
Abstract
Objective:
The aim of this study was to investigate whether MiR-30a regulates autophagy by regulating the Beclin-1 protein, which is the marker for autophagosomes during myocardial injury, when induced by angiotensin II (Ang II).
Methods:
We randomly assigned 20 rats into two equal groups: Control group and Ang II group. We detected the expression of MiR-30a by quantitative real-time polymerase chain reaction (RT-PCR), and we employed western blotting to detect the protein expression of Beclin-1.
Results:
In this study, we found that Ang II induced cardiomyocyte autophagy, together with down-regulation of MiR-30a and upregulation of the Beclin-1 protein. We also found that the Beclin-1 protein is regulated by MiR-30a, by transferring a MiR-30a mimic or AMO-204 into the cardiomyocytes.
Conclusion:
These studies provided evidence that MiR-30a plays an important role in regulating autophagy through the Beclin-1 protein, during myocardial injury induced by Ang II.
Keywords
Introduction
Autophagy plays a critical role in growth and development, and its physiological function is to be involved in the normal growth and development process of cells and in maintaining metabolic balance; to promote the production of amino acids by self-degradation when nutritional deficiencies occur1–3 and under certain pressure conditions, such as in the presence of injury or nutritional deficiencies, to remove unwanted or injured cells in organs. Autophagy is a type of programmed cell death. It has been suggested to be essential for cellular homeostasis. It can determine cellular survival, together with apoptosis and necrosis.4,5
Autophagy activity levels are very low in physiological conditions, and it is upregulated in many pathophysiological processes.6,7 Because cardiomyocytes are terminally-differentiated cells that cannot divide again, suitable autophagy is essential for the maintenance of cardiomyocyte homeostasis. So autophagocytic deficiencies or excesses are associated with many cardiac pathologies, such as ischemia and heart failure. 8–10 It seems that modest levels of autophagy appear to be protective, while high levels of autophagy may cause self-digestion and promote cell death. 11
Autophagy is regulated by many autophagy-related genes (Atgs), which are involved in autophagosome formation.12,13 Beclin-1 is an important marker of autophagy. 14 It is possible to control the process of autophagy by up-regulating or down-regulating microtubule-associated protein light chain3 (LC3): The molecular mechanism for this effect has yet to be elucidated. As we know, it has been demonstrated that microRNAs, which negatively regulate protein expression in diverse biological and pathological processes, play an important role in myocardial injury.15–17 It was observed that many microRNAs regulate cell apoptosis18–21; however, it is well known that when apoptosis is blocked, the cells that preferentially die by apoptosis, may die by autophagy. 22 So it is beneficial for cell survival if autophagy becomes inhibited, together with apoptosis. We found that MiR-30a, which has an anti-apoptotic effect, may also regulate Beclin-1 expression through the nine complementary bases, according to the bioinformatics of Targetscan. Thus, the present study was undertaken to see whether MiR-30a was dysregulated by angiotensin II (Ang II), and if it may inhibit autophagy during hypoxia-reoxygenation, by regulating Beclin-1.
Materials and methods
Animal care
All animal experiments were approved by the Animal Research Ethics Committee of Guangzhou Medical University, Guangzhou, China. The investigation conformed to the guide for the care and use of laboratory animals published by the US National Institutes of Health (NIH). We randomly assigned 20 rats into two equal groups: Control group (Con group) and Ang II group (n = 10 each).
LDH assay
We collected serum from blood after 180 min of reperfusion, for determination of lactate dehydrogenase (LDH) levels.
Quantitative real-time RT-PCR of MiR-30a
Total RNA of cells was isolated by using TRIzol reagent, and then reverse transcribed according to the manufacturer’s instructions (Fermentas, Shanghai, China). The annealing temperature of miRNA-204 was set at 60°C. The comparative threshold cycle (Ct) method with the arithmetic formula (Equation (1)) below was used to determine relative quantitation of gene expression of both target and housekeeping genes (β-actin):

The MiR-30a primers used in the study are shown in Table 1.
Primers used for quantitative real-time RT-PCR.

PCR: polymerase chain reaction.
Western blot
We determined protein concentrations with the BCA protein assay kit (Amersham Biosciences, Piscataway, NJ, USA), according the manufacturer’s protocol. Equal amounts of protein (60 µg) from the cardiomyocytes were subjected to Western blot analysis, to evaluate Beclin-1 expression with an ECL detection kit (Amersham Biosciences, Piscataway, NJ). Beclin-1 immunoreactivity was detected using a rabbit antiserum specific for rat Beclin-1 protein (Sigma, St. Louis, MO, USA) as primary antibodies. Detection of antigen-antibody complex formation was performed with horseradish peroxidase (HRP)-conjugated goat anti-rabbit secondary antibody. The Beclin-1 concentrations were normalized with the values for β-actin concentration.
Statistical analysis
Quantitative data are presented as mean ± standard error (SE). Statistical significance was determined using the T test or 1-way analysis of variance (ANOVA); p < 0.05 was considered statistically significant.
Results
Myocardium injury was induced by Ang II
The extent of myocardial infarction was evaluated after injury. Representative photographs of midventricular cross-sections of Evans blue and Triphenyltetrazolium chloride (TTC)-stained hearts were taken from the Control and Ang II groups. Infarct area (INF)/ area at risk (AAR) and LDH are shown in Figure 1.

The heart injury induced by IR.
Ang II decreased the expression of MiR-30a
To demonstrate the effect of Ang II on MiR-30a, we compared the MiR-30a between the Con group and Ang II group (n = 10 each). Using RT-PCR, we found that Ang II significantly decreased MiR-30a (Figure 2).
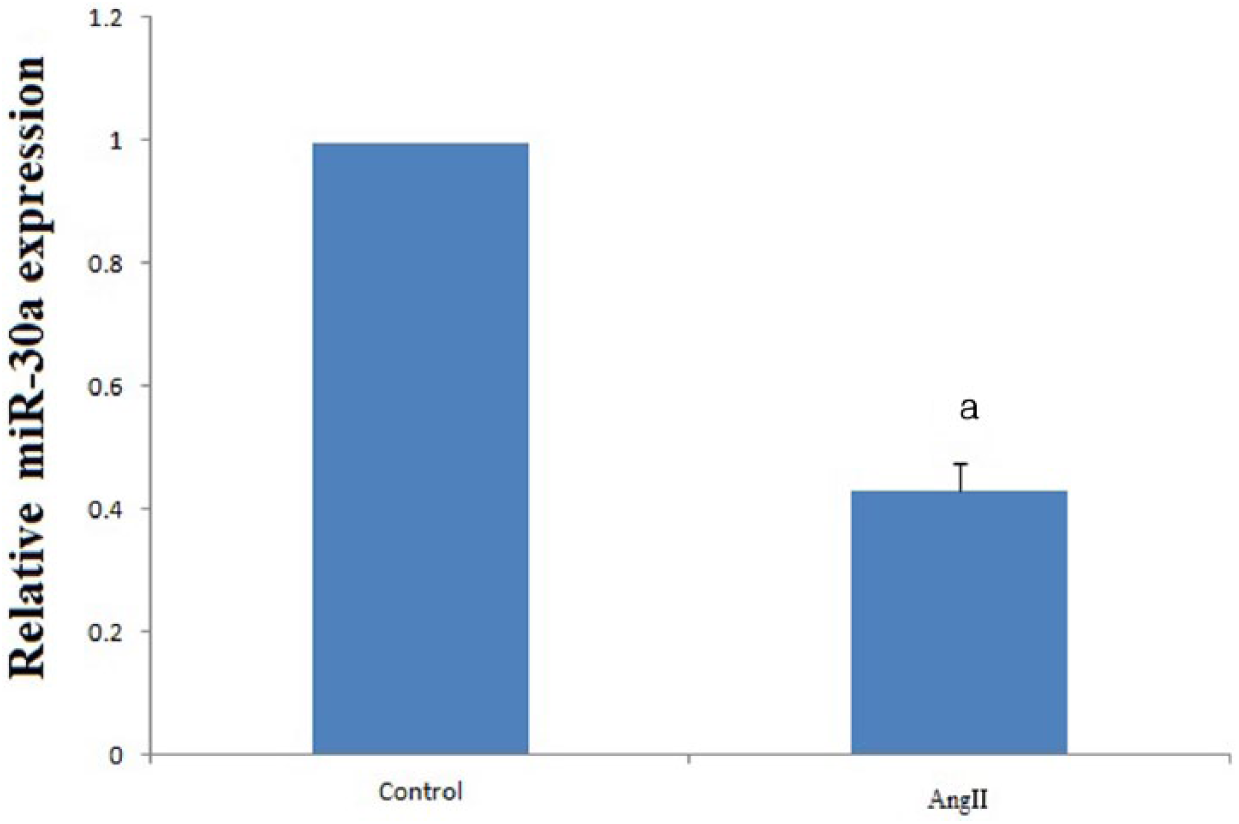
Results of MiR-30a expression with RT-PCR after IR injury. It was found that MiR-30a was down-regulated by IR (n = 10 each; p = 0.026 < 0.05).
Ang II up-regulated the protein level of Beclin-1
As a marker of autophagosomes, the protein level of Beclin-1 represents the amount of autophagosomes present; thus, we compared the ratio of Beclin-1/β-actin between the control group and IR group. It has been reported that autophagy contributes to cell death when apoptosis is inhibited, and sometimes the early stages of autophagy were required for apoptosis. Beclin-1 was the marker of autophagosomes, so the ratio of Beclin-1/β-actin could stand for the level of autophagy. In our study, we found that Ang II up-regulated the protein expression of Beclin-1, together with increasing the ratio of the Ang II group (n = 10); and we found that Beclin-1 was enhanced by Ang II (Figure 3).
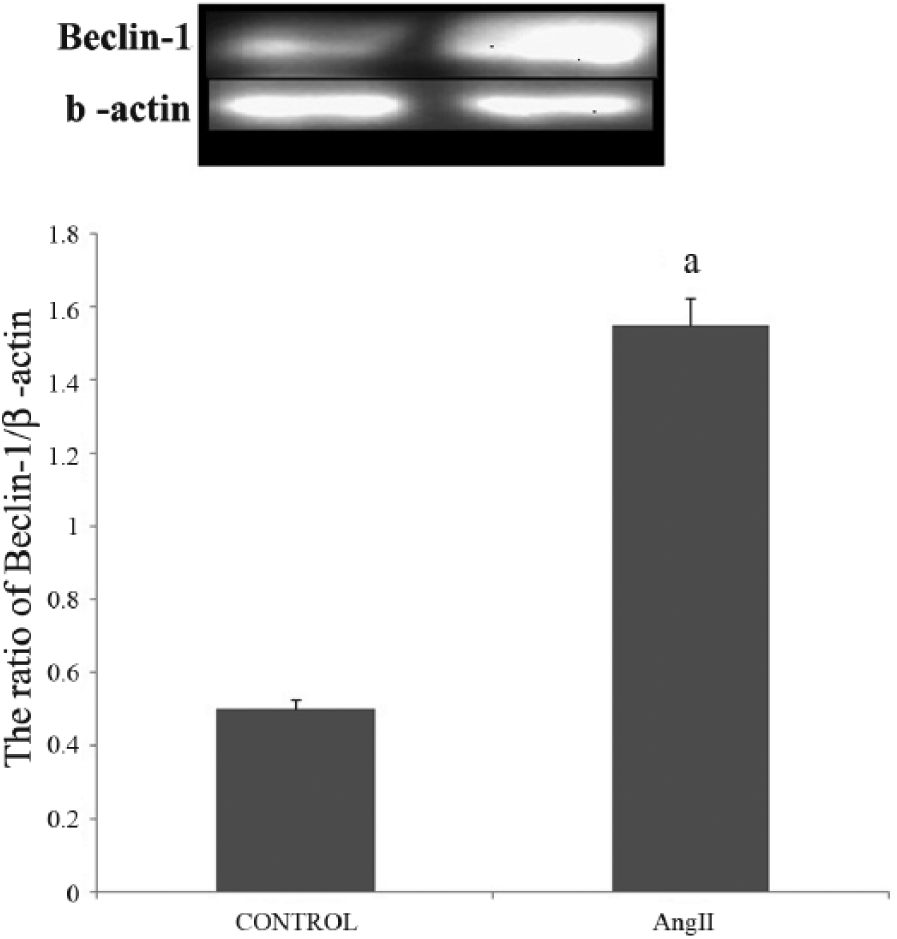
Results of Beclin-1 protein expression with western blot, after IR injury.
Discussion
Autophagy is a lysosome-dependent degradation pathway widely existent within eukaryotic cells as a self-protection mechanism in response to harmful stimulation; and the functions of autophagy are not only to be involved in surrounding and isolating damaged organelles, but more importantly, in transporting the surrounded materials to lysosomes for degradation, and thus to produce amino acids, ATP and other substances in order to maintain the energy metabolism cycle of the body.
Autophagy has attracted great interest because it is involved in many physiological processes. If autophagy destroys the cytosol and organelles beyond a certain threshold, autophagic cell death will occur. Autophagy was detrimental during reperfusion, although it protected the cardiomyocytes during ischemia, as demonstrated by Matsui et al. 23 and his autophagy cell. Thus, it would be beneficial for revascularized hearts to find a method of regulating Beclin-1 expression. Non-coding RNAs regulate gene expression in a sequence-dependent manner: They are endogenous regulators of gene expression, demonstrated to be involved in cardiac injury.
According to the bioinformatics of Targetscan, MiR-30a, which has an anti-apoptosis effect, may regulate the expression of Beclin-1. In our study, we found that Ang II could down-regulate MiR-30a, together with up-regulating Beclin-1 protein. When a MiR-30a mimic was transferred into cardiomyocytes, Beclin-1 protein was attenuated; while Beclin-1 protein was up-regulated by AMO-204, which was concentration-dependent. But LC3-I was not regulated by MiR-30a, as was seen in the previous study. These results demonstrated that MiR-30a may regulate cardiomyocyte autophagy through Beclin-1, during injury induced by Ang II.23,24
Our rat study results demonstrated that MiR-30a plays an important role by regulating the Beclin-1 protein during Ang II damage, so it became possible to control the autophagy under a beneficial threshold by regulating MiR-30a expression, to protect cardiomyocytes against injury induced by Ang II.
Footnotes
Conflict of interest
None declared.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.