
Abstract
Background:
In the present study, we investigated the potential effect of aliskiren on smooth muscle cell (SMC) migration in response to prorenin.
Methods:
Cultured human SMCs were incubated with angiotensinogen (ANG) (1.5 × 10−7M) and increasing concentrations of aliskiren (10−6−10−5M). After 24 h, SMC migration was assessed by Boyden’s chamber chemotactic assay using prorenin as chemotactic factor (10−8M). The effect of aliskiren on RhoA and Rac activity was also determined by G-LISA assay and the lamellipodia formation by rhodamine–phalloidin staining. Changes in cell morphology were recorded in real-time using the iCelligence system.
Results:
Aliskiren determined, at 10−5M, a significant inhibition of SMC migration induced by prorenin (−66.4±18.1%; p < 0.05), while no significant effect was observed when PDGF-BB was utilized as chemotactic agent. Aliskiren also reduced Rac-GTP levels in response to prorenin (−54.2±5.4%) without affecting the RhoA-GTP levels. Finally, aliskiren inhibited both the lamellipodia formation and morphological changes induced by prorenin with no significant effect on PDGF-BB activity.
Conclusions:
Taken together, we provide the first evidence of the inhibitory action of aliskiren on SMC migration induced by prorenin.
Introduction
In advanced atherosclerotic lesions, smooth muscle cell (SMC) accumulation in the intima results from cell proliferation and directed migration of SMCs from the media into the intima. 1 These processes are regulated by various cytokines and chemokines through the activation of their receptors expressed on the cell surface of vascular SMCs. 2 In particular, the discovery of the prorenin receptor (PRR) expressed in vascular SMC has opened to a completely new scenario on the functional role of local renin–angiotensin–aldosterone system (RAAS) on atherosclerosis.3,4 Pioneer studies have reported the presence of all the components of the classical RAAS pathway in atherosclerotic lesions, implying its potential contribution on SMC accumulation in the atherosclerotic plaque.5–8
PRR can be activated by either renin or prorenin; 9 however, prorenin appears to be the most relevant agonist since its plasma concentrations are approximately tenfold higher than those of renin and the binding affinity is threefold higher than renin.10,11 The activation of PRR has been shown to drive cell cycle progression, DNA synthesis, ERK1(p44) and ERK2(p42) phosphorylation,12–14 and p38 MAP kinase/HSP27 activation, 15 all events associated with cell proliferation. Moreover, we have recently provided evidence that prorenin, through the activation of PRR, induces SMC migration. 16 It is therefore conceivable to hypothesize that PRR activation may promote both SMCs proliferation and migration, pivotal features of atherogenesis. 17
Aliskiren represents the first direct renin inhibitor to be approved for clinical use in hypertension that blocks the RAAS at its point of origin,18,19 the renin–ANG interaction, thus preventing the conversion of angiotensinogen (ANG) to angiotensin I and interrupting the RAAS cascade. The renin inhibition by aliskiren determines a significant increase of renin and prorenin plasma concentrations, 20 and thus potentially the activity of PRR in vascular SMCs.
From the crystal structure modeling and the in vitro results conducted with cells overexpressing PRR, it was demonstrated that aliskiren does not interfere with the binding of renin and prorenin to the PRR.13,21 However, in a model of hypertensive diabetic renal damage, renin inhibition with aliskiren suppressed renal gene expression of the PRR and its target genes transforming growth factor-β (TGF-β) and type I collagen. 22 A similar inhibitory effect of aliskiren on PRR has been observed in human aortic SMCs incubated with both prorenin and ANG. 23 Such effect determined a significant inhibition of the transcription of three genes, previously shown to be regulated by prorenin, such as TGF-β, type I collagen and plasminogen activator inhibitor-1 (PAI-1).3,24 Thus, aliskiren inhibits the in vivo RAAS activity by binding to the active site of circulating renin or the unmasked catalytic site of prorenin bound to PRR and by reducing the PRR expression.4,22,23 Although the patho-physiological relevance of the PRR down-regulation by aliskiren still needs to be fully elucidated, results from in vivo experimental studies suggest that, to some extent, the anti-atherosclerotic properties of aliskiren are independent from the hypotension effect.25–28
Based on these premises, in the present study we tested the hypothesis that aliskiren may interfere with the prorenin-dependent chemotactic effect on human aortic SMCs.
Material and methods
Cell culture
Human aortic SMCs were purchased from PromoCell and cultured in Smooth Muscle Cell Growth Medium 2 supplemented with 5%FCS, 0.5 ng/mL of epidermal growth factor, 2 ng/mL of basic fibroblast growth factor, and 5 µg/mL of insulin. SMCs were utilized between passage 5 and 9. ANG and prorenin were purchased from Vinci-Biochem.
Experimental protocols
For the determination of cell migration, SMCs were seeded in complete medium at the density of 2 × 105 SMCs per 35 mm petri dish. After 24 h, cells were incubated in Dulbecco’s modified Eagle’s medium (DMEM) containing 0.4% FCS, ANG (1.5 × 10−7M) and increasing concentrations of aliskiren (10−6−10−5M). After an additional 24 h, SMCs were harvested by trypsinization, resuspended in their conditioned media and the migration determined by Boyden’s chamber chemotactic assay with prorenin (10−8M) or platelet-derived growth factor-BB (PDGF-BB) (10−9M) as chemotactic agents for 6–8 h. Under the same experimental conditions, the lamellipodia formation were assessed by rhodamine-phalloidin staining after 1 h stimulation, while for G-LISA assay cells were stimulated for 10 minutes. Finally, changes of cell morphology were monitored every minute for 6 h after the addition of prorenin or PDGF-BB by iCelligence system.
Cell migration assay
The Boyden chamber and the polycarbonate membrane were purchased from Biomap (Agrate Brianza, Milan Italy). The membranes were coated with a 0.1 mg/mL of type I collagen solution (PureCol®, Nutacon BV, Leimuiden, The Netherlands) in 0.1M acetic acid at 37°C. The lower compartments of the wells of a modified Boyden chamber were filled (in triplicate) with DMEM containing 0.4% FCS in the absence or presence of prorenin or PDGF-BB, whereas the upper compartments were filled with SMCs suspensions containing 106 cells/mL. The cells were resuspended with their conditioned media containing ANG and increasing concentrations of aliskiren. The chamber was incubated at 37°C for 6 h. The membrane was then carefully removed. Adherent SMCs on the top were eliminated and the membrane was stained with the Diff-Quik staining set (Biomap, Agrate Brianza, Milan Italy). The number of transmigrated cells was counted in four random high-power fields (HPFs) under high magnification (objective lens 20×). 29
Cytoskeleton staining
Cells were fixed in 4% paraformaldehyde at room temperature for 10 min, permeabilized in 0.1% Triton X-100 in phosphate-buffered saline for 15 min, and incubated with rhodamine-phalloidin (Sigma Aldrich, Milan, Italy). Cytoskeleton staining of cells was analyzed using a fluorescence microscope (Axiovert M220; Zeiss Instruments).
Real-time monitoring of cell morphology using the iCELLigence system
The iCELLigence system (ACEA Biosciences Inc, San Diego, CA, USA) monitors cellular events in real time by recording the electrical impedance that is correlated with cell number, morphology and viability in a given culture well. For analyzing SMCs morphology, 40.000 cells/well were plated into the E-Plate L8, and the extent of cell spreading was monitored every minute for 6 h with iCELLigence. The assay system expresses impedance in arbitrary cell index units (Rn − Rb)/4.6, where Rn is the cell–electrode impedance of the well when it contains cells and Rb is the background impedance of the well with the media alone.
G-LISA assay for Rac and RhoA
The intracellular amount of Rac-GTP and RhoA-GTP were determined by using the G-LISA assay according to the manufacturer’s instruction (Cytoskeleton Inc, Denver, CO, USA). 30
Statistical analysis
All data shown are representative of at least three replicate experiments. Data are expressed as mean±SD. Statistical analyses were performed using either the unpaired student’s t test or 1-way ANOVA as appropriate; p values <0.05 were considered significant.
Results
In a first series of experiments, the effect of aliskiren on SMC migration was determined by Boyden’s chamber chemotactic assay. As previously reported, 16 prorenin induced SMC migration (2.2±0.3 fold; p < 0.05) and the pre-incubation with 10−5M aliskiren for 24 h determined a significant inhibition of the chemotactic effect (−66.4±18.1%) (Figure 1). The prorenin-mediated SMC migration has been shown to be associated with the activation of both RhoA and Rac small GTPase proteins. 16 Thus the effect of aliskiren on the intracellular levels of Rac-GTP and RhoA-GTP was investigated by G-LISA assay. As shown in Figure 2, prorenin increased both RhoA-GTP and Rac-GTP levels by 39.0±1.4% and 38.0±5.0%, respectively. The pre-incubation with 10−5M aliskiren inhibited the activation of Rac (−54.2±5.4%; p < 0.05) while no effect was observed on RhoA-GTP levels (Figure 2).
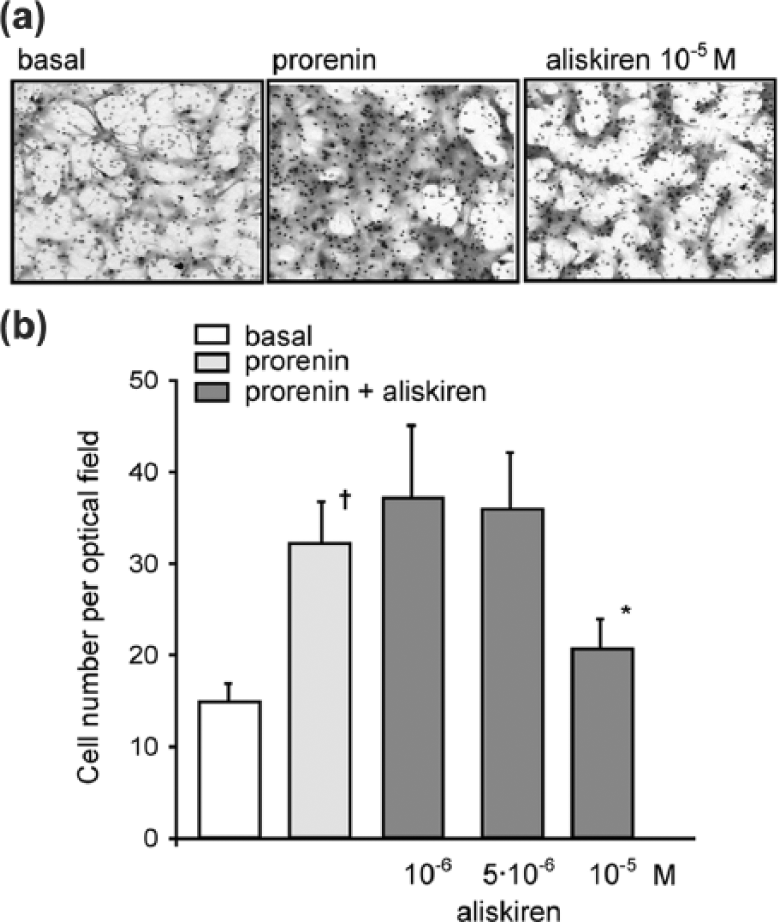
Aliskiren inhibits human SMC migration in response to prorenin. Human SMCs were cultured for 24 h in Smooth Muscle Cell Growth Medium 2 (50.000 cells per well/48 well tray). After 24 h, medium was changed with DMEM/0.4%FCS, ANG (1.5 × 10−7M) in the presence or absence of aliskiren (10−6M, 5 × 10−6M, and 10−5M). After 24 h, cells were harvested by trypsinization and the migration measured by Boyden’s chamber chemotactic assay by using prorenin (10−8M) as chemotactic agent. (a) Representative images of migrated SMCs stained with the Diff-Quik staining kit. (b) The number of transmigrated cells was counted in four random high-power fields (HPFs) under high magnification (objective lens 20×). Basal indicates cells incubated in DMEM/0.4%FCS and ANG (1.5 × 10−7M) and not stimulated with prorenin. †p < 0.05 prorenin vs. basal; *p < 0.05 aliskiren vs. prorenin. The results are representative of five independent experiments.
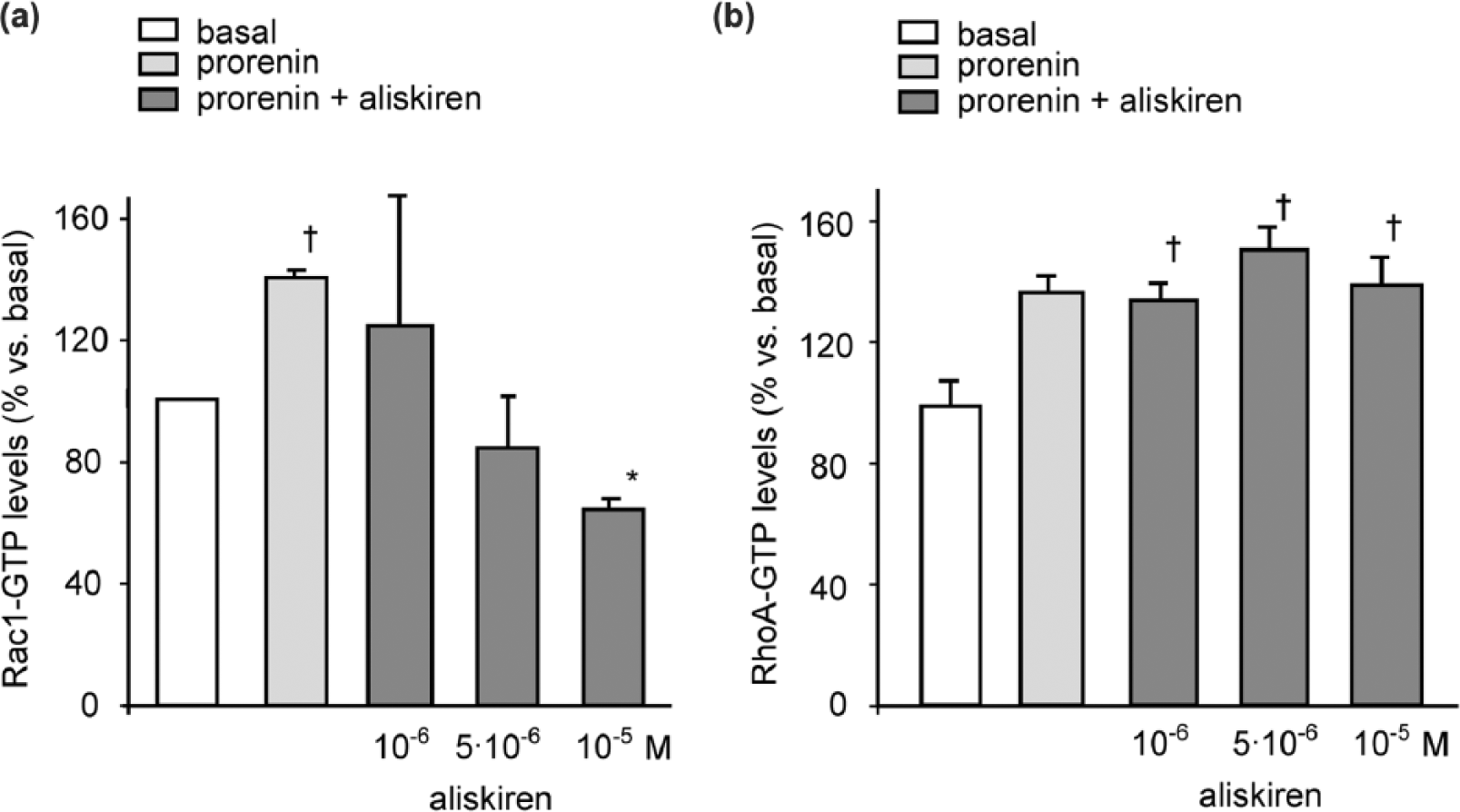
Rac-GTP levels are reduced in human SMCs treated with aliskiren after stimulation with prorenin. Human SMCs were cultured for 24 h in Smooth Muscle Cell Growth Medium 2 (50.000 cells per well/48 well tray). After 24 h, medium was changed with DMEM/0.4%FCS, ANG (1.5 × 10−7M) in the presence or absence of aliskiren (10−6M, 5 × 10−6M, and 10−5 M). After 24 h, cells were stimulated with prorenin (10−8M) for 10 minutes and G-LISA assay for Rac-GTP (a) and RhoA-GTP (b) performed from the total protein extracts. Basal indicates cells incubated in DMEM/0.4%FCS and ANG (1.5 × 10−7 M) and not stimulated with prorenin. †p < 0.05 prorenin vs. basal; *p < 0.05 aliskiren vs. prorenin. The results are representative of three independent experiments.
In order to investigate whether the effect of aliskiren on SMC migration was specific to the prorenin stimulation, a series of Boyden’s chamber chemotactic assays using PDGF-BB as chemotactic agent was performed. SMCs were incubated for 24 h in DMEM containing 0.4%FCS and ANG in the presence or absence of increasing concentrations of aliskiren and then divided in two aliquots one for the migration towards prorenin and one towards PDGF-BB. Interestingly, aliskiren significantly affected the SMC migration induced by prorenin and no effect was observed with PDGF-BB (Figure 3). These data suggest that aliskiren selectively interferes with the prorenin mediated chemotactic action potentially by inhibiting Rac activation.

Aliskiren does not affect human SMC migration in response to PDGF-BB. The experimental conditions are the same as in Figure 2. The Boyden’s chamber chemotactic assay was performed by using prorenin (10−8M) and PDGF-BB (10−9M) as chemotactic agents. †p < 0.05 prorenin and PDGF-BB vs. basal; *p < 0.05 aliskiren vs. prorenin. The results are representative of three independent experiments.
The effect of aliskiren on lamellipodia formation was examined by rhodamine-phalloidin staining of filamentous actin (F-actin). After 1 h stimulation of SMCs with prorenin or PDGF-BB, a significant redistribution of F-actin was observed, resulting in the production of lamellipodia and membrane protrusions (Figure 4). The pre-incubation of 10−5M aliskiren prior cell stimulation, significantly affected the lamellipodia formation in response to prorenin while no effect was observed with PDGF-BB (Figure 4).
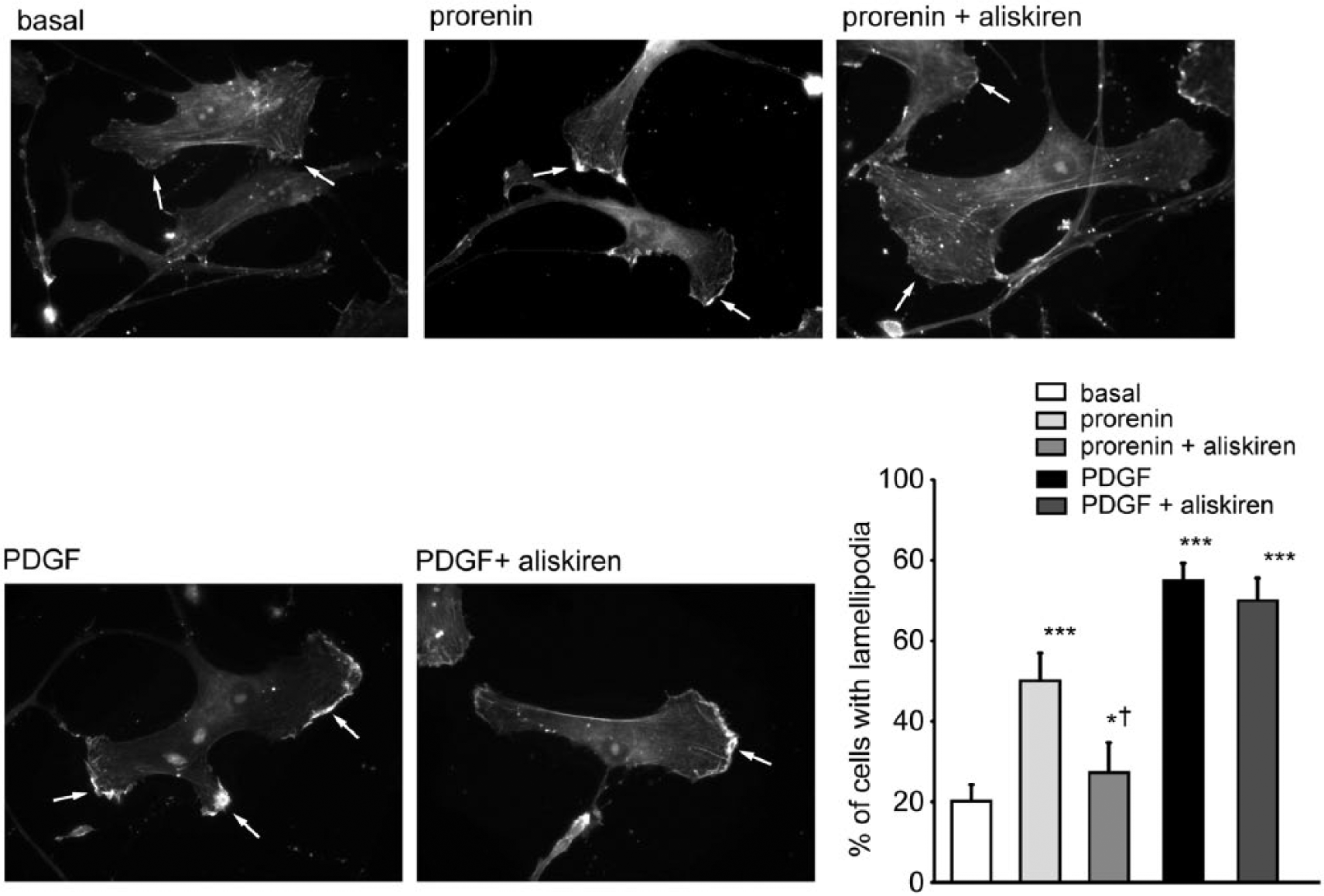
Effect of aliskiren on lamellipodia formation in human SMCs. Human SMCs were cultured for 24 h in Smooth Muscle Cell Growth Medium 2 (40.000 cells per well/24 well tray). After 24 h, media was changed with DMEM/0.4%FCS, ANG (1.5 × 10−7M) in the presence or absence of aliskiren (10−5M). After 24 h, cells were stimulated with prorenin (10−8M) and PDGF-BB (10−9M) for 1 h then fixed and the cytoskeleton stained with rhodamine–phalloidin. Basal indicates cells incubated in DMEM/0.4%FCS and ANG (1.5 × 10−7M) and not stimulated with prorenin. Arrows indicate F-actin localization in lamellipodia. Results are representative of three independent experiments. The histogram reports the number of cells with lamellipodia counted in three random high-power fields (HPFs) from three replicates under high magnification (objective lens 20×). ***p < 0.01; *p < 0.05 prorenin vs. basal; aliskiren vs. prorenin. †p < 0.05 prorenin + aliskiren vs. prorenin.
In order to better define the effect of aliskiren on cell morphology a real-time recording iCelligence system was utilized. The major advantages of this system is that the changes in cell morphology are monitored in real time and reported as average of 80.000 cells. As shown in Figure 5, prorenin induced significant changes in cell morphology as indicated by an increased cell index during the first 3 h of stimulation reaching a plateau that was sustained up to 6 h. The presence of 10−5M aliskiren significantly reduces the cell index values by approximately 40% during all the duration of the recorded time. Differently from prorenin, PDGF-BB induced a strong and transient increase of cell index with a peak at 1h and the presence of aliskiren did not affect this response. Taken together, aliskiren significantly affected the SMC migration induced by prorenin affecting the lamellipodia formation and changes in cell morphology with no effect on PDGF-BB.

Effect of aliskiren on time-dependent morphological change induced by prorenin and PDGF-BB. Human SMCs were cultured for 24 h in Smooth Muscle Cell Growth Medium 2 (40.000 cells per well of E-Plate L8). After 24 h, media was changed with DMEM/0.4%FCS, ANG (1.5 × 10−7M) in the presence or absence of aliskiren (10−5M). After 24 h, cells were stimulated with prorenin (10−8M) and PDGF-BB (10−9M) and cell index monitored in real time every minute for 6 h. Basal indicates cells incubated in DMEM/0.4%FCS and ANG (1.5·10−7 M). Results are representative of three independent experiments.
Discussion
In the present report we have provided evidence that the direct renin inhibitor, aliskiren, interferes with the SMC migration induced by prorenin. Recently, our group has demonstrated that PRR mediates the chemotactic effect of prorenin on SMCs and this action is mainly mediated by the activation of the small GTPases Rac and RhoA. 16 In addition, we and others, have documented that aliskiren reduces the PRR expression in vivo and in vitro,22,23 which, in turn, determines an inhibitory action on PRR activity.22,23 Indeed, the effect of aliskiren on PRR expression levels in human SMCs was shown to be sufficient to inhibit the transcription of at least three target genes of prorenin, such as TGF-β, PAI-1, and Type I collagen22,23 In the present report, we extend these observations, reporting the inhibitory effect of aliskiren on Rac activation in response to prorenin and SMC migration. The effect of aliskiren on cell migration was demonstrated by the Boyden’s chamber chemotactic assay and confirmed by the real-time monitoring of cell morphology by iCelligence system and the analysis of lamellipodia formation. All these events were shown to be significantly inhibited by aliskiren, at the concentration of 10−5M, and only when SMCs were stimulated with prorenin but not in response to PDGF-BB. Thus, aliskiren appears to have a selective action on prorenin-induced SMC migration.
Several lines of evidence indicate that Rac plays an important role on SMC migration, associated with the development of atherosclerotic plaque, either by inducing the lamellipodia formation or by generating reactive oxygen species (ROS) by NADPH oxidase.31,32 Our results suggest that the inhibitory effect of aliskiren on Rac activity may be responsible for the reduction of SMC migration mediated by prorenin. Indeed, we have previously demonstrated that the inhibition of Rac by siRNA completely abrogated the chemotactic effect of prorenin on SMCs. 16 Nevertheless, the actual inhibitory mechanism of action of aliskiren on Rac activity is still unknown and further investigations are required in order to determine if aliskiren affects the post-translational modifications of Rac, such as geranylgeranylation, or the GDP/GTP exchange processes. Indeed, previous studies have established the requirement of protein geranylgeranylation of the CAAX motif of Rac and RhoA for their interaction with the plasma membrane and with the GDP/GTP exchange proteins.33–36 In addition, it will be interesting to investigate the effect of aliskiren on Rac-mediated ROS formation and its contribution on the hypertension-associated end-organ damage. 37 On this matter it is important to mention that, recently, aliskiren has been reported to reduce vascular NADPH oxidase activity by 41.6% in the aortic tissue of ApoE-deficient mice. 38 This evidence further supports a potential inhibitory effect of aliskiren on Rac.
Interestingly, aliskiren does not reduce the intracellular levels of RhoA-GTP in SMC stimulated with prorenin. The differential action of aliskiren on Rac and RhoA could be related to the effect of angiotensin I receptor (AT1R) and PRR on the two small GTPases. Indeed, while the PRR-mediated activation of both small GTPases has only been recently observed by our group, 16 the role of the AT1 receptor and angiotensin II in vascular SMCs is well established.31,39,40 Under our experimental conditions, the addition of prorenin, in the presence of ANG, determines a significant increase of angiotensin II formation, 23 and thus the stimulation of both receptors, PRR and AT1R. Since aliskiren was shown to fully inhibit the conversion of ANG to angiotensin I, and only partially reducing PRR expression, 23 the differential effect on the two pathways may account for the selective action on Rac.
Our finding on the effect of aliskiren on cell migration, although obtained in an in vitro cultured system, may contribute to better understand the pharmacological properties of aliskiren. The demonstration that aliskiren can accumulate in the arterial wall of the small cortical vessels in the kidney, 22 suggests a possible inhibition of RAAS in the vasculature and PRR expression in vascular SMCs. The relevance of our data are therefore reinforced by these observations suggesting a possible in vivo pharmacological action of aliskiren on prorenin-mediated SMC migration.
Conclusions
Taken together, in the present study, we have shown that aliskiren inhibits prorenin-mediated SMC migration. Additional studies are needed to determine the in vivo contribution of this pharmacological action in experimental models of cardiovascular diseases. 31
Footnotes
Conflict of interest
Dr. Alberto Corsini has received fees for consulting and research funding from Novartis Farma S.p.A. and Dr. Giuseppe Maiocchi is employee by Novartis Farma S.p.A.
Funding
The present work was supported by a grant from Novartis Farma S.p.A., Origgio, Italy.