
Abstract
Background:
Perivascular adipose tissue is implicated in vasoreactivity; however, its effect on atherosclerosis remains undefined.
Methods and results:
We examined the effect of a high-cholesterol diet (HCD) on phenotypic alterations of the thoracic periaortic adipose tissue (tPAT) in apoE-deficient (apoE–/–) mice. Gene expression of the components of the renin angiotensin system and that of macrophage markers were significantly higher in apoE–/– mice fed an HCD than in those fed a chow diet (CD). These changes were absent both in angiotensin II (AngII) receptor blocker (ARB)-treated apoE–/– mice and in Ang II type 1 (AT1) receptor-deficient apoE–/– (Agtr1–/–/apoE–/–) mice. To evaluate their effect on atherosclerosis, we transplanted tPAT into apoE–/– mice alongside the distal abdominal aorta. Transplanted tPAT was harvested from apoE–/– and Agtr1–/–/apoE–/– mice fed a CD (tPAT-CD/apoE–/–, tPAT-CD/Agtr1–/–/apoE–/–), HCD (tPAT-HCD/apoE–/–, tPAT-HCD/Agtr1–/–/apoE–/–), or HCD in combination with ARB treatment (tPAT-HCD/ARB/apoE–/–). Four weeks after transplantation, a significantly increased oil red O-positive area was observed in the aorta of tPAT-HCD/apoE–/– mice than in tPAT-CD/apoE–/– mice. Such a change was absent in tPAT-HCD/ARB/apoE–/– and tPAT-HCD/Agtr1–/–/apoE–/– mice.
Conclusions:
Our findings demonstrated that AT1 receptor plays a crucial role in HCD-induced phenotypic alterations of tPAT, modulation of which could exert beneficial effects on atherosclerosis.
Keywords
Introduction
Visceral adipose tissue is not only a storage depot of excess energy, but also an endocrine organ that secretes a number of factors into circulation, some of which exaggerate systemic inflammation, leading to the development of insulin resistance and cardiovascular diseases.1–5 Perivascular adipose tissue (PAT), one of the ectopic adipose tissues, is thought to serve as a structural support for blood vessels. However, recent studies show that PAT secretes numerous vasoactive molecules involved in vascular remodeling and vasoreactivity.6–10 Takaoka et al. found elevated expression of pro-inflammatory genes and lower adiponectin expression in the PAT of obese mice, and concluded that this contributed to exaggerated neointimal formation following arterial injury. 11 Chatterjee et al. also found elevated pro-inflammatory gene expression and diminished adiponectin expression in the PAT surrounding diseased human coronary arteries. 12 These findings suggested that, through its paracrine effects on vasculature, PAT influenced the development of atherosclerosis. However, the precise roles of PAT in atherogenesis remained unknown.
Thoracic periaortic adipose tissue (tPAT) surrounding the descending thoracic aorta has similar characteristics to those of brown adipose tissue 13 and has recently become a focal point of cardiovascular disease risk assessment. Fox et al. reported that the volume of tPAT, quantified by computed tomography, correlated with the incidence of peripheral arterial disease. 14 These authors also found that a high volume of tPAT in the absence of a high volume of visceral adipose tissue correlated with a higher prevalence of cardiovascular disease. 15 Considering that phenotypic alterations of tPAT, such as higher expressions of pro-inflammatory cytokines accompanied by inflammatory leukocytes infiltration, were observed in mice fed a high-fat diet (HFD), 12 these changes likely had a proatherogenic effect. However, Chang et al. recently reported that, owing to reduced thermogenic capacity, the absence of tPAT in their gene-manipulated mouse model exaggerated atherosclerosis development upon cold exposure. 16 These findings led us to hypothesize that, depending on the phenotypic alterations that occur under specific conditions, tPAT plays contrasting roles in the development of atherosclerosis.
We have previously reported that tPAT-specific activation of the renin angiotensin system (RAS) occurred in uninephrectomized apoE-deficient (apoE–/–) mice, and that this was partially responsible for the accelerated atherosclerotic development observed in chronic kidney disease. 17 Here, we examined high-cholesterol diet (HCD)-induced phenotypic alterations in tPAT, and investigated their roles in atherosclerosis development using a tPAT transplantation model. An HCD promoted the expression of RAS components as well as that of macrophage markers. Such changes were not observed in epididymal white adipose tissue (WAT). These tPAT-specific phenotypic alterations were absent both in angiotensin II (Ang II) receptor blocker (ARB)-treated apoE–/– mice and in Ang II type 1 (AT1) receptor-deficient apoE-/- (Agtr1–/–/apoE–/–) mice. We demonstrate for the first time that, compared to tPAT from chow diet-(CD) fed apoE–/– mice, transplantation of tPAT from HCD-fed apoE–/– mice significantly exaggerates atherosclerosis development in the entire aorta. Further, transplantation of tPAT from ARB-treated apoE–/– mice or Agtr1–/–/apoE–/– mice markedly inhibited atherosclerosis development when compared to tPAT transplantation from control mice. Our findings suggest that HCD-induced phenotypic alteration of tPAT, mediated by AT1 receptor, directly contributes to atherosclerosis development, and that therapeutic targeting of the phenotypic changes of tPAT could potentially remediate and prevent cardiovascular diseases.
Methods
Animals
ApoE–/– mice (C57BL/6) were obtained from Taconic Co., Ltd. (Germantown, NY, USA). AT1a receptor-deficient (Agtr1–/–) mice (C57BL/6) were obtained from Tanabe Seiyaku Co., Ltd. (Osaka, Japan). Agtr1–/– mice were crossed with apoE–/– mice. Heterozygous animals were crossed until homozygous double knockout (Agtr1–/–/apoE–/–) mice were obtained (backcrossed > 10 times). Eight-week-old male apoE–/– mice were fed a CD or HCD (36% fat, 1.25% cholesterol; Oriental Yeast Co., Tokyo, Japan) in combination with ARB (Olmesartan, 1 mg/kg/day) or saline (using an osmotic minipump) for four or eight weeks. Eight-week-old male Agtr1–/–/apoE–/– mice were fed a CD or HCD for four or eight weeks. The animals were housed in a room that was maintained at 22°C under a 12-hour light/dark cycle and were provided with drinking water ad libitum. Mice were anaesthetized using isoflurane (2%, 0.2 ml/min) throughout the surgery during transplantation. The depth of anesthesia was confirmed by lack of tail pinch response. A laparotomy was performed under sterile conditions with the assistance of an operating stereomicroscope. A periaortic fat pad near the descending thoracic aorta (tPAT) was harvested from 16-week-old donor apoE–/– mice, fed for eight weeks either with a CD (tPAT-CD), an HCD (tPAT-HCD), or an HCD in combination with ARB treatment (tPAT-HCD/ARB), and subsequently transplanted over the endogenous infrarenal abdominal periaortic adipose tissue of 20-week-old apoE–/– mice fed an HCD for 12 weeks. A periaortic fat pad from 16-week-old donor Agtr1–/–/apoE–/– mice, fed for eight weeks with a CD or HCD, was similarly transplanted into 20-week-old apoE–/– mice. Finally, the musculofascial and skin incisions were sutured. Sham surgeries of control animals were performed in the same manner, but without transplantation of tPAT. Mice were anaesthetized before the implantation of osmotic minipumps. At 24 weeks of age, the area of the atherosclerotic lesion in the aorta was evaluated. Before harvesting the tissues (aortas), mice were euthanized by trans-cardiac perfusion under terminal anesthesia induced by intraperitoneal administration of pentobarbital (200 mg/kg). All experiments were performed by strictly adhering to Directive 2010/63/EU of the European Parliament and to the Guidelines for Animal Experiments of the Kyoto Prefectural University of Medicine, following approval by a local university ethics review board.
Hemodynamic analysis
Mean blood pressure and heart rates were measured under conscious and unrestrained conditions using a programmable sphygmomanometer (BP-98A; Softron, Tokyo, Japan).
Plasma lipid analysis
Measurements of total cholesterol, triglyceride, low-density lipoprotein (LDL)-cholesterol, and high-density lipoprotein (HDL)-cholesterol were outsourced to SRL, Tokyo, Japan.
Quantitative measurement of atherosclerotic lesions
Mice were euthanized at the age of 24 weeks, and atherosclerotic lesions were analyzed. Imaging and analysis of oil-red O-stained aortas were accomplished using Scion Imaging Software. The aortic lesion area in each animal was expressed as the percentage of lesion area per total aortic area.
Immunohistochemistry
tPAT was quickly removed after phosphate-buffered saline (PBS) perfusion, embedded in paraffin, and immunofluorescently labeled using a combination of anti-mouse Mac2 antibody (CEDARLANE, Burlington, Ontario, Canada) and Alexa Fluor 488-conjugated secondary antibodies (Invitrogen, Carlsbad, CA, USA). Nuclei were labeled using diamidino-2-phenylindole (DAPI) solution (DOJINDO, Kumamoto, Japan), and the sections were examined using an LSM 510 META confocal microscope (Carl Zeiss, Jena, Germany). For a negative control, non-immune immunoglobulin and Alexa Fluor 488-conjugated secondary antibodies were used. The number of Mac2-positive cells was assessed using data collected from five sections and six animals in each group.
Real-time polymerase chain reaction (PCR)
Total RNA was extracted from adipose tissue and reverse transcribed to prepare cDNA. Real-time PCR was performed on a Thermal Cycler Dice (Takara Bio, Shiga, Japan), using SYBR Premix Ex Taq 2 (Takara Bio, Shiga, Japan). Dissociation curves were examined for the aberrant formation of primer dimers. PCR-amplified products were electrophoresed on 2% agarose gels to confirm the presence of a single amplicom. Data were expressed as gene expression levels relative to that of control.
Enzyme-linked immunosorbent (ELISA) assay
Blood from the right ventricular was collected in tubes containing citrate at a final concentration of 0.01 M. Serum was separated by centrifugation at 1000 g for 20 minutes, and stored −80°C. Adipose tissue was homogenized in 600 µl buffer containing PBS and 0.04% Tween 80 using a rotary homogenizer. Homogenized tissue samples were centrifuged at 10,000 g for 10 minutes and the supernatants were collected. The amounts of tumor necrosis factor alpha (TNF-α), interleukin-6 (IL-6) and adiponectin present in the serum were estimated using commercial ELISA kits (MTA00B, M6000B and MRP300, R&D Systems, Minneapolis, MN). The levels of TNF-α, IL-6 and adiponectin in the adipose tissue were estimated using ELISA kits (cat.88-7324 and cat.88-7064, eBioscience, San Diego, CA, and MRP300, R&D Systems, Minneapolis, MN). The assays were carried out according to the manufacturers’ instructions.
Statistical analysis
All data are expressed as the mean ± SE. Mean values were compared using ANOVA. If a statistically significant effect was found, Scheffe’s F test was performed to analyze the differences between the groups. p < 0.05 was considered statistically significant.
Results
Periaortic adipose tissue-specific activation of the RAS in HCD-fed apoE–/– mice
To examine the effect of an HCD on phenotypic alterations of adipose tissue, 8-week-old apoE–/– mice were fed either a CD or HCD until they reached 12 weeks of age. The body weight and body weight ratios of tPAT and epididymal WAT pads were not affected by HCD feeding (Figure 1(a)). The histomorphological analysis of both adipose pads revealed no discernible differences between the two groups of mice (Figure 1(b)). We compared the expression of adipocyte differentiation-related genes, RAS components, monocyte/macrophage markers, and adipocytokines. An HCD significantly upregulated the expression of adipocyte differentiation-related genes, including those of peroxisome proliferator-activated receptor-γ (PPARγ) and fatty acid binding protein 4 (FABP4) both in tPAT and WAT (Figures 2(a) and 2(b)). The expression levels of RAS components, including angiotensinogen (AGT), angiotensin-converting enzyme (ACE), and AT1a receptor (AT1aR), were significantly higher in the tPAT of HCD-fed mice, while AGT expression level was significantly lower in the WAT of HCD-fed mice (Figures 2(c) and 2(d)). Furthermore, the levels of monocyte/macrophage markers, including CD68 and CD206, were significantly higher in the tPAT of HCD-fed apoE–/– mice, whereas no such differences were observed in the WAT (Figures 2(e) and 2(f)). In both tPAT and WAT, the expression levels of the inflammatory adipokines, TNF-α, IL-6, and monocyte chemoattractant protein 1 (MCP-1), were not altered by the HCD (Figures 2(g) and 2(h)). These findings suggested that HCD feeding led to the activation of adipose tissue RAS in a tissue-specific manner, and that this was accompanied by increased accumulation of monocytes/macrophages.

The effects of a high-cholesterol diet on body weight, fat pad/body weight ratio, and microscopic appearance. (a) The body weights and fat pad/weight ratios of tPAT and epididymal WAT showed no difference in either between CD and HCD groups. (b) Histomorphological analysis of hematoxylin-eosin staining showing no discernable difference in both tPAT and epididymal WAT between CD and HCD groups. BW: body weight; tPAT: thoracic periaortic adipose tissue; WAT: white adipose tissue; CD: chow diet; HCD: high-cholesterol diet. The scale bar shows 200-µm intervals.

Periaortic adipose tissue-specific activation of the renin angiotensin system following high-cholesterol diet feeding. (a) and (b) Quantitative real-time PCR analysis of the mRNA expression levels of adipocyte differentiation-related genes in the tPAT (a) and epididymal WAT (b). Values are the mean ± SE relative to that in chow diet-fed mice (CD). Each group has at least six mice. *p < 0.05 vs. CD. PCR: polymerase chain reaction; tPAT: thoracic periaortic adipose tissue; WAT: white adipose tissue, CD: chow diet, HCD: high-cholesterol diet. (c) and (d) Quantitative real-time PCR analysis of the mRNA expression levels of RAS components in the tPAT (c) and epididymal WAT (d). Values are the mean ± SE relative to that in chow diet-fed mice (CD). Each group has at least six mice. *p < 0.05 vs. CD. RAS: renin angiotensin system; AGT: angiotensinogen; ACE: angiotensin-converting enzyme, AT1aR: angiotensin II type1a receptor. (e) and (f) Quantitative real-time PCR analysis of the mRNA expression of monocyte/macrophage marker genes in the tPAT (e) and epididymal WAT (f). Values are the mean ± SE relative to that in chow diet-fed mice (CD). Each group has at least six mice. *p < 0.05 vs. CD. PCR: polymerase chain reaction; tPAT: thoracic periaortic adipose tissue; WAT: white adipose tissue. (g) and (h) Quantitative real-time PCR analysis of mRNA expression of proinflammatory cytokines/chemokines in the tPAT (g) and epididymal WAT (h). Values are the mean ± SE relative to that in chow diet-fed mice (CD). Each group has at least six mice. PCR: polymerase chain reaction; tPAT: thoracic periaortic adipose tissue; WAT: white adipose tissue.
HCD-induced phenotypic alterations are significantly suppressed by angiotensin II receptor blockade or AT1 receptor deficiency
We next examined the involvement of the AT1 receptor in tPAT-specific HCD-induced phenotypic alterations. Neither ARB treatment nor AT1 deficiency (data not shown) affected pad weight. Four weeks of ARB treatment along with HCD significantly suppressed the mRNA expression of AGT, ACE and PPARγ (Figures 3(a) and 3(c)). Additionally, treatment with ARB blocker led to a significant reduction in the gene expression levels of the monocyte/macrophage markers, including F4/80 (Figure 3(e)). Notably, the gene expression of the proinflammatory adipokines, TNF-α, IL-6, and MCP-1, markedly diminished after ARB treatment (Figure 3(g)). The HCD-induced phenotypic alterations observed in apoE–/– mice were absent in Agtr1–/–/apoE–/– mice (Figures 3(b), 3(d), 3(f) and 3(h)). These findings suggested that the AT1 receptor plays a critical role in tPAT-specific HCD-induced phenotypic alterations, and that AT1 blockade could be used as a potential strategy to suppress inflammatory responses in the tPAT.

Angiotensin receptor blockade or AT1 receptor deficiency prevents high-cholesterol diet-induced phenotypic alterations in the tPAT. (a) and (b) Quantitative real-time PCR analysis of the mRNA expression of adipocyte differentiation-related genes in apoE–/– mice (a) and AT1 receptor-deficient apoE–/– (Agtr1–/–/apoE–/–) mice (b). Values are the mean ± SE relative to that in control mice. Each group has at least six mice. *p < 0.05 vs. control mice. AT1: Ang II type 1; tPAT: thoracic periaortic adipose tissue; CD: chow diet, HCD: high-cholesterol diet; HCD/ARB: high-cholesterol diet along with angiotensin receptor blocker (ARB) treatment. (c) and (d) Quantitative real-time PCR analysis of the mRNA expression levels of RAS components in apoE–/– mice (c) and AT1 receptor-deficient apoE–/– (Agtr1–/–/apoE–/–) mice (d). Values are the mean ± SE relative to that in control mice. Each group has at least six mice. *p < 0.05 vs. control mice. PCR: polymerase chain reaction; RAS: renin angiotensin system; AGT: angiotensinogen; ACE: angiotensin-converting enzyme; AT1aR: angiotensin II type1a receptor. (e) and (f). Quantitative real-time PCR analysis of the mRNA expression levels of monocyte/macrophage marker genes in apoE–/– mice (e) and AT1 receptor-deficient apoE–/– (Agtr1–/–/apoE–/–) mice (f). Values are the mean ± SE relative to that in control mice. Each group has at least six mice. *p < 0.05 vs. control mice. (g) and (h) Quantitative real-time PCR analysis of mRNA expression of proinflammatory adipokines in apoE–/– mice (g) and AT1 receptor-deficient apoE–/– (Agtr1–/–/apoE–/–) mice (h). Values are the mean ± SE relative to that in control mice. Each group has at least six mice. *p < 0.05 vs. control mice. PCR: polymerase chain reaction; AT1: Ang II type 1.
Intra-abdominal transplantation of the tPAT from ARB-treated apoE–/– mice prevents atherosclerotic development throughout the aorta
Intra-abdominal tPAT transplantation was performed to examine whether tPAT-specific phenotypic alterations contributed toward the development of atherosclerosis. The tPAT was harvested from 16-week-old apoE–/– mice fed a CD (tPAT-CD), an HCD (tPAT-HCD), or an HCD in combination with ARB administration (tPAT-HCD/ARB), and transplanted alongside the distal abdominal aorta of 20-week-old apoE–/– mice fed an HCD starting from 8 weeks of age. The weights of transplanted tPAT were 11.3 ± 0.7, 8.8 ± 0.5, and 6.0 ± 0.3 mg, respectively. The weights of tPAT from HCD-fed and ARB-treated HCD-fed apoE–/– mice were significantly lower than CD-fed apoE–/– mice (p < 0.05). Four weeks after transplantation, compared with tPAT-CD transplanted mice, the atherosclerotic lesion area of the entire aorta in tPAT-HCD transplanted mice showed a modest, yet significant, increase (29%, p < 0.01) (Figures 4(a) and 4(b)). The body weight and body weight ratios of epididymal WAT pads were also equivalent among the groups after fat transplantation (Supplemental Figures 1(a) and 1(b)). The hemodynamic parameters and lipid profiles did not differ between these two groups (Supplemental Figures 2(a) and 2(b)). In contrast, the areas of atherosclerotic lesions in tPAT-HCD/ARB transplanted mice were reduced by 34%, 24% and 42% compared with those of sham-operated control, tPAT-CD transplanted, and tPAT-HCD-transplanted mice, respectively (p < 0.01). These results suggested that transplantation of tPAT-HCD/ARB not only ameliorated the augmented atherosclerotic development as observed in tPAT-HCD transplanted mice, but also counteracted it when compared to no transplantation in the sham control. To further elucidate the mechanisms of tPAT transplantation-associated atherosclerosis development, we evaluated the atherosclerotic lesion area of the suprarenal and infrarenal aorta separately. The atherosclerotic lesion area of the infrarenal aorta differed only between tPAT-HCD- and tPAT-HCD/ARB-transplanted mice, suggesting that the anti-atherogenic actions of tPAT-HCD/ARB-transplanted mice are likely due to its paracrine effects on the vasculature of the abdominal aorta (Figure 4(c)). The atherosclerotic development of the suprarenal aorta followed similar trends as that in the entire aorta (Figure 4(d)), supporting the notion that transplanted tPAT exerted both systemic and local paracrine effects on atherogenesis.

Intra-abdominal transplantation of tPAT from high-cholesterol diet-fed apoE–/– mice exaggerates the atherosclerotic lesion development. (a) Representative photographs of oil-red O-stained entire aortas from sham control (a), tPAT-CD (b), tPAT-HCD (c), and tPAT-HCD/ARB (d). Arrows indicate right renal artery. The scale bar shows 5-mm intervals. (b) Quantitative analysis of the atherosclerotic lesion area in the entire aorta. (c) Quantitative analysis of the atherosclerotic lesion area in the infrarenal aorta. (d) Quantitative analysis of the atherosclerotic lesion area in the suprarenal aorta. Values are the mean ± SE for at least 11 mice in each group. *p < 0.05 vs. sham control. #p < 0.05 vs. tPAT-CD. †p < 0.05 vs. tPAT-HCD. tPAT-CD: apoE–/– mice transplanted with the tPAT from chow diet-fed apoE–/– mice. tPAT-HCD: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed apoE–/– mice. tPAT-HCD/ARB: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed apoE–/– mice treated with angiotensin receptor blocker. tPAT: thoracic periaortic adipose tissue; CD: chow diet; HCD: high-cholesterol diet; ARB: angiotensin II (AngII) receptor blocker.
Intra-abdominal transplantation of the tPAT from HCD-fed Agtr1–/–/apoE-/- mice ameliorates atherosclerosis development throughout the aorta
To investigate the effects of AT1 receptor deficiency on tPAT transplantation-associated atherosclerosis, the tPAT was harvested from 16-week-old Agtr1–/–/apoE–/– mice fed a CD (tPAT-CD/Agtr1–/–/apoE–/–) or HCD (tPAT-HCD/Agtr1–/–/apoE–/–) for eight weeks, and transplanted into 20-week-old recipient apoE–/– mice fed an HCD from 8 weeks of age. The weights of transplanted tPAT from HCD-fed Agtr1–/–/apoE–/– mice were significantly lower than CD-fed Agtr1–/–/apoE–/– mice (7.6 ± 0.6 vs. 14.0 ± 1.6 mg, p < 0.05). A significant reduction in the atherosclerotic lesion area was observed in tPAT-HCD/Agtr1–/–/apoE–/–-transplanted mice compared with tPAT-CD/Agtr1–/–/apoE–/–-transplanted animals (30%, p < 0.01) (Figures 5(a) and 5(b)), a result that was in contrast with the effects of tPAT transplantation from apoE–/– mice. The body weight and body weight ratios of epididymal WAT pads were also equivalent among the groups after fat transplantation (Supplemental Figures 1(c) and 1(d)). Hemodynamic parameters and lipid profiles did not differ between the two groups (data not shown). A significant reduction in the atherosclerotic lesion area was observed both in the suprarenal and infrarenal aortas (Figures 5(c) and 5(d)), suggesting that the inhibitory effect of AT1 receptor deficiency on atherogenesis occurred at the systemic level.
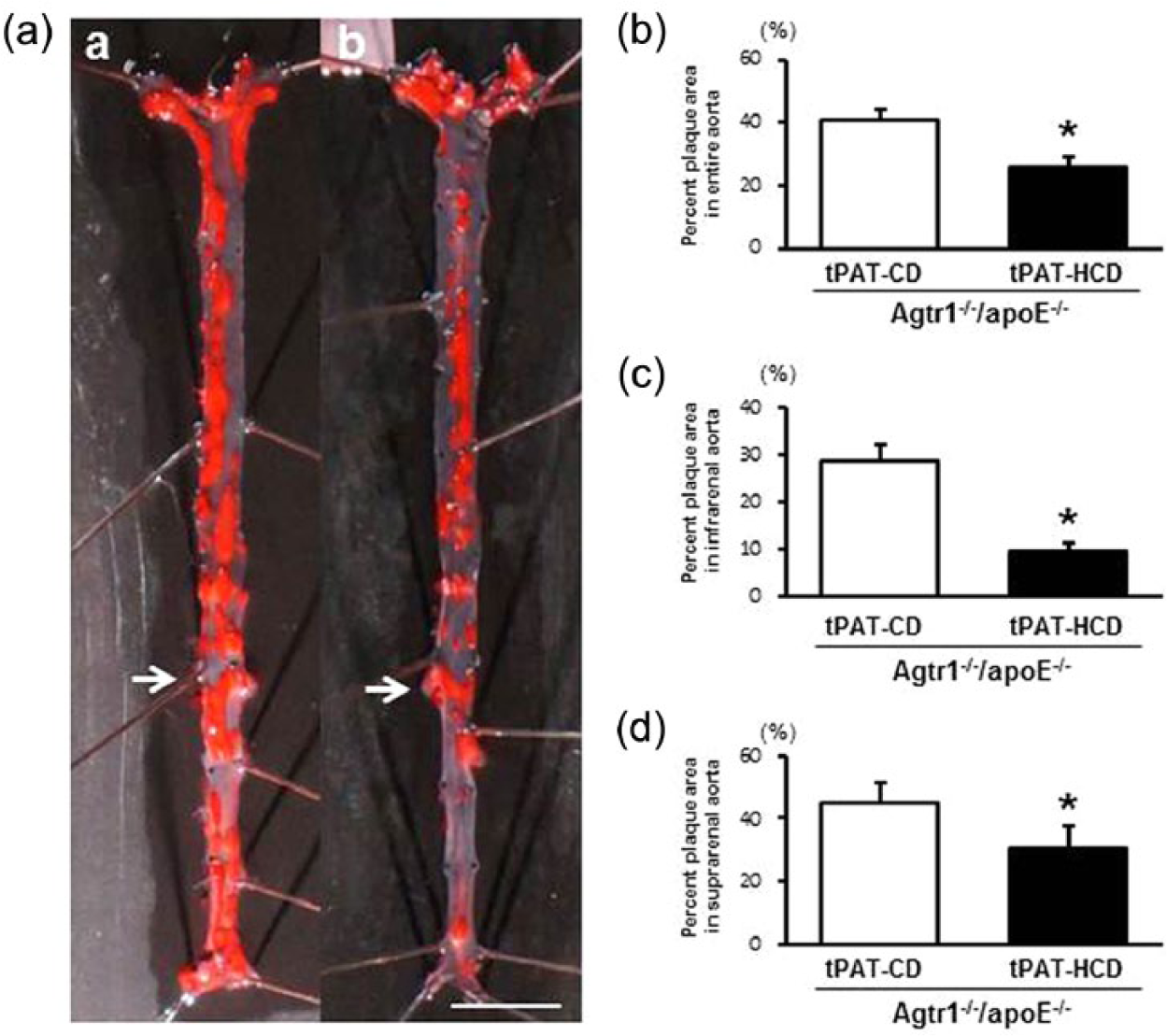
Intra-abdominal transplantation of tPAT from high-cholesterol diet-fed Agtr1–/–/apoE–/– mice ameliorates atherosclerotic development. (a) Representative photographs of en face oil-red O-stained entire aortas in tPAT-CD/Agtr1–/–/apoE–/– (a) and tPAT-HCD/Agtr1–/–/apoE–/– (b). Arrows indicate right renal artery. The scale bar shows 5-mm intervals. (b) Quantitative analysis of the atherosclerotic lesion area in the entire aorta. (c) Quantitative analysis of the atherosclerotic lesion area in the infrarenal aorta. (d) Quantitative analysis of the atherosclerotic lesion area in the suprarenal aorta. Values are the mean ± SE for at least eight mice in each group. *p < 0.05 vs. tPAT-CD/Agtr1–/–/apoE–/–. tPAT-CD/Agtr1–/–/apoE–/–: apoE–/– mice transplanted with the tPAT from chow diet-fed Agtr1–/–/apoE–/– mice. tPAT-HCD/Agtr1–/–/apoE–/–: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed Agtr1–/–/apoE–/– mice; CD: chow diet; HCD: high-cholesterol diet.
Effect of tPAT transplantation on the circulating levels of adipocytokines
To elucidate the mechanisms by which transplanted tPAT influenced the development of atherosclerosis, we measured the serum concentrations of adipocytokines related to atherogenesis. The concentrations of TNF-α, IL-6, and adiponectin in the serum were comparable between tPAT-CD- and tPAT-HCD-transplanted animals. However, TNF-α and IL-6 levels were significantly lower in tPAT-HCD/ARB-transplanted mice compared to tPAT-CD- or tPAT-HCD-transplanted animals (Figures 6(a) and 6(b)). Conversely, the serum concentration of adiponectin in tPAT-HCD/ARB-transplanted animals was significantly higher than that in other groups (Figure 6(c)). These findings suggested that marked reduction in serum proinflammatory adipokine levels and an associated increase in adiponectin concentration are likely responsible for the anti-atherogenic effects observed in tPAT-HCD/ARB-transplanted mice.

Transplantation of the tPAT from ARB-treated apoE–/– mice attenuates the circulating levels of proinflammatory adipocytokines. Circulating levels of adipocytokines in mice showing significantly lower concentrations of TNF-α (a) and IL-6 (b), and a higher concentration of adiponectin (c) four weeks after the transplantation of tPAT-HCD/ARB. Values are the mean ± SE for at least eight mice in each group. *p < 0.05 vs. sham control. #p < 0.05 vs. tPAT-CD. †p < 0.05 vs. tPAT-HCD. tPAT-CD: apoE–/– mice transplanted with the tPAT from chow diet-fed mice. tPAT-HCD: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed mice. tPAT-HCD/ARB: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed mice treated with angiotensin receptor blocker. tPAT: thoracic periaortic adipose tissue; ARB: angiotensin II (AngII) receptor blocker; TNF-α: tumor necrosis factor alpha; IL-6: interleukin 6; HCD: high-cholesterol diet; CD: chow diet; APN: adiponectin.
Transplanted tPAT from ARB-treated apoE–/– mice exerts anti-inflammatory effects in recipient mice
To identify the primary tissue source of proinflammatory cytokines and adiponectin, we examined the transplanted tPAT grafts in recipient mice. tPAT grafts harvested from tPAT-CD and tPAT-HCD transplanted mice showed unilocular fat droplets similar to those of a WAT, whereas a transplanted tPAT graft from tPAT-HCD/ARB showed multilocular fat droplets characteristic of brown adipose tissue (Figure 7(a)). Immunohistochemical analysis revealed that the number of Mac-2-positive cells was significantly lower in tPAT-HCD/ARB than in tPAT-HCD (Figures 7(b) and 7(c)). Consistent with the circulating levels of proinflammatory cytokines, the mRNA expression levels of TNF-α and IL-6 were lower in tPAT-HCD/ARB, whereas adiponectin mRNA expression was similar in all three groups (Figure 7(d)). The protein levels of TNF-α, IL-6, and adiponectin were significantly lower in tPAT-HCD/ARB (Figure 7(e)). In contrast to the tPAT, the morphological characteristics and mRNA expression levels of adipocytokines in the WAT of all three groups were comparable (Supplemental Figures 3(a) and 3(b)). The protein levels of adipocytokines did not differ between tPAT-HCD and tPAT-HCD/ARB (Supplemental Figure 3(c)). These findings suggested that reduced expression of TNF-α and IL-6 in transplanted tPAT, accompanied by a relatively limited accumulation of macrophages, likely led to lower circulating levels of these pro-inflammatory cytokines and contributed to the prevention of atherosclerotic development in tPAT-HCD/ARB-transplanted mice.
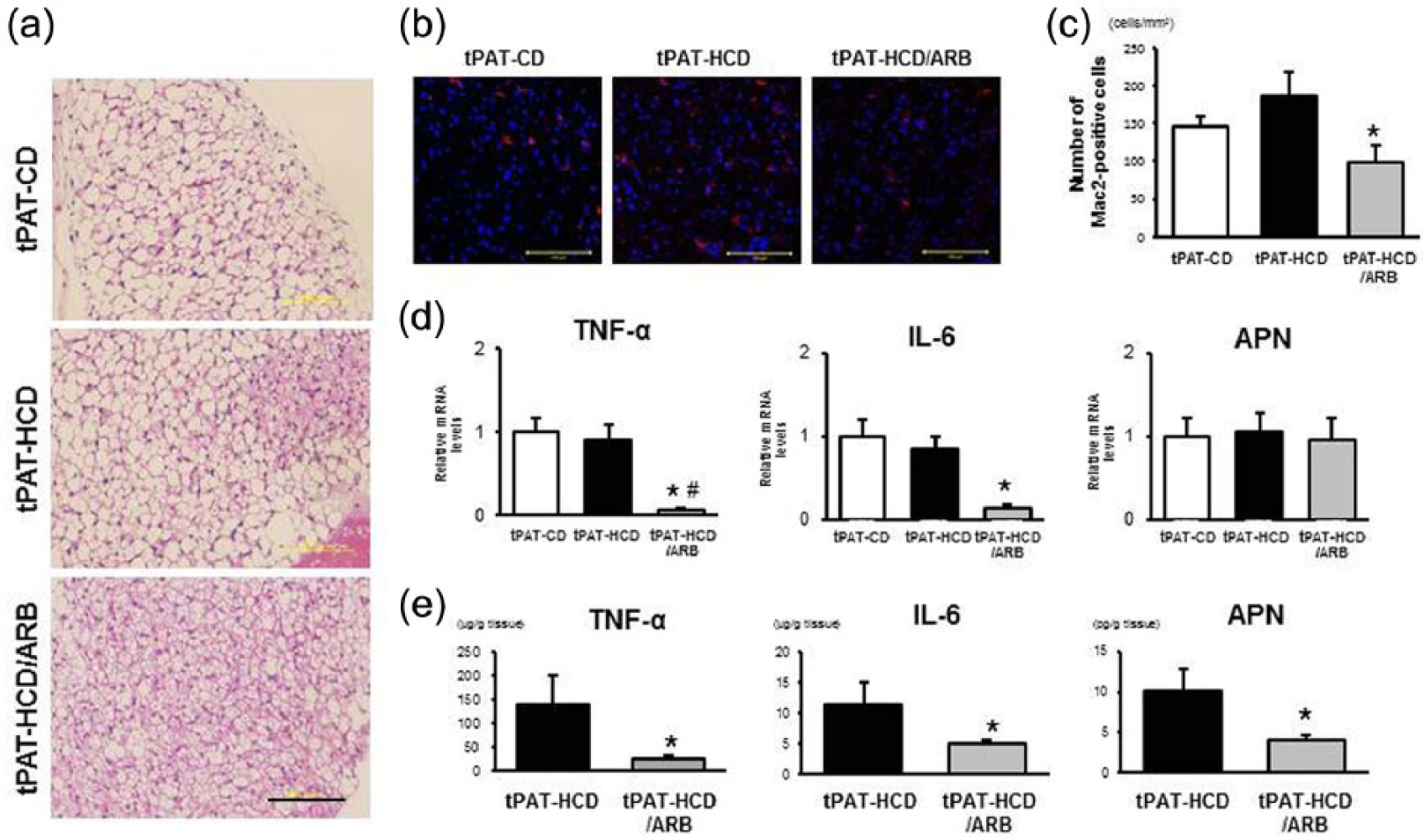
tPAT graft from ARB-treated apoE–/– mice exhibits anti-inflammatory properties. (a) Histomorphological analysis of hematoxylin-eosin staining in tPAT graft. tPAT-CD: apoE–/– mice transplanted with the tPAT from chow diet-fed mice. tPAT-HCD: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed mice. tPAT-HCD/ARB: apoE–/– mice transplanted with the tPAT from high-cholesterol diet-fed mice treated with angiotensin receptor blocker. The scale bar shows 100-µm intervals. (b) and (c) Immunohistochemical staining and quantitative analysis of Mac-2-positive cells. Values are the mean ± SE for at least six mice in each group. *p < 0.05 vs. tPAT-HCD. The scale bar shows 100-µm intervals. (d) Quantitative real-time PCR analysis of the gene expression of TNF-α, IL-6, and adiponectin. Values are the mean ± SE relative to tPAT-CD for at least six mice in each group. *p < 0.05 vs. tPAT-CD, #p < 0.05 vs. tPAT-HCD. APN: adiponectin. (e) Tissue concentrations of TNF-α, IL-6, and adiponectin in tPAT grafts. Values are the mean ± SE for at least five mice in each group. *p < 0.05 vs. tPAT-HCD. tPAT: thoracic periaortic adipose tissue; ARB: angiotensin II (AngII) receptor blocker; TNF-α: tumor necrosis factor alpha; IL-6: interleukin 6; HCD: high-cholesterol diet; CD: chow diet; PCR: polymerase chain reaction; APN: adiponectin.
Discussion
In this study, we demonstrated that an HCD induced tPAT-specific enhanced expression of RAS components and accumulation of monocytes/macrophages, and that such changes were absent in ARB-treated apoE–/– mice as well as Agtr1–/–/apoE–/– mice. Further, intra-abdominal transplantation of the tPAT from HCD-fed apoE–/– mice significantly exaggerated atherosclerotic development throughout the aorta; atherosclerotic development was markedly suppressed in mice transplanted with the tPAT from ARB-treated HCD-fed apoE–/– mice. We also determined that transplantation of the tPAT from HCD-fed Agtr1–/–/apoE–/– mice suppressed atherosclerotic development to a greater extent than transplantation from CD-fed Agtr1–/–/apoE–/– mice. The tPAT grafts from ARB-treated apoE–/– mice expressed lower amounts of proinflammatory cytokines and had fewer monocytes/macrophages. This was associated with significantly lower circulating levels of TNF-α and IL-6, and higher adiponectin levels. Our findings clearly suggest that AT1 receptor-mediated proinflammatory phenotypic alterations of the tPAT are involved in the development of atherosclerosis, and that modulation of tPAT phenotype may be a useful therapeutic strategy to prevent cardiovascular diseases.
It has been reported that the tPAT exhibits a brown adipose tissue-like appearance and the expression profile of adipocyte-related genes.13,18 Recently, Chang et al. developed a smooth muscle cell-specific PPARγ-deficient mouse model, in which absence of tPAT markedly reduced the thermogenic capacity and energy expenditure of excessive nutrition. 16 The authors demonstrated that atherosclerosis development was significantly exaggerated when the animals were housed at 16°C, but not at 22°C, because of the intravascular temperature-associated endothelial dysfunction and impaired lipid clearance. They also showed that mRNA expression levels of uncoupling protein 1 (UCP-1) and peroxisome proliferator-activated receptor-γ coactivator (PGC-1α) were significantly higher in tPAT from control mice housed at 16°C than at 22°C, suggesting that cold-mediated activation of the tPAT exerted anti-atherogenic effects. Therefore, we examined the relationship between tPAT activation and atherogenesis. UCP-1 mRNA expression was significantly higher in tPAT-HCD than in tPAT-CD, whereas such an increase was absent in tPAT-HCD/ARB (Supplemental Figure 4(a)). Similarly, PGC-1α mRNA expression was markedly lower in tPAT-HCD/ARB than in the other groups (Supplemental Figure 4(b)). Considering that mice were housed at 22°C throughout the study, tPAT activation was unlikely to be involved in the tPAT-associated development of atherosclerosis.
It has been established that the AT1 receptor exerts proatherogenic actions, and that pharmacological blockade or a deficiency of the AT1 receptor ameliorates atherosclerotic development.19–21 Adipose tissue is a mixture of various cell types, 22 and adipocytes as well as monocytes/macrophages abundantly express RAS components. 23 Putnam et al. have recently generated adipocyte-specific AT1a receptor-deficient mice that developed striking adipocyte hypertrophy when fed a low-fat diet. 24 The authors showed that incubation of 3T3-L1 adipocytes with Ang II increased oil red O staining and mRNA expression of PPARγ, suggesting that AT1 receptors regulated adipocyte differentiation and morphology. Consistent with their results, we observed that an HCD promoted the expression of PPARγ as well as RAS components, and treatment with ARB suppressed PPARγ mRNA expression (Figures 2(a), 2(c), and 3(a)). However, suppression of adipocyte differentiation by AT1 blockade may increase lipid storage and hypertrophy of remaining adipocytes, and promote the secretion of pro-inflammatory cytokines/chemokines, leading to increased accumulation of inflammatory leukocytes in adipose tissue.25,26 Therefore, future studies using adipocyte-specific AT1 receptor-deficient atherosclerosis-prone mice are needed to elucidate the role of adipocyte AT1 in atherogenesis. In contrast, HCD feeding led to increased monocytes/macrophages accumulation in the tPAT, which was blocked by ARB treatment (Figures 2(e) and 3(e), Supplemental Figures 5(a) and 5(b)). We have previously reported that bone marrow AT1 receptors play a crucial role in monocyte differentiation and/or proliferation. 27 Considering that monocytes/macrophages abundantly express all components of RAS 23 and may serve as a predominant source of pro-inflammatory cytokines/chemokines in adipose tissue,25,26 AT1 receptors expressed in these cells likely play an important role in HDC-induced phenotypic alterations in the tPAT and atherosclerotic development.
Visceral adipose tissue has been reported to release several pro-inflammatory cytokines into circulation, causing systemic inflammation and contributing to the development of metabolic disorders such as insulin resistance.1–5 Clinical studies have shown that a circulating concentration of pro-inflammatory cytokines is well correlated with the incidence of atherosclerotic cardiovascular diseases.28,29 Studies using animal models also demonstrate the role of pro-inflammatory cytokines on atherosclerotic development.30–32 We examined whether tPAT-derived adipocytokines modulated the development of atherosclerosis in recipient mice. Unexpectedly, there was no difference in the circulating levels of TNF-α, IL-6, and adiponectin between tPAT-CD- and tPAT-HCD-transplanted mice. The precise mechanism responsible for the exaggerated atherosclerosis observed in tPAT-HCD-transplanted mice is not clear. However, tPAT from ARB-treated mice exerted an inhibitory effect on atherosclerotic development. tPAT-HCD/ARB transplanted mice exhibited higher concentrations of circulating adiponectin and HDL cholesterol (Figure 6(c) and Supplemental Figure 2(b)). Pro-inflammatory cytokines have been reported to inhibit adiponectin secretion.33,34 Our results suggested that transplantation tPAT from tPAT-HCD/ARB exerted a beneficial effect on lipid homeostasis by suppressing systemic inflammation.
We examined the circulating cytokine levels in the recipient mice transplanted with AT1-deficient tPAT; however, serum concentrations of TNF-α and IL-6 did not differ between tPAT-CD/Agtr1–/–/apoE–/– and tPAT-HCD/Agtr1–/–/apoE–/– mice (Supplemental Figures 6(a) and 6(d)). Cytokine levels in transplanted tPAT graft also showed no difference between the two groups (Supplemental Figures 6(b) and 6(e)). Considering the discrepancy between the data from tPAT-HCD/ARB apoE–/– mice and tPAT-HCD/Agtr1–/–/apoE–/– mice, we cannot completely exclude the possibility that residual ARB within the transplanted tPAT could act as a source of ARBs in the recipient animal. Conversely, proinflammatory cytokine levels in epididymal WAT was markedly decreased in tPAT-HCD/Agtr1–/–/apoE–/– mice compared with tPAT-CD/Agtr1–/–/apoE–/– mice (Supplemental Figures 6(c) and 6(f)), suggesting that tPAT graft from tPAT-HCD/Agtr1–/–/apoE— mice could exert an anti-inflammatory effect on endogenous adipose tissue. Potential mediators and underlying mechanisms responsible for these effects need to be investigated in a future study.
In this study, HCD-induced tPAT phenotypic alteration was examined in apoE–/– mice; however, HFD-induced tPAT modulation in wild-type mice was a quite important issue, especially in the setting of obesity. We also examined HFD (but not HCD)-induced phenotypic alteration of tPAT in wild-type mice and found that macrophage accumulation and gene expression of the RAS components were exaggerated in tPAT of obese wild-type mice compared with lean control mice (data not shown). Their relevance to atherosclerosis development needs to be examined in a future study using a tPAT transplant model.
In conclusion, our results showed that periaortic adipose tissue-specific activation of RAS is involved in the atherosclerotic development in HCD-fed apoE–/– mice, and that inhibition of this depot-specific activation of RAS exerts an anti-atherogenic effect by suppressing systemic inflammation. Our findings shed new light into the role of tPAT in atherogenesis, and suggest that perivascular adipose tissue may be a useful new therapeutic target for the prevention of cardiovascular diseases.
Footnotes
Conflict of interest
None declared.
Funding
This work was supported by a grant from the Ministry of Education, Culture, Sports, Science and Technology of Japan (C: KAKENHI-24591120).