
Abstract
Objective:
Experimental evidence suggests that aldosterone directly contributes to organ damage by promoting cell growth, fibrosis, and inflammation. Based on these premises, this work aimed to assess the glomerular effects of aldosterone, alone and in combination with salt.
Methods:
After undergoing uninephrectomy, 75 rats were allocated to five groups: control, salt diet, aldosterone, aldosterone + salt diet, aldosterone + salt diet and eplerenone, and they were all studied for four weeks. We focused on glomerular structural, functional, and molecular changes, including slit diaphragm components, local renin–angiotensin system activation, as well as pro-oxidative and profibrotic changes.
Results:
Aldosterone significantly increased systolic blood pressure, led to glomerular hypertrophy, mesangial expansion, and it significantly increased the glomerular permeability to albumin and the albumin excretion rate, indicating the presence of glomerular damage. These effects were worsened by adding salt to aldosterone, while they were reduced by eplerenone. Aldosterone-induced glomerular damage was associated with glomerular angiotensin-converting enzyme (ACE) 2 downregulation, with ACE/ACE2 ratio increase, ANP decrease, as well as with glomerular pro-oxidative and profibrotic changes.
Conclusions:
Aldosterone damages not only the structure but also the function of the glomerulus. ACE/ACE2 upregulation, ACE2 and ANP downregulation, and pro-oxidative and profibrotic changes are possible mechanisms accounting for aldosterone-induced glomerular injury.
Introduction
Aldosterone is a mineralocorticoid hormone, whose effects are mediated by the binding to the mineralocorticoid receptors (MR), which are located in both epithelial and non-epithelial tissues.1,2 In the kidney, aldosterone acts not only on distal tubule epithelial cells, where it leads to sodium reabsorption and potassium excretion, but also on mesangial cells, podocytes, fibroblasts, and on glomerular vascular cells. 3 In recent years there has been a paradigm shift in our understanding of aldosterone actions, 3 which go far beyond sodium and potassium transport in the renal tubule and involve the ability of this hormone to modulate the local renin–angiotensin system (RAS),4,5 generate reactive oxygen species, 6 and promote fibrosis. 7
It has been shown that there is a correlation between kidney damage and circulating aldosterone,8–10 which could be due to aldosterone pro-oxidative, pro-inflammatory, and profibrotic effects, overall leading to glomerulosclerosis and tubulointerstitial fibrosis. 3 This is consistent with experimental studies showing that MR antagonists, such as spironolactone and eplerenone, protect against progressive renal injury.11–13
Although it is abundantly clear that aldosterone causes renal damage, it has not been fully elucidated yet how this hormone affects the glomerulus and whether it could damage the glomerular filtration barrier. Based on these observations, here we studied: (i) to what extent the glomerular permeability to albumin is affected by aldosterone; (ii) what are aldosterone molecular targets in isolated glomeruli (focusing on slit diaphragm components, local RAS mediators, pro-oxidative and profibrotic molecules), and (iii) whether aldosterone-induced changes are due to the binding to MR receptors.
Materials and methods
Animal model and experimental protocol
To evaluate aldosterone effects, the animal model that we chose was that of uninephrectomized, high-salt diet fed rats infused with aldosterone. 8 This is a well-established animal model to study aldosterone effects, 8 where both uninephrectomy and salt accelerate aldosterone-induced kidney damage. Several works have in fact demonstrated that salt and aldosterone synergistically contribute to renal impairment.14–16 Moreover, to discriminate the contribution of salt to glomerulosclerosis development, we randomized the rats to either salt or aldosterone, or both. Based on these premises, 75 wild-type male Wistar rats, weighing 250 g, underwent left uninephrectomy at baseline and were randomized to 5 different groups one week after. A total of 15 rats were fed with a 0.2% NaCl diet, considered the control diet (CNT), 15 rats were fed with a 1.2% NaCl diet, considered the high-salt diet (SALT), 15 rats were fed with a 0.2% NaCl diet and treated with aldosterone at a dose of 72 μg/kg/day (ALDO), 15 rats were fed with a 1.2% NaCl diet and treated with aldosterone (ALDO+SALT), and 15 rats were fed with a 1.2% NaCl diet and treated with aldosterone and its antagonist eplerenone at a dose of 100 mg/kg/day (EPL). All the rats were fed with the Harlan Teklad 18% Protein Global Diet (Harlan Laboratories, Cat#2018), which contains 0.2% NaCl, 44.2% carbohydrates, 18.6% proteins, 18.2% fibers, and 6.2% fat. The rats of the SALT and ALDO+SALT groups had also the 1.2% NaCl solution to drink. Aldosterone was infused using an Alzet 2004-osmotic minipump (Alzet 2ML4model; AzaCorp, Palo Alto, CA, USA) and its dose was chosen taking into account what has been published previously. 8 Eplerenone (Sigma, Cat#E6657) was administered incorporated into the Harlan Teklad 18% rodent diet at a concentration of 1.2 mg/g of chow (∼100 mg/kg/day). Previous works have demonstrated the stability and efficacy of this dose of eplerenone incorporated into the diet.8,14 All the rats were then followed for 4 weeks. At the end of the study, systolic blood pressure (SBP) was assessed by tail cuff plethysmography, and the rats were then placed in individual metabolic cages for 24 hours so that water intake could be recorded and total urine output collected. After performing these procedures the animals were sacrificed, and bloods and kidneys were collected for biochemical, morphological, and molecular analyses. The rats were housed at the animal house of Trieste University and were studied according to Institutional guidelines.
This study was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. A committee of the Italian Health Ministry approved the experimental protocol (permit number: prot 85 pos II/9 10/02/2006). The animals were anesthetized by intraperitoneal injection of 2,2,2-tribromoethanol (Sigma Chemical, St Louis, MO, USA), at a dose of 25 mg/100 g of body weight. Buprenorphine (Temgesic, Reckitt Benckiser) was used as analgesic and was injected subcutaneously at a dose of 0.05 mg/kg the day of the intervention and at a dose of 0.025 mg/kg the day after. All efforts were made to minimize suffering.
General parameters and biochemical data
Body weight, relative kidney mass (kidney weight/body weight), SBP, daily water intake, daily urinary volume, daily sodium excretion, AER (albumin excretion rate), serum potassium, creatinine, and aldosterone were measured at the end of the study in all the rats. Urinary sodium, serum potassium, and creatinine were assessed by autoanalyzer. Urinary AER was measured by ELISA (Bethyl Laboratories, IMTEC Diagnostics, Antwerp, Belgium). Serum aldosterone was determined by EIA (DRG diagnostics international, Marburg, Germany). Urinary nephrin was measured by ELISA (Exocell, Philadelphia, USA).
Assessment of glomerular albumin permeability
A total of 18–20 glomeruli were isolated from half kidney by standard sieving technique in media containing 50 g/L of bovine serum albumin (BSA) in PBS as an oncotic agent. Half of them were used for albumin permeability (Palb) assessment and half for RNA extraction. The glomeruli that were used for Palb assessment were incubated for 10 min at 37°C and then transferred to a glass coverslip where they were videotaped after a passage from a media containing 50 g/L of BSA to a media containing 10 g/L of BSA. This media change creates an oncotic gradient across the basement membrane, resulting in a glomerular volume change (ΔV = (Vfinal – Vinitial)/Vinitial), which we measured off-line by a video-based image analysis program. In particular, since the computer program allows to determine the average glomerular radius in a two-dimensional space, the glomerular volume can be calculated by the formula V = 4/3πr3. Then σalb, which expresses the degree of membrane permeability or leakiness, can be calculated by the formula
(σalb) = (ΔV)experimental/(ΔV)control,
and Palb, which refers to the movement of albumin subsequent to water flux, can be calculated by the formula 1 − σalb. 17
Kidney structural features
Two coronal 3 μm paraffin kidney sections were stained with hematoxylin-eosin, for tubular and glomerular area measurement, as described previously, 18 and with periodic acid-Schiff, for mesangial matrix score assessment. The amount of mesangial matrix was scored with a range from 0 to 4 (0 is for no change; 1 is for changes affecting <25%; 2 is for changes affecting 25–50%; 3 is for changes affecting 50–75%; 4 is for changes affecting >75% of the glomerulus).
Gene expression quantification by qRT-PCR
A total of 3 μg of RNA extracted from isolated glomeruli were used to synthesize cDNA with Superscript First Strand synthesis system for qRT-PCR (Gibco BRL, Grand Island, NY, USA), as previously described.19,20 Gene expression was analyzed by real-time qRT-PCR using the TaqMan system based on real-time detection of accumulated fluorescence (ABI Prism 7900 HT, Perkin-Elmer Inc, Foster City, CA, USA). In order to control for variation in the amount of cDNA available in the samples, gene expression of the target sequence was normalized in relation to the expression of an endogenous control, 18s ribosomal RNA, and then reported as arbitrary units compared to the level of expression in untreated control, which were given an arbitrary value of 1. In particular, we analyzed the glomerular gene expression of ACE (angiotensin-converting enzyme), ACE2 (angiotensin-converting enzyme 2), ANP (atrial natriuretic peptide), CD2AP (CD2-adaptor protein), CTGF (connective tissue growth factor), CuZnSOD (copper zinc superoxide dismutase), iNOS (inducible nitric oxide synthase), MMP-9 (matrix metallopeptidase-9), MnSOD (manganese superoxide dismutase), nephrin, NOX4 (NADPH oxidase 4), podocin, and USP2 (ubiquitin-specific protease 2). For probe and primers sequences, see Supplementary Table 1 online.
Immunostainings
Glomerular ACE, ACE2, ANP (atrial natriuretic peptide), CTGF, and nitrotyrosilated proteins were quantified by immunostainings on 4 μm paraffin kidney sections. Kidney sections were incubated with the following primary antibodies: mouse anti-ACE (Chemicon, Temecula, CA, USA, dilution 1:100); goat anti-ACE2 (R&D Systems, Minneapolis, MN, USA, dilution 1:100); rabbit anti-ANP (Millipore, Billerica, MA, USA, dilution 1:200), rabbit anti-CTGF (Abcam, Cambridge, UK, dilution 1:200), rabbit anti-nitrotyrosine (Upstate, Lake Placid, NY, USA, dilution 1:100). For more details on these primary antibodies, see Supplementary Table 2 online. Biotinylated immunoglobulins (Vector Laboratories, Burlingame, CA, USA), were diluted 1:500 for ACE and ACE2, and 1:200 for ANP, CTGF, and nitrotyrosine and applied as secondary antibodies. Sections were counterstained with either Aniline blue (Ventana, France) or hematoxylin. A total of three sections per animal were analyzed, where a minimum of 20 glomeruli per section were counted. The percentage area occupied by the staining was calculated within the glomeruli. The results were expressed as the percentage of positive staining per glomerulus, i.e. percentage stained area.
Statistical analysis
The data were evaluated by analysis of variance (ANOVA) calculated using Statview 512 software for Apple Macintosh computer (Brainpower, Calabasas, CA, USA). Mean comparisons were performed by Fisher least significant difference method (post-hoc test). Linear regression analysis was used for testing two variable relationships. Results were expressed as mean ± SEM, unless otherwise specified. The criterion for statistical significance was p < 0.05.
Results
General parameters and biochemical data
Salt led to a significant increase in water intake, urinary volume, and sodium urinary excretion over 24 hours compared to the controls (Table 1). Aldosterone infusion significantly raised blood pressure levels and reduced serum potassium. Hypertension and hypokalemia were even greater when aldosterone was administered in combination with the salt diet and were significantly reduced by eplerenone, as shown in Table 1. In our study, aldosterone levels increased by 3.6 times in the rats infused with it, mimicking what is seen in patients with idiopathic hyperaldosteronism or aldosterone-producing adenomas where aldosterone levels increase by 2.67–3.82 times as compared to patients with essential hypertension. 21
General parameters and biochemical data.

Data are expressed as mean ± SEM; * p < 0.05 vs. CNT;
Renal structural features
Both salt and aldosterone led to kidney and tubular hypertrophy, as compared to the controls (Table 2). These effects were accentuated by combining salt to aldosterone, whereas they were blunted by eplerenone. Only aldosterone infusion, though, led to glomerular hyperthropy, which was significantly more pronounced in the high-salt diet fed rats, whilst it was reduced by eplerenone (Table 2). Consistent with these changes, aldosterone infusion only led to glomerular hypercellularity and to mesangial expansion, which corresponds to the dark pink staining that can be seen in Figure 1 and whose score is reported in Table 2. These changes were more pronounced in the high-salt diet fed rats, where they were partially reduced by eplerenone.
Glomerular structural and functional features.
Data are expressed as mean ± SEM; * p < 0.05 vs. CNT;

Effect of aldosterone, alone or in combination with salt, on mesangial matrix expansion. Representative periodic acid-Shiff (PAS) stained sections of kidneys (original magnification 25×): (a) CNT; (b) SALT; (c) ALDO; (d) ALDO+SALT; (e) EPL. CNT is 0.2% NaCl diet; SALT is 1.2% NaCl diet; ALDO is aldosterone and 0.2% NaCl diet; ALDO+SALT is aldosterone and 1.2% NaCl diet; EPL is aldosterone, eplerenone, and 1.2% NaCl diet.
Aldosterone and the glomerular filtration barrier
Aldosterone-induced glomerular structural changes (hypertrophy and mesangial expansion) were associated with a significant increase in glomerular Palb and AER (Table 2). In particular, there was a significant correlation between glomerular Palb and both glomerular hypertrophy (r = 0.550; p < 0.001) and AER (r = 0.516; p < 0.01). Having said that, we did not find any change in the glomerular gene expression of nephrin, podocin, CD2AP, and USP2 between the groups studied (data not shown). Moreover, urinary nephrin was almost undetectable in the groups studied (data not shown).
Glomerular renin–angiotensin system
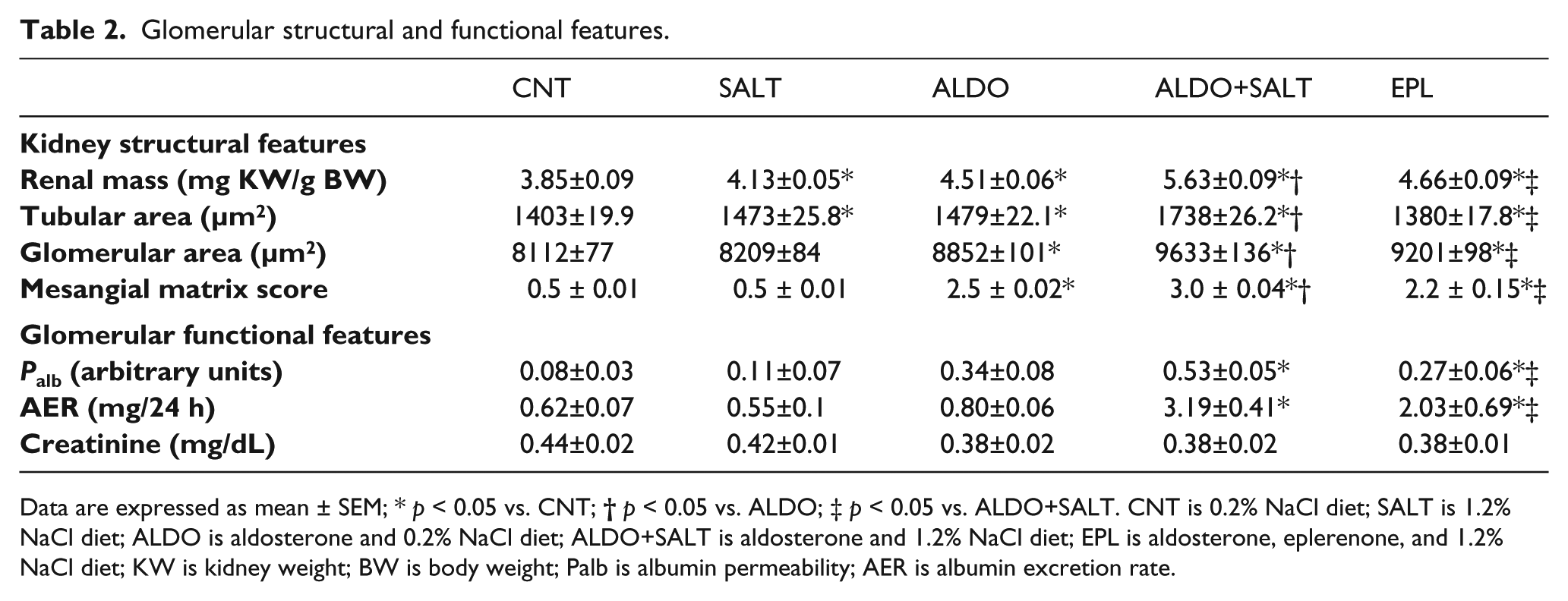
The gene expression of ACE, ACE2, and ANP was measured in the glomeruli of all the groups of rats, where we found that salt and aldosterone significantly upregulated ACE and downregulated ACE2 gene and protein expression (Figure 2). Similarly to ACE2, also ANP was significantly downregulated by aldosterone (Figure 2). All these changes were abolished by eplerenone (Figure 2).
Effect of aldosterone, alone or in combination with salt, on glomerular RAS. (a) Glomerular ACE, ACE2, and ANP messenger RNA expression, reported as relative gene units. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; ‡ p < 0.05 vs. ALDO+SALT. (b) Glomerular ACE, ACE2, and ANP protein expression, reported as percentage stained area. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; † p < 0.05 vs. ALDO; ‡ p < 0.05 vs. ALDO+SALT. (c–q) Representative ACE (upper panel), ACE2 (middle panel), and ANP (lower panel) immunostained sections of glomeruli (original magnification 25×): (c, h, m) CNT; (d, i, n) SALT; (e, j, o) ALDO; (f, k, p) ALDO+SALT; (g, l, q) EPL.
Glomerular pro-oxidative and profibrotic changes
Since the generation of oxidative stress and the balance between production and degradation of extracellular matrix play an important role in aldosterone-driven kidney damage, 3 we decided to evaluate the glomerular expression of molecules involved in these processes. In particular, we focused on CuZnSOD, iNOS, MnSOD, and NOX4 as well as on CTGF and MMP-9, whose gene expression was measured in the glomeruli previously isolated from the kidneys of all the groups of rats. With respect to oxidative stress generation, although the glomerular expression of MnSOD and NOX4 did not change between the groups studied (data not shown), CuZnSOD, which is an anti-oxidant, and iNOS, which is a pro-oxidant, were significantly down- and upregulated, respectively, in the glomeruli of the ALDO+SALT rats (Figure 3). The staining for nitrosylated proteins, which are a marker of oxidative stress, showed that there was a significant increase in glomerular nitrotyrosine in the SALT, ALDO, and ALDO+SALT groups, and that eplerenone reduced it significantly (Figure 3).

Effect of aldosterone, alone or in combination with salt, on glomerular oxidative stress. (a) Glomerular CuZnSOD and iNOS messenger RNA expression, reported as relative gene units. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; ‡ p < 0.05 vs. ALDO+SALT. (b) Glomerular nitrotyrosine protein expression, reported as percentage stained area. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; # p < 0.05 vs. SALT; † p < 0.05 vs. ALDO; ‡ p < 0.05 vs. ALDO+SALT. (c–g) Representative nitrotyrosine immunostained sections of glomeruli (original magnification 25X): (c) CNT; (d) SALT; (e) ALDO; (f) ALDO+SALT; (g) EPL. CNT is 0.2% NaCl diet; SALT is 1.2% NaCl diet; ALDO is aldosterone and 0.2% NaCl diet; ALDO+SALT is aldosterone and 1.2% NaCl diet; EPL is aldosterone, eplerenone, and 1.2% NaCl diet; CuZnSOD is copper zinc superoxide dismutase; iNOS is inducible nitric oxide synthase.
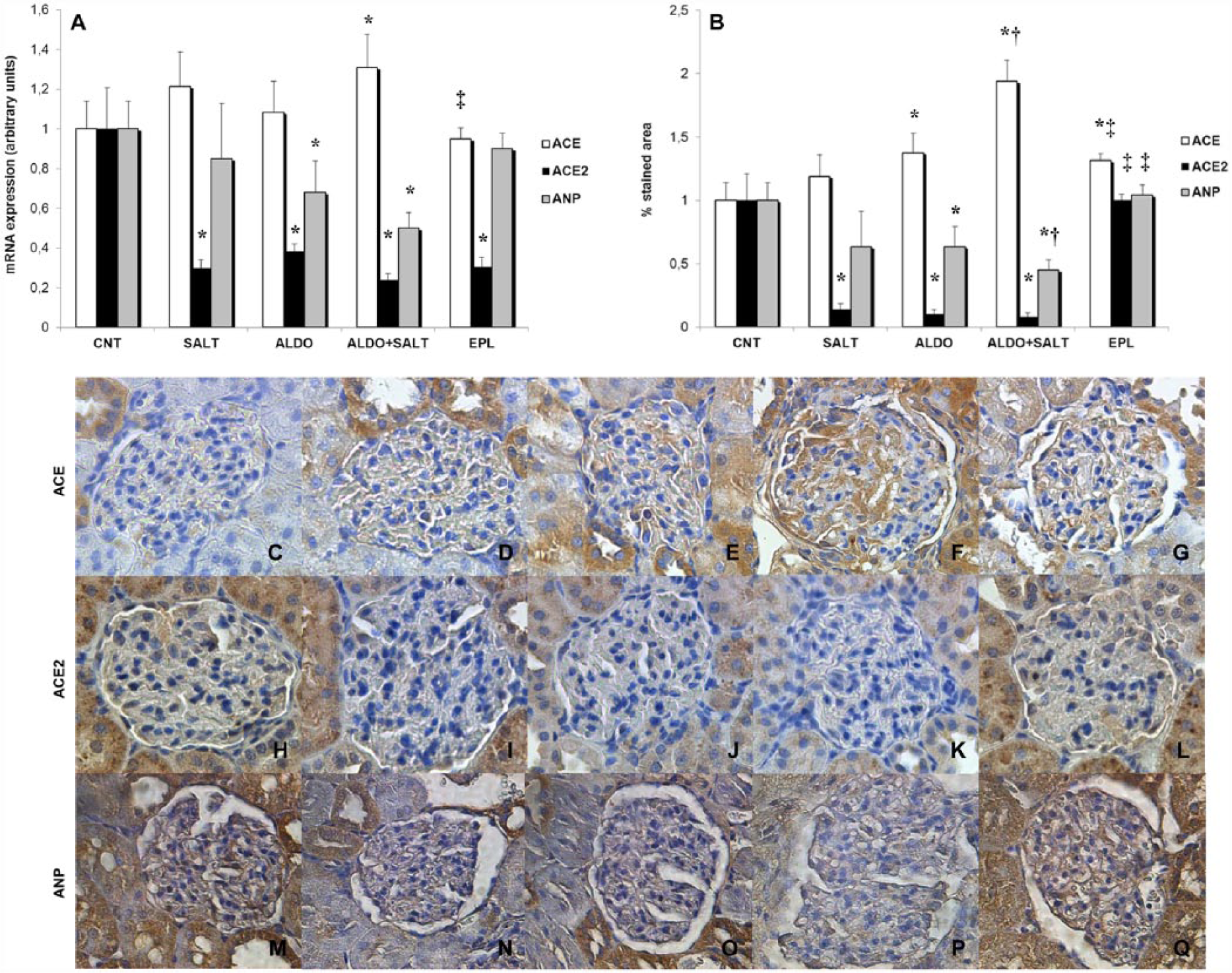
In terms of profibrotic changes, aldosterone combined with salt significantly upregulated CTGF and downregulated MMP-9 glomerular gene expression (Figure 4). This was consistent with CTGF immunostaining showing a significant increase of this protein expression in the glomeruli of the rats treated with aldosterone alone or in combination with salt (Figure 4). Eplerenone significantly reduced these effects (Figure 4).
Effect of aldosterone, alone or in combination with salt, on glomerular profibrotic changes. (a) Glomerular CTGF and MMP-9 messenger RNA expression, reported as relative gene units. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; ‡ p < 0.05 vs. ALDO+SALT. (b) Glomerular CTGF protein expression, reported as percentage stained area. Data are expressed as mean ± SEM. * p < 0.05 vs. CNT; ‡ p < 0.05 vs. ALDO+SALT. (c–g) Representative CTGF immunostained sections of glomeruli (original magnification 25×): (c) CNT; (d) SALT; (e) ALDO; (f) ALDO+SALT; (g) EPL. CNT is 0.2% NaCl diet; SALT is 1.2% NaCl diet; ALDO is aldosterone and 0.2% NaCl diet; ALDO+SALT is aldosterone and 1.2% NaCl diet; EPL is aldosterone, eplerenone, and 1.2% NaCl diet; CTGF is connective tissue growth factor; MMP-9 is matrix metallopeptidase.
Discussion
This work aimed at investigating the glomerular effects of aldosterone, alone and in combination with salt, and we found that aldosterone damages the glomerulus, where it induces structural and functional changes. In particular, this is the first study where the glomerular permeability to albumin has been measured in aldosterone-infused rats. Here we found that aldosterone significantly increased the glomerular permeability to albumin and albuminuria, which were positively correlated, in line with the concept that albuminuria is related to an alteration of glomerular permeability. To test the hypothesis that aldosterone increases glomerular permeability by damaging the slit diaphragm,22–23 we studied three important proteins of the slit diaphragm, which are nephrin, podocin, and CD2AP, whose mutations have been linked to nephrotic syndrome development. 24 In addition, we also quantified USP2, which seems to play a role in mesangial cell proliferation during glomerulonephritis. 25 Contrary to what has been previously reported, 22 we found no changes in the glomerular expression of these genes. Likewise, urinary nephrin was almost undetectable, suggesting that there was no podocyte loss. A first explanation to this negative data may be ascribed to the percentage of salt of our diet as compared to the 8% salt diet used by Shibata et al., 22 given that the 8% salt diet usually worsens glomerular damage as compared to the 1.2% salt diet. 18 Secondly, it is also possible that aldosterone needs a much longer period of time before it damages the podocytes, 26 consistent with the observation that the downregulation of slit diaphragm proteins correlates with the stage of glomerular damage. 27 In the third place, proteinuria does not only rely on podocyte changes,28,29 but it can also be ascribed to abnormalities of endothelium and basement membrane. 30
Consistent with the concept that a high-salt diet has a detrimental effect on kidney function, 18 and that it worsens the glomerular changes due to aldosterone, 15 here we found that the ALDO+SALT group displayed the highest degree of glomerular permeability to albumin and proteinuria. The fact that salt worsens the effect of aldosterone is due to the ability of salt to cause paradoxical MR activation through Rac1, 31 which modulates MR activity, as the inhibition of the Rac-1-MR cascade by eplerenone ameliorates salt-mediated kidney injury. 16 Here we found that eplerenone reduced most, but not all, aldosterone-driven effects. This suggests that mechanisms other than MR activation are involved in the aldosterone/salt-induced nephropathy, such as the non-genomic effects that aldosterone can exerts in the absence of MR activation. 32 Moreover, as suggested by other authors, 33 the reason underlying the discrepancy between the degree of aldosterone-induced renal damage and the modest effect of eplerenone is that aldosterone levels may be too high to be efficiently counteracted by MR antagonists.
In recent years there has been a paradigm shift in our understanding of aldosterone actions, which go far beyond sodium and potassium transport in the renal tubule and involve the ability of this hormone to stimulate the local RAS,4,5 generate reactive oxygen species, 6 and promote fibrosis, 7 whereby it leads to organ damage. 7 It is well known that the activation of the RAS is involved in the development and progression of kidney disease, as its blockade is one of the most efficient ways to reduce proteinuria as well as to delay renal insufficiency. 3 Such blockade has traditionally focused on inhibiting the synthesis of angiotensin (Ang) II and/or preventing activation of Ang II type 1 receptor. Nevertheless, the degradation of Ang II is also important, 34 and the major enzyme that converts Ang II to Ang 1-7 is ACE2. 35 So, it is the balance between ACE and ACE2 that is currently considered critical for the activation of renal RAS.36,37 Having said that, the second novel aspect of this work is that we show the effect that aldosterone, alone and in combination with salt, has on ACE2 glomerular expression in vivo. Here salt and aldosterone significantly reduced ACE2 expression, while ACE increased, leading to an upregulation of ACE/ACE2 ratio, which is in line with previous observations.38,39 Now, based on recent experimental evidence,40–42 it is possible to speculate that the glomerular changes that follow aldosterone infusion are partly relying on ACE2 downregulation. First of all, ACE2 deficiency/inhibition alone can increase glomerular ACE gene and protein expression,18,43 with a subsequent increase of local Ang II. Secondly, ACE2 downregulation contributes to aldosterone-induced glomerular changes, as ACE2 deficiency has been found associated with pro-oxidative, pro-inflammatory, and profibrotic glomerular changes.40–42 It should be noted that ACE2 protects the kidney not only by Ang II degradation, but also by Ang 1-7 and ANP generation, 40 which are both renoprotective.44–46 In line with the concept that ACE2 regulates ANP renal production, 40 here we found that aldosterone significantly reduced also ANP glomerular expression, which is the third novel aspect of this paper.
Besides RAS activation, which interacts with aldosterone in inducing tissue damage, aldosterone led to pro-oxidative and profibrotic changes, which were reduced by eplerenone and can also underpin the glomerular changes that we observed. On one hand, reactive oxygen species (ROS) production might have in fact contributed to the glomerular functional changes induces by aldosterone, as superoxide and hydroxyl radicals increase the glomerular permeability to albumin (possibly by lipid peroxidation of cell membrane cytoskeleton), and this is reversed by blocking these mediators with scavengers. 47 On the other hand, CTGF upregulation could also explain glomerular structural and functional changes. CTGF, which is not normally expressed in the healthy kidney, is induced in the early stages of renal diseases, 48 and its levels correlate with the severity and progression of renal fibrosis. 49 Interestingly, it has been shown that CTGF induces mesangial cell cycle arrest, mesangial cell hypertrophy, and mesangial matrix expansion in renal non-epithelial cells, 50 as well as that CTGF can damage the slit diaphragm. 51 Moreover, treatment of diabetic mice with CTGF antisense oligonucleotide attenuates albuminuria. 52
In conclusion, our work further characterizes the effects of aldosterone on the glomerulus. Here we show that aldosterone infusion induces glomerular hypertrophy, mesangial expansion, and it increases the glomerular permeability to albumin. These aldosterone-induced glomerular effects are likely to be mediated by the binding to the mineralocorticoid receptors, as eplerenone reverses them. In particular, we speculate that some of the aldosterone effects might depend on RAS activation, as aldosterone significantly reduced ACE2, increased the ACE/ACE2 ratio, and decreased ANP, which were partly reversed by eplerenone. Besides RAS activation, aldosterone led also to local oxidative stress generation and profibrotic changes, which altogether might have contributed to the induction of glomerular damage.
Footnotes
Conflict of interest
The authors declare that there are no conflicts of interest.
Funding
This work was supported in part by the Ministero dell’Istruzione dell’Università e della Ricerca (MIUR) (grant number 2006060985_004).