
Abstract
The aim of this study was to investigate the protective effects of sulfur dioxide (SO2) on the endothelial function of the aorta in D-galactose (D-gal)-induced aging rats. Sprague Dawley rats were randomized into a D-gal group, a D-gal + SO2 group and a control group, then injected with D-gal, D-gal + SO2 donor or equivalent volumes of saline, respectively, for 8 consecutive weeks. After 8 weeks, the mean arterial pressure was significantly increased in the D-gal group, but was lowered by SO2. SO2 significantly ameliorated the endothelial dysfunction induced by D-gal treatment. The vasorelaxant effect of SO2 was associated with the elevated nitric oxide levels and upregulated phosphorylation of endothelial nitric oxide synthase. In the D-gal group, the concentration of angiotensin II in the plasma was significantly increased, but was decreased by SO2. Moreover, levels of vascular tissue hydrogen peroxide (H2O2) and malondialdehyde were significantly lower in SO2-treated groups than those in the D-gal group. Western blot analysis showed that the expressions of oxidative stress-related proteins (the angiotensin II type 1 receptor (AT1R), and nicotinamide adenine dinucleotide phosphate oxidase subunits) were increased in the D-gal group, while they were decreased after treatment with SO2. In conclusion, SO2 attenuated endothelial dysfunction in association with the inhibition of oxidative stress injury and the downregulation of the angiotensin II/AT1R pathway in D-gal-induced aging rats.
Introduction
Aging is a complex, inevitable biological process induced by a wide variety of genetic, temporal, environmental and other factors. It is one of the primary risk factors for the progression of cardiovascular diseases such as hypertension and atherosclerotic vascular disease, independently from traditional risk factors. 1 With aging, the arterial endothelia, vascular walls and adventitia undergo structural, mechanical and functional changes that induce vascular stiffening and increased vascular tone; these changes are characterized by endothelial dysfunction, 2 thickening of the vascular wall 3 and remodeling of the adventitial extracellular matrix, 4 respectively. Endothelial dysfunction has been widely recognized as a key contributor to this process of vascular aging. 5
Multiple mechanisms are involved in aging-related endothelial dysfunction, including reduced nitric oxide (NO) dilator activity, increased reactive oxygen species (ROS), the stimulation of proinflammatory cytokines, and accelerated endothelial cell senescence; however, most important is the activation of the renin-angiotensin system (RAS), which plays a central role in the regulation of endothelial function.6–9 Increased RAS activity has been widely described in association with the onset and progression of vascular aging, and RAS blockade has been shown to protect against aging-related deleterious effects and to promote longevity.10–11 Although the molecular mechanisms of vascular aging have been well investigated, effective therapeutics are extremely scarce and interventions are limited to the improvement of lifestyle habits, such as engaging in moderate exercise and limiting caloric intake.12–13 With the continuous increase in the proportion of the population who are elderly, there is a great need for interventions in vascular aging.
Sulfur dioxide (SO2) has traditionally been thought of an industrial waste gas that can cause respiratory and cardiovascular diseases with chronic exposure. 14 However, in recent years, increasing evidence suggests that SO2 could not only be endogenously generated from the oxidation of sulfur-containing amino acids, such as cysteine and homocysteine, catalyzed by aspartate aminotransferase (AAT) in the cardiovascular system, 15 but may also play an important physiological role in maintaining homeostasis, including regulating heart rate, 16 vasodilation 17 and negative regulation of cardiac function. 18 An increasing number of studies have shown that endogenous SO2 also participates in many pathophysiological processes related to vascular diseases. It can attenuate increased blood pressure and vascular remodeling in spontaneously hypertensive 19 and hypoxic pulmonary hypertensive rats, 20 and it significantly ameliorates vascular calcification in association with downregulation of the TGF-β/Smad pathway. 21 Studies have also found that the downregulation of the SO2/AAT pathway is involved in the pathogenesis of atherosclerosis. 22 In addition, a recent study reported that SO2 attenuates vascular smooth muscle cell proliferation in angiotensin II (Ang II)-induced hypertensive mice. 23 However, although many studies have revealed various cardiovascular protective effects of SO2, its action on vascular aging, especially on aging-related endothelial dysfunction, has not been fully investigated, and the underlying mechanisms remain to be established.
With this in mind, the aim of this study was to investigate a possible role of SO2 in aging-related endothelial dysfunction by using an animal model of aging induced by D-galactose (D-gal). Moreover, we attempted to explore whether SO2 exerts its protective effects via downregulating of the Ang II/Ang II type 1 receptor (AT1R) pathway.
Materials and methods
Animals and experimental protocol
Eight-week-old male Sprague Dawley rats were purchased from Vital River Laboratories and housed under constant environmental conditions (12-hour light/dark cycle) in a temperature controlled (22 ± 1°C) facility with free access to water and food. All animal experimental procedures were performed according to the Guide for the Care and Use of Laboratory Animals of the US National Institutes of Health (NIH) (publication number 85-23, revised 1996).
A total of 24 rats were randomly divided into three groups with eight rats in each group: control, D-gal, and D-gal + SO2. The rats in the D-gal and D-gal + SO2 groups were injected subcutaneously daily with D-gal (150 mg/kg) dissolved in saline for 8 consecutive weeks, and the control group rats were administered the equivalent volume of saline. In the D-gal + SO2 group, sodium sulfite (Na2SO3) and sodium bisulfite (NaHSO3) were used as the SO2 donors (molar ratio of Na2SO3 and NaHSO3 was adjusted to 3:1, pH 7.4), and were administered intraperitoneally at a dose of 85 mg/kg daily for 8 consecutive weeks. The control and D-gal group rats were intraperitoneally administered with the equivalent volume of saline at the same time points.
Measurement of blood pressure
After eight weeks, mean arterial pressure (MAP) was measured noninvasively by tail-cuff plethysmography (BP-100A, Chengdu, Sichuan, China) after the rats were stabilized and remained quiescent. The value was taken as the average of at least three measurements after removing the outliers and any readings associated with excess noise or animal movement on each occasion.
Rat aorta preparation and isometric force studies
After 8 weeks, the rats were sacrificed by CO2 suffocation, and the thoracic aorta was dissected, cleaned of connective tissue and cut into ring segments (3 mm lengths). Then vasodilatation function was assessed in these isolated intact thoracic aortic rings, as previously described. 24 Briefly, an optional baseline tone of 2 g was applied to all rings as a resting tension. After 60 min equilibration, arteries were precontracted with phenylephrine (1 μmol/L). When the steady contraction was achieved, acetylcholine (ACh, 10−8~10−4 mol/L) or sodium nitroprusside (SNP, 10−9~10−6 mol/L) was added in a cumulative manner to record the endothelial-dependent relaxation or endothelial-independent relaxation response curve. The extent of relaxation was expressed as the percentage of the relaxation arising from a submaximal phenylephrine-induced constriction.
Measurement of plasma Ang II level
Radioimmunoassay was used to measure plasma Ang II concentration. Blood samples were collected and centrifuged at 3000 r/min for 15 minutes at 4°C. The plasma was collected and detected with an Ang II radioimmunoassay kit (China Institute of Atomic Energy, Beijing, China), according to the manufacturer’s instructions.
Measurement of ROS and antioxidant enzyme activities
The thoracic aortas were dissected and snap-frozen at −80°C. After homogenization with 50 mmol/L potassium phosphate buffer and centrifugation at 10,000 × g for 5 min, hydrogen peroxide (H2O2) and malondialdehyde (MDA) concentrations, as well as superoxide dismutase (SOD) activities, in the supernatant were measured with the corresponding assay kits (Jiancheng Bioengineering Institute, China), according to the manufacturer’s instructions. These indicators were all standardized by protein content, determined using a bicinchonininc acid (BCA) protein assay kit.
Measurement of plasma NO concentration
Plasma NO concentrations were determined with commercially available kits (Jiancheng Biomedical Engineering, China), according to the manufacturer’s instructions.
Western blot analysis
Protein extracted from the thoracic aorta was quantified using BCA reagent and protein samples (50 μg/lane) were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis gels, transferred to polyvinylidene fluoride membranes and blocked with 5% nonfat milk for 1 hour. The membranes were then incubated overnight at 4°C with primary antibodies: Nox2, Nox4, AT1R, SOD1, endothelial nitric oxide synthase (eNOS) (1:1000, Abcam, USA) and phosphorylated eNOS (p-eNOS) (Ser1177, 1:1000, Abcam, USA), with glyceraldehyde 3-phosphate dehydrogenase as the internal control (1:2000, Proteintech, USA). After washing with Tris-buffered saline with Tween three times, the membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies at room temperature for 1 hour. Target bands were detected with SuperSignal West Pico Chemiluminescent Substrate (Thermo Scientific-Pierce, USA). The band intensity was quantified using Image J software.
Statistical analyses
Results were expressed as means ± SEMs. Statistical analysis was performed using an SPSS software package, version 17.0 (SPSS, Inc., Chicago, IL, USA). The results for three or more groups were compared using one-way analysis of variance followed by a Student–Newman–Keuls test. Comparisons between two groups were made using the Students’ t-test. p < 0.05 was considered significant.
Results
SO2 attenuated the impairment in endothelial function in D-gal-induced aging rats
After 8 weeks of treatment, there was a significant increase in MAP (Figure 1(a)) in the D-gal group (99.38 ± 2.71 mmHg) compared with the control group (81.13 ± 2.70 mmHg, p < 0.01), but 8 weeks of SO2 treatment significantly decrease the MAP in D-gal-induced aging rats (89.25 ± 3.03 mmHg, p < 0.05).
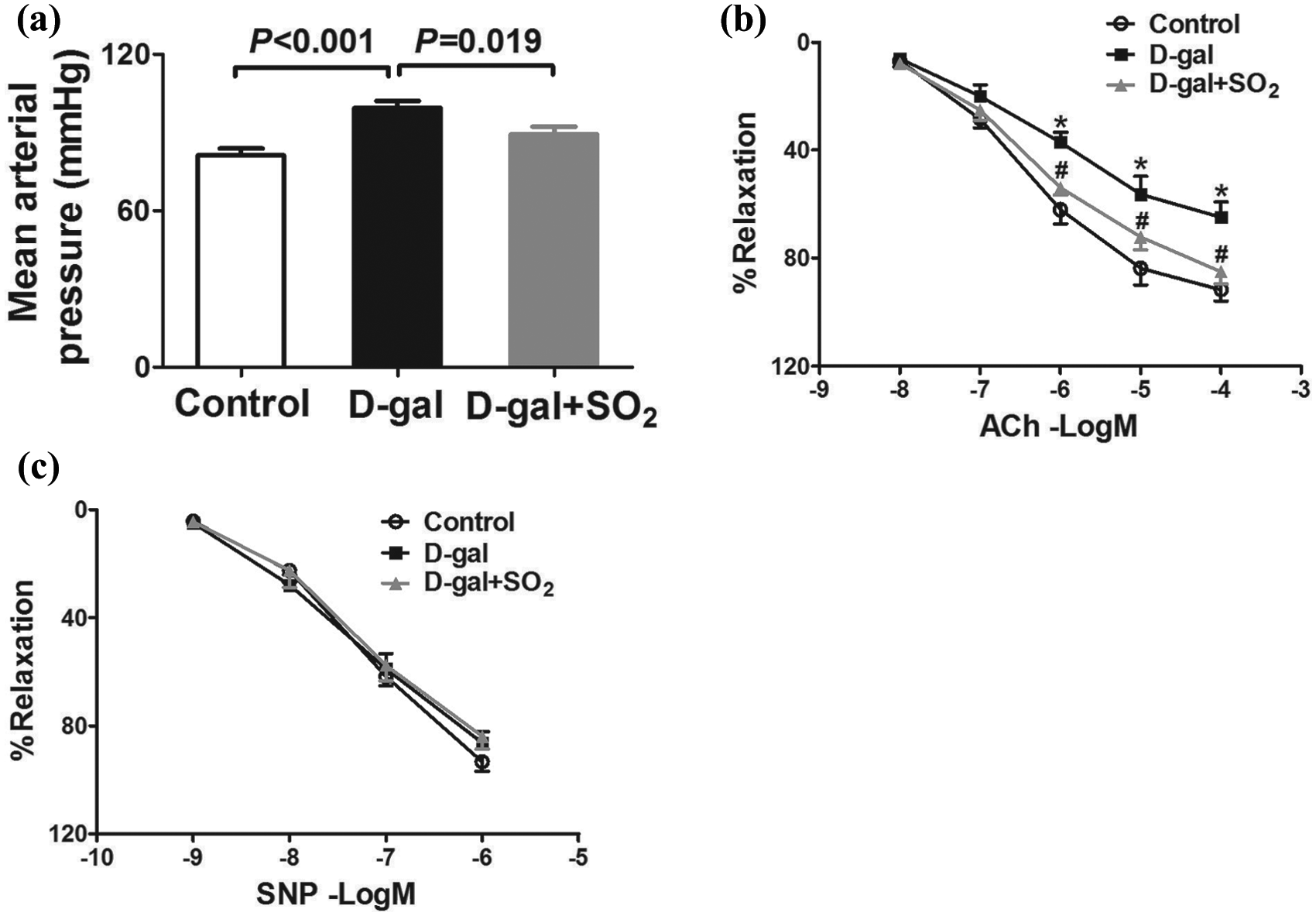
Sulfur dioxide attenuated the impairment in endothelial function in D-galactose-induced aging rats.
Isometric studies showed impairment of ACh-induced relaxation in the aortas of the D-gal group rats (Emax 64.87 ± 5.67 % vs 91.75 ± 4.19 % in the control group, p < 0.05). Treatment with SO2 for 8 weeks markedly improved ACh-induced endothelium-dependent relaxation (85.02 ± 4.58 %, Figure 1(b)). In contrast, SNP-induced endothelium-independent relaxation showed no difference between the groups (Figure 1(c)).
SO2 upregulated the eNOS/NO pathway in D-gal-induced aging rats
As shown in Figure 2(a), the plasma NO concentration was significantly lower in the D-gal group rats than those in the control group (22.66 ± 2.42 μmol/L vs 38.74 ± 2.98 μmol/L, p < 0.01). Meanwhile, p-eNOS at the activation site of Ser1177 was also decreased in the aortas of the D-gal group rats (p < 0.01) (Figure 2(b)). Eight weeks of SO2 treatment significantly reversed the downregulation of p-eNOS (p < 0.05) and increased the plasma NO concentration (p < 0.05)
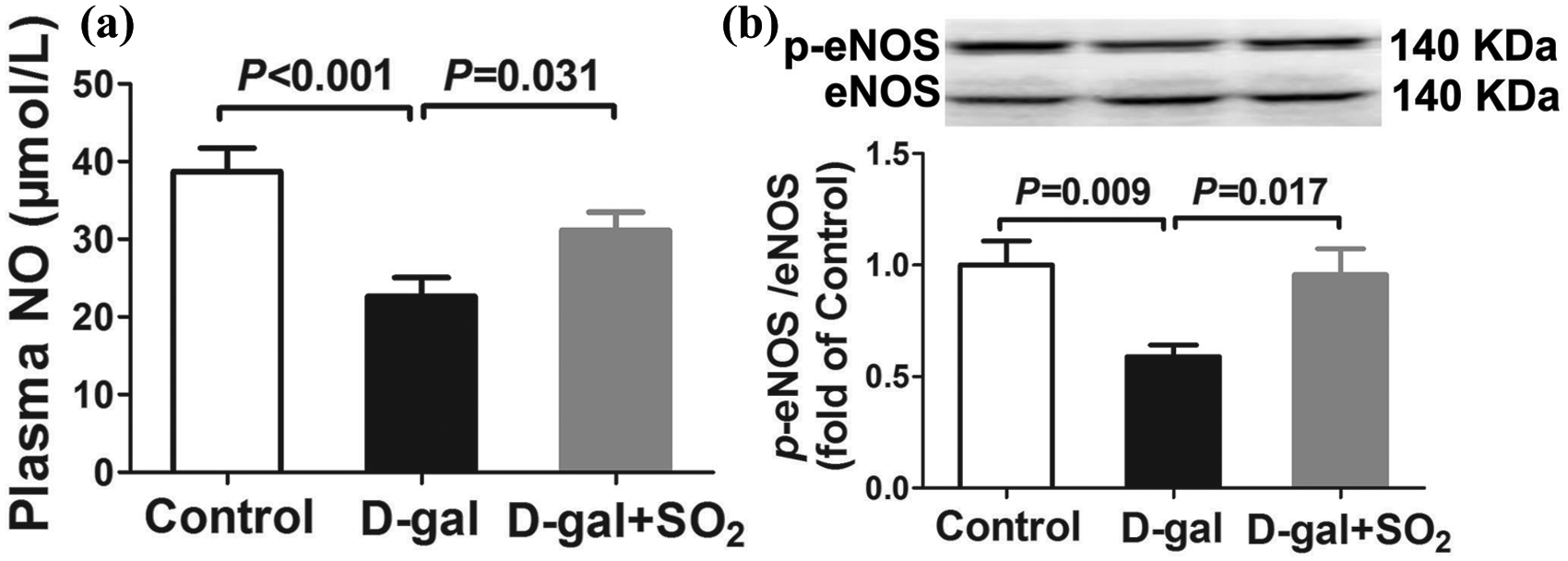
Sulfur dioxide upregulated the endothelial nitric oxide synthase/nitric oxide pathway in D-galactose-induced aging rats.
SO2 resists oxidative stress in D-gal-induced aging rats
There was a significant increase in oxidative stress reflected by the increased levels of H2O2 and MDA in the aortas of the D-gal group rats compared with the control group (both p < 0.01), but these increases were markedly attenuated in the D-gal + SO2 group (both p < 0.05), indicating that SO2 prevented oxidative stress injury in D-gal-induced aging rats (Figure 3(a) and (b)).

Sulfur dioxide resisted oxidative stress in D-galactose-induced aging rats.
Concomitant with the increased H2O2 and MDA levels, there was a significant decrease in the activity of the antioxidant enzyme SOD in the D-gal group compared with the control group (19.44 ± 2.17 U/mg protein vs 29.64 ± 5.30 U/mg protein, p < 0.05), while treatment with SO2 did not change the activity and protein expression of SOD in the aortas of D-gal-induced aging rats (Figure 3(c) and (e)).
However, western blot analysis showed that the protein level of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase subunits Nox2 and Nox4 were elevated in the thoracic aortas of the D-gal group rats, but that this increase was reduced in the D-gal + SO2 group (Figure 3(f) and (g)).
SO2 inhibited the Ang II/AT1R pathway in D-gal-induced aging rats
As is shown in Figure 4(a), the plasma concentration of Ang II was significantly increased in the D-gal group compared with the control group (124.49 ± 8.84 pg/mL vs 86.50 ± 7.53 pg/mL, p < 0.01). However, treatment with SO2 for 8 weeks lowered the Ang II level to 99.79 ± 5.88 pg/mL (p < 0.05).

Sulfur dioxide inhibited the Ang II/AT1R pathway in D-galactose-induced aging rats.
Western blot analysis (Figure 4(b)) was used to determine AT1R protein expression in the thoracic aorta in the three groups of animals. The results showed that the protein levels of AT1R were elevated in the D-gal group rats compared with the control group (p < 0.01), and that this overexpression was reversed in the D-gal + SO2 group (p < 0.05).
Discussion
The two main findings of the present study were that: (a) SO2 improved the endothelium-dependent relaxation of the aging aortas and (b) that SO2 downregulated the Ang II/AT1R pathway and oxidative stress in a rat model of aging induced by D-gal. Taken together, the results suggest that SO2 could improve aging-related endothelial dysfunction by inhibiting oxidative stress injury by downregulating the Ang II /AT1R pathway in D-gal-induced aging rats. Moreover, the findings suggest a potential role for SO2 in clinical interventions to reverse the adverse Ang II profile in aged populations.
Advancing age is a complex and irreversible process in which a progressive loss of physiological integrity results in functional impairment and decreased quality of life. Age-related alteration of the structure and function of the vasculature is a dominant risk factor for cardiovascular diseases, the leading cause of death worldwide. Given the increasing elderly population, interventions to combat vascular aging could not only improve the health of the elderly but might also extend their life span by delaying cardiovascular disease-related deaths.
Several animal models have been used to gain a better understanding of aging and to help establish methods to prevent its effects on various organs. Amongst these, the D-gal-induced aging model is the most common model for mimetic aging.25–27 D-gal, a reducing sugar normally present in the body, can be oxidized by galactose oxidase to form H2O2, leading to oxidative stress, inflammation and apoptosis at high levels. 28 Evidence has shown that abnormal D-gal metabolism can cause aging-related changes, and that D-gal-induced aging rodents exhibit features that are in many ways markedly similar to those of rodents that have aged naturally. 29 Therefore, the chronic injection of D-gal has been regarded as an ideal model for studying the possible mechanisms of aging-related diseases, especially for vascular aging. 30
In the current study, chronic systemic D-gal exposure for 8 weeks resulted in impaired endothelium-dependent vasorelaxation with an associated increase in blood pressure, but there were no changes in endothelium-independent relaxation. Our data were consistent with several previous studies on vascular aging, which showed a linear rise in systolic blood pressure induced by arterial stiffness and a concurrent increase in diastolic blood pressure resulting from endothelial dysfunction-induced high peripheral vascular resistance with advancing age. 31 Long-term treatment with SO2 can notably ameliorate all the D-gal-induced changes. SO2 has historically been regarded as a toxic gas and environmental pollutant. However, research in recent years has demonstrated that it is endogenously synthesized during the oxidation of sulfur-containing amino acids by AAT in mammalian tissues. Endogenous SO2 is now regarded to be the fourth gasotransmitter after NO, carbon monoxide and hydrogen sulfide, and it performs various important regulatory functions that maintain homeostasis.32–34 Increasing numbers of studies have shown that SO2 participates in pathophysiological processes related to various vascular diseases. In spontaneously hypertensive and pulmonary hypertensive rats, the SO2/AAT pathway is significantly downregulated, accompanied by an increase in blood pressure; however, treatment with SO2 can attenuate the increase in blood pressure through its vasorelaxant effect and can inhibit vascular remodeling by suppressing the proliferation of vascular smooth muscle cells, promoting apoptosis and inhibiting the synthesis of extracellular collagen while promoting its degradation.35–37 The findings of the present study clearly demonstrate that SO2 can, to some extent, improve endothelial dysfunction, one of the major vascular changes caused by D-gal-induced aging; however, the specific molecular mechanisms for this remain to be established.
Under healthy conditions, the endothelium plays a pivotal role in maintaining vascular homeostasis, attributed to the balance between the production of endothelium-derived relaxing factors and contracting factors. NO produced by eNOS is a major endothelium-derived relaxing factor, and it has been proposed that the decreased bioavailability or synthesis of NO is a common molecular mechanism underlying predisposition to aging-dependent disorders, including endothelial dysfunction. 38 Thus, new strategies to restore NO bioavailability may have therapeutic benefit. Our results showed that the plasma NO concentration decreased after treatment with D-gal alone, but that this decrease was significantly attenuated when SO2 was administered concomitantly with the D-gal. This indicated that SO2 could improve D-gal-induced endothelial dysfunction by increasing NO concentrations. Similarly, Lu et al. observed that SO2 reduced blood pressure and increased vasorelaxation in spontaneously hypertensive rat arteries by increasing the NO level in the aortic tissues. 19 In the endothelial cells, NO is endogenously produced from L-arginine by eNOS, one of three NOSs, with the formation of L-citrulline. A further finding of the present study was that the phosphorylation of eNOS at the activation site of Ser1177 decreased in D-gal-induced aging rat aortas and that SO2 treatment significantly reversed the downregulation of p-eNOS. These results are consistent with the study by Li et al., 39 who reported that SO2 could upregulate the eNOS-NO-cyclic guanosine monophosphate pathway, resulting in vasodilation. They also found that SO2 could increase the NO/eNOS pathway and significantly reduce atherosclerotic lesions. 22 Conversely, the excessive generation of ROS can impair NO availability. Oxidative stress induced by increased oxygen-derived free radicals is widely acknowledged to be an important factor associated with the aging process. Excess superoxide (O2-) can react with NO to produce peroxynitrite (ONOO-), a powerful oxidant that impairs endothelial cell function and results in vascular aging. 40 This reaction can reduce NO bioavailability, which then increases vascular tone and blood pressure. We therefore investigated the antioxidant effects of SO2 in D-gal-induced aging rats. Our results suggested that SO2 could reduce the levels of two final products of lipid oxidation, MDA and H2O2, to reduce the oxidative stress injuries induced by the presence of D-gal. As a reductant, the antioxidant effects of SO2 have been confirmed by a growing number of articles,41–42 although the detailed mechanisms remain unclear.
ROS are oxygen-containing chemically reactive molecules. The mitochondria, endoplasmic reticulum and NADPH oxidase system are the major sites for the generation of ROS, which are produced by cellular respiration, metabolism and phagocytosis. O2- generated by NADPH oxidase is converted by SOD to H2O2, which can then be converted either to H2O by catalase or to hydroxyl radicals in the presence Fe2+. Further, H2O2 can also be reduced by antioxidants such as glutathione peroxidase. 43 An imbalance between prooxidant stimuli and antioxidant defenses can result in cellular damage referred to as oxidative stress. Jin et al. 44 reported that SO2 preconditioning increased plasma SOD, GSH and GSH-Px levels, and myocardial SOD1 protein expression, in rat with myocardial ischemia-reperfusion injury. We attempted to verify whether SO2 increased the antioxidative capacity of the aortas of D-gal-induced aging rats by upregulating the expression or increasing the activity of SOD1. We found that treatment with SO2 did not affect the activity and protein expression of SOD1, which were reduced in both groups of rats treated with D-gal. This implied that the inhibition of ROS by SO2 takes place via a different pathway, with further study needed.
Several studies have reported that the mitochondrial and membrane NADPH-oxidase complexes are the major enzymatic sources of ROS production in vascular cells. Nox proteins, the catalytic subunits of NADPH oxidase, produce high levels of extracellular O2- by transferring an electron to oxygen. Nox upregulation has been implicated in the oxidative stress that is related to the pathogenesis of several age-related diseases such as hypertension, diabetes and atherosclerosis. Nox isoforms include Nox1, Nox2, Nox4 and Nox5, of which Nox2 and Nox4 are the most important isoforms in vascular cells. 45 In the present study, we measured the expression of Nox2 and Nox4 and found that treatment with D-gal alone significantly increased the protein expression levels of Nox2 and Nox4 in aortic tissue, but that concomitant treatment with SO2 normalized the expression levels. There is a large body of evidence in support of chronic elevation of Ang II activating intracellular NADPH oxidase via AT1R to generate O2- during the process of aging.46–47
The RAS is widely recognized as a key factor contributing to the pathophysiological processes of various aging-related disorders. Ang II is a key regulator of cell senescence, and the activation of AT1R signaling is a form of stimulation that promotes aging. Increased Ang II levels and markedly upregulated AT1R can be found in the aged arterial wall.48–49 However, several studies have shown that chronic treatment with ACE inhibitors and AT1R antagonists can prevent the ultra- and microstructural age-related changes and extend life span.50–51 Consistent with this, mice null for AT1R show increased longevity. 52 Furthermore, AT1R deletion results in a decrease in the age-related progression of atherosclerosis. 53 Consistent with the previous observation, we found that the plasma Ang II levels and the expression of AT1R protein in the vasculature were elevated in D-gal-induced aging rats as compared with those in the control group, but that treatment with SO2 could suppress the increased activity of the RAS. Chen et al. have reported that SO2 prevented Ang II-induced myocardial hypertrophy accompanied by downregulating cardiomyocyte autophagy. 54 SO2 also can inhibit extracellular signal-regulated kinase signaling to attenuate vascular smooth muscle cell proliferation in Ang II-induced hypertensive mice. 23
The present study had several limitations. Importantly, we did not test the endogenous SO2 levels in either the tissue or the plasma. Whether changes in endogenous SO2 levels cause or are the consequence of the pathological state is not well understood. Further studies are therefore needed to explore the roles of endogenous SO2 in the pathogenesis of D-gal-induced aging. In addition, if SO2 is to be used for clinical treatment, it is important its highly toxic actions at supraphysiological levels are fully considered, and great care should be taken during the development of SO2-based therapeutic agents. Thus, there is still a long way to go.
Conclusion
In conclusion, the findings of this study suggested that SO2 could attenuate the endothelial dysfunction observed in D-gal-induced aging rats, which might be associated with the inhibition of oxidative stress, the downregulation of Ang II and the expression of AT1R. These findings provided evidence that SO2 could become a potential therapeutic approach for aging patients presenting with endothelial dysfunction.
Footnotes
Declaration of conflicting interest
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by special research funds of the Suining Central Hospital.