 expression and activity increased after MI, exerting a cardioprotective effect due to an increase in the levels of angiotensin (Ang) (1–7). Referring to the B2 receptor, we found an increase in the mRNA during the acute and chronic phases of MI in male and female rats. This receptor has cardioprotective effects during the acute phase of MI, and its overexpression was related to an improvement in left ventricular function by reducing inflammation and cardiac remodeling.40–42 On the other hand estrogens through their genomic pathways activate transcription factors, which promote the synthesis of endothelial NOS and superoxide dismutase of manganese resulting in a greater production of nitric oxide in the vasculature, 43 promoting vasodilation and a decrease in superoxide anion levels reducing ROS so, in the absence of estrogen there is an uncoupling of respiratory increasing oxidative stress in the mitochondria, these ROS are released into the cell through the mitochondrial transition pore which remains open, promoting apoptosis and cellular necrosis. 44 On the other hand, it is known that estrogens regulate gene expression through genomic pathways, among these the modulation has been studied of the AT1 receptor and ACE that are related to the increase in blood pressure, calcium overload, and cardiac remodeling. 45 However, in this study only the mRNA expression of different components of the RAS and KKS was determined, so it is necessary to quantify the protein of each one at two and four weeks to be able to relate them as damage or protection biomarkers.
Conclusion
The expression of RAS and KKS biomarkers occurs at earlier times in the female compared to the male rat, in addition during chronic MI these biomarkers remain unchanged in females while they decreased in males. Due to the results obtained herein, we should focus on the study of sex-dependent phases to identify the physiopathological and molecular changes related to hormones, receptors, and enzymes and to determine the differences between males and females. Such studies will provide more information and adequate prognosis, diagnosis, and pharmacological treatment for females.
Limitations
Since gene expression is dependent on each species, it is possible that the temporal expression of the genes analyzed is not the same in humans as in rats. By determining the expression of several RAS and KKS markers, it is not ensured that the quantification and activity of the protein is the same as the mRNA of each component. On the other hand, it should be considered that a sample (n=6) was taken for each group, so to ensure that the expression of these genes is regulated in the rat according to our data a larger population sample should be taken.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from PAPIIT IN213318, PAPIIT IA205119 DGAPA UNAM, PIAPIME ID 2.17.02.18 and PIAPI 1828 FES Cuautitlan, Universidad Nacional Autonoma de México. CONACYT A1-S-8958.