
Abstract
Introduction:
Isolated nuclei of sheep proximal tubules express angiotensin (Ang) receptors as well as angiotensinogen (AGT) and renin. The present study characterized the NRK-52E tubular epithelial cell line for the intracellular expression of renin-angiotensin system (RAS) components.
Methods:
RAS components were visualized by immunofluorescent staining in intact cells and protein expression in isolated nuclei.
Results:
An antibody to the angiotensin I (Ang I) sequence of AGT (AI-AGT) revealed only cytosolic staining, while an antibody to an internal sequence of AGT (Int-AGT) revealed primarily nuclear staining. Immunoblots of nuclear and cytosolic fractions confirmed the differential cell staining of AGT. Immunostaining for renin was present on nuclei of intact cells. Nuclear renin activity averaged 0.77±0.05 nmol/mg protein/h that was reduced by aliskiren (0.13±0.01 nmol/mg/h, n=3, p<0.01); trypsin activation increased activity three-fold. Peptide staining localized angiotensin II (Ang II) and Ang-(1-7) to the nucleus and peptide content averaged 59±2 and 57±22 fmol/mg (n=4), respectively. Peptide metabolism in isolated nuclei revealed the processing of Ang I to Ang-(1-7) by thimet oligopeptidase.
Conclusion:
We conclude that the NRK-52E cells express an intracellular RAS localized to the nucleus and may be an appropriate cell model to elucidate the functional relevance of this system.
Introduction
The renin-angiotensin system (RAS) is an endocrine system that plays a major role in the physiological regulation of blood pressure and fluid homeostasis. Dysregulation of the RAS is also thought to contribute to the development and progression of cardiovascular and renal injuries. Moreover, the pharmacological agents that block various components of the RAS now encompass the primary therapies for treatment of hypertension, heart failure and diabetic renal injury. Although originally identified as a classic endocrine system, the evidence clearly reveals a local or tissue RAS in various organs including the kidney, heart, adrenals and brain.1–3 In this regard, an intracellular system localized to cellular organelles including the nucleus and mitochondria have been described in both the tubular epithelial and mesangial cells of the kidney, as well as the myocytes and fibroblasts of the heart.4–11 The physiological relevance and the regulation of this intracellular RAS have not been established. Indeed, it is not clear precisely how the intracellular system functions at the cellular level to synthesize the active RAS peptides angiotensin II (Ang II) and Ang-(1-7), nor the contribution of these peptides to intracellular signaling and cell function. Moreover, there is now compelling evidence for a functional renin receptor that binds prorenin to non-proteolytically activate the enzyme, as well as to mediate the functional signaling of the receptor that is not dependent on Ang II generation.12–14 Interestingly, the prorenin receptor (PRR) is primarily localized intracellularly rather than on the cell membrane suggesting that the receptor may also contribute to a functional intracellular RAS. 15
Elucidation of the physiological relevance of an intracellular RAS is important clinically as well. The current therapeutic regimens to treat high blood pressure and attenuate renal and cardiovascular injury include AT1 receptor antagonists (ARBs), angiotensin converting enzyme (ACE) and renin inhibitors; however, the therapeutic benefit of these agents to target the intracellular system is not known. Previous studies in our laboratory and others have demonstrated a high density of Ang II receptors on isolated nuclei from the kidney.16,17 In the rat kidney, the majority of these nuclear binding sites are the AT1 subtype that is functionally linked to Ang II-dependent increases in oxidative stress and calcium.18,19 AT1 receptor-dependent formation of reactive oxygen species (ROS) was also demonstrated in isolated nuclei from the sheep kidney; however, both AT2 and Ang-(1-7) (AT7) sites were functionally coupled to nitric oxide formation and may antagonize the actions of the nuclear AT1 receptor.20–22 Additional studies revealed the precursor components angiotensinogen (AGT) and renin in isolated nuclei from proximal tubules of the sheep kidney that may portend for the intracellular or nuclear formation of Ang II and Ang-(1-7). 21 Moreover, we detected the peptidase activities for ACE and ACE2 in intact nuclei that processed exogenous Ang I to Ang II and Ang II to Ang-(1-7), respectively. 20 To facilitate our understanding of the tubular RAS within the kidney, the current study sought to identify a renal cell line that expresses the components of this system and to determine their intracellular localization.
Materials and methods
Cell culture
Normal kidney proximal epithelial cells (NRK-52E)
Nuclear and cytosolic fractions
Confluent cells were washed twice with Phosphate buffered saline (PBS) and harvested. Harvested cells were centrifuged at 1000 g for 10 min and the resulting cell pellet was homogenized on ice with a glass pestle in 20 mM Tris buffer containing 5mM MgCl2 and 25 mM KCl at pH 7.8. Homogenates were centrifuged at 1000 g to obtain the crude nuclear pellet and further purified in a high sucrose buffer. The crude pellet was reconstituted with 0.3 M sucrose in 10 mM HEPES buffer pH 7.4, layered over the 0.88 M sucrose buffer and centrifuged at 1200 g for 10 min at 4○C to obtain the nuclear pellet. 23 The cell supernatant was centrifuged at 100,000 g for 60 min at 4○C to obtain the cytosolic fraction.
Western blotting
Nuclear and cytosolic pellets (~50 μg) were boiled in PBS (pH 7.4), diluted in Laemmli buffer with β-mercaptoethanol, separated on 10% Sodium dodecyl sulfate (SDS) polyacrylamide gels for 1 h at 120V in Tris-glycine SDS and transferred to a polyvinylidene difluoride (PVDF) membrane. Blots were blocked with 5% Bio-Rad Dry Milk (Bio-Rad, Hercules, CA, USA) and Tris buffered saline (TBS) with Tween and probed overnight at 4°C with primary antibodies against the following: rat AGT (internal sequence of AGT (Int-AGT), residues 42–57, 1:1000), Ang I sequence of AGT (AI-AGT, residues 25–34, 1:1000) and renin (Inagami antibody no. 826, 1:5000). Membranes were treated with Horseradish peroxidase (HRP)-labeled polyclonal anti rabbit secondary antibodies (1:5000) for 1 h and detected with chemiluminescent substrates (Pierce Biotechnology, Rockford, Illinois, USA). Membranes were probed with mouse monoclonal anti-β-actin (Sigma, St. Louis, Missouri, USA, 1:5000) antibody as a loading control and bands were quantified using MCID densitometry software (InterFocus Imaging, Linton, UK).
Immunofluorescent microscopy
NRK-52E cells were grown in eight chamber slides for two days in DMEM/F12 5% FBS which was replaced with serum free media for one day. Cells were washed with PBS and fixed with 2% paraformaldehyde for 15 min. Following a PBS rinse, cells were permeabilized with 0.2% Triton and then blocked with 3% bovine serum albumin (BSA) (Sigma A-8022). The fixed cells were probed with primary antibodies for AGT and renin previously described for western blot analysis. Cells were also probed with affinity purified antibodies to Ang II (Phoenix Pharmaceutical, Burlingame, California,USA) and Ang-(1-7) (custom antibody), PRR (Abcam ab64957, Cambridge, UK)) antibodies. Antibodies were diluted in 3% normal donkey serum as follows: Int-AGT (1:100), AI-AGT (1:100), renin (1:500), PRR (1:500), Ang II (1:500) Ang-(1-7) (1:25), calnexin (1:100). After overnight incubation with the primary antibody at 4°C, cells were rinsed with PBS, incubated with fluorescent anti-rabbit Alexa Fluor 488 secondary antibody (Invitrogen, Carlsbad, California, USA), and the slides mounted with Molecular Probes ProLong mounting media with 4’,6-diamidino-2-phenylindole (DAPI) (Invitrogen) to stain the nuclei.
Peptide assays
Cell nuclei were isolated as described above and stored at −80°C. The nuclear pellet was reconstituted in MilliQ water and placed in a boiling water bath for 15 min. The nuclear fraction was acidified with trifluoroacetic acid (TFA) to a final concentration of 0.2%, sonicated and centrifuged at 20,000 g for 20 min at 4°C. The resultant supernatant was applied to an activated Sep-Pak C18 extraction column, washed with 0.2% TFA, and the peptide fraction eluted with 3 ml of 80% methanol in 0.2% TFA. Measurement of immunoreactive Ang II and Ang-(1-7) in the extracted nuclei was performed using two distinct radioimmunoassays (RIAs). 24 The Ang-(1-7) RIA fully recognizes Ang-(1-7) and Ang-(2-7), but cross-reacts less than 0.01% with Ang-(3–7), Ang II, Ang I, and their fragments. The Ang II RIA equally recognizes Ang III, Ang-(3-8), and Ang-(4-8), but cross-reacts less than 0.01% with Ang I and Ang-(1-7). The limit of detection was 4 fmol/ tube for Ang-(1-7) and 0.5 fmol/ tube for Ang II.
Renin assay
The cell nuclei were isolated as described above and stored at −80°C. The nuclear pellet was reconstituted in 2 ml of 25 mM HEPES, 125 mM NaCl at pH 7.4 and left for 60 min on ice. A sample from the lysate was taken for protein content. For basal renin activity assay, 0.4 ml of nuclear lysate was added to 0.2 ml of nephrectomized rat plasma (NRP) as the source of exogenous AGT substrate. The following protease inhibitors were added to prevent the metabolism of Ang I: amastatin (AM; 2 μM), bestatin (BS; 10 μM), chymostatin (CHYM; 10 μM), benzyl succinate (BSC; 10 μM), para-chloromercuribenzoic acid (PCMB; 0.5 mM) and Ethylenediaminetetraacetic acid (EDTA) (2.5 mM) (all at final concentrations). The assay was performed at 37ºC for 90 min alone or with the renin inhibitor aliskiren (1 μM, final concentration). 24 Activation of prorenin was performed by addition of 0.1 mg trypsin (Sigma) to the nuclear sample with the peptidase inhibitors for 60 min on ice. 24 Trypsin was inactivated by incubation with an excess of soybean trypsin inhibitor (SBTI; 1 mg/ml, Sigma) for 15 min at room temperature. SBTI is a potent serine protease inhibitor that does not attenuate renin, an aspartyl protease. Following SBTI, the renin assay was performed as described above. All renin and prorenin samples were extracted on Sep-Pak C18 columns prior to the assessment of Ang I by RIA as described. 24 Blank values for the assay (aliskerin with and without lysates) were subtracted from the sample values; renin activity was expressed as nmol Ang I generated per h per mg protein.
AGT assay for renin
A rat kidney cortical homogenate as a source for renin was added to a cocktail of protease inhibitors (Sigma) (1/100) and SBTI (1 mg) in the presence or absence of aliskiren (10 μM) in a final volume of 1 ml buffer (10 mM HEPES, 125 mM NaCl, 10 μM ZnCl2, pH 7.4). NRP as the source of intact AGT was then added and incubated at 37°C for 60 min. Aliquots were removed at 5, 15, 30 and 60 min and analyzed by western blot using the AI-AGT antibody. The membrane was then stripped and probed with the second AGT antibody, Int-AGT.
Peptide metabolism
To characterize the processing of the peptides in vitro, 125I-Ang-I or Ang II (0.5 nM) were incubated with the solubilized nuclear fractions as previously described. 25 Metabolism assays were conducted at 37°C in 10 mM HEPES, 125 mM NaCl, 10 μM ZnCl2, pH 7.4, with 2-3 μg protein of the nuclear fraction in a final volume of 0.5 ml with or without the indicated inhibitors. The reaction was stopped by addition of ice-cold 1.0% phosphoric acid, centrifuged at 16,000 g, and the supernatants stored at −20°C. Samples were separated by reverse-phase high-performance liquid chromatography (HPLC) and the 125I-products were detected by a flow-through γ detector. 25 Products were identified by comparison of their retention times to 125I-angiotensin standards. Peptides were iodinated by the chloramine T method and purified by HPLC (specific activity > 2000 Ci/mmol). The aminopeptidase inhibitors amastatin AM (2 μM) and BS (10 μM) were included in the basal assay conditions. We subsequently used the following inhibitors to block thiol peptidases (PCMB; 0.5 mM); neprilysin (SCH39370 or SCH, 10 μM), prolyl oligopeptidase (Z-prolyl prolinal or ZPP, 50 μM) and thimet oligopeptidase (c-phenylpropyl-alanine-alanine-phenylalanine-p-aminobenzoate or CPP, 50 μM). Enzyme activities were expressed as fmol product per mg protein per min (fmol/mg/min).
Statistical analysis
All measurements are expressed as mean±standard error (SEM). Differences between the groups were analyzed by one-way analysis of variance (ANOVA) and Newman-Keuls multiple comparison analysis. All figures were constructed with GraphPad Prism V plotting and statistical software (GraphPad Software, San Diego, California, USA). A probability value of <0.05 was required for statistical significance.
Results
AGT antibodies
The present study utilized antibodies raised against two different epitopes of the AGT protein. The AI-AGT antibody recognized the N terminus or Ang I sequence of rat AGT while the Int-AGT antibody was directed against an internal sequence distal to Ang I. 21 We characterized both antibodies by protein immunoblots using 0.5–5.0 µl of either NRP as a source of intact AGT or commercial FBS in which the NRK-E52 cells were routinely maintained (Figure 1). As shown in Figure 1(a), the AI-AGT antibody recognized two protein bands of approximately 60 and 55 kDa from the NRP, as well as two similar sized bands in the FBS. The NRP samples were then exposed to renin from a rat cortical homogenate at 37°C or 4°C with or without the renin inhibitor aliskiren and the immunoblot probed with the AI-AGT antibody. In the absence of aliskiren, the AGT band was reduced at 15 min and essentially abolished by 30 min at 37°C (Figure 1(b)). Addition of aliskiren or incubation at 4°C prevented the disappearance of the protein up to 60 min. In contrast, the Int-AGT antibody clearly recognized AGT in the NRP samples (Figure 1(c), 0.5–5.0 µl serum), but did not detect AGT in FBS (Figure 1(c)). The Int-AGT antibody appeared to exhibit greater sensitivity to rat AGT; the antibody detected 0.5 µl of NRP while the AI-AGT antibody required a sample volume two- to five-fold higher (lane 4, Figure 1(c) versus 1(a)). Moreover, re-probing the immunoblot from Figure 1(b) with the Int-AGT antibody revealed the AGT bands in the samples incubated with renin (Figure 1(d)). These data confirm that the Int-AGT antibody recognized residues distal to the Ang I sequence of AGT in rat plasma, but not FBS, and that the AI-AGT antibody does not detect the renin-processed or des-Ang I form of rat AGT (and likely FBS). Utilizing the two AGT antibodies, we assessed expression of the protein by immunofluorescent (IMF) staining and protein immunoblots of isolated nuclear and cytosolic fractions of the NRK-52E cells (Figure 2). As shown in Figure 2(a) and 2(b), IMF staining with the AI-AGT antibody revealed primarily cytosolic staining of the protein that was apparently exclusive of the nucleus. Moreover, the immunoblot showed a 55 kDa band in the cytosolic fraction (lanes 4–6), but no immunoreactive band in the nuclear fraction of the NRK-52E cells (lanes 1–3, Figure 2(c)). In contrast, IMF staining with the Int-AGT antibody revealed the predominant localization of AGT to the area of the cell nucleus (Figure 2(d) and 2(e)). The immunoblot with the Int-AGT antibody also revealed a prominent protein band (55 kDa) in the nuclear fraction (lanes 1–3), as well as a faint band in the cytosol (lanes 4–6, Figure 2(f)).
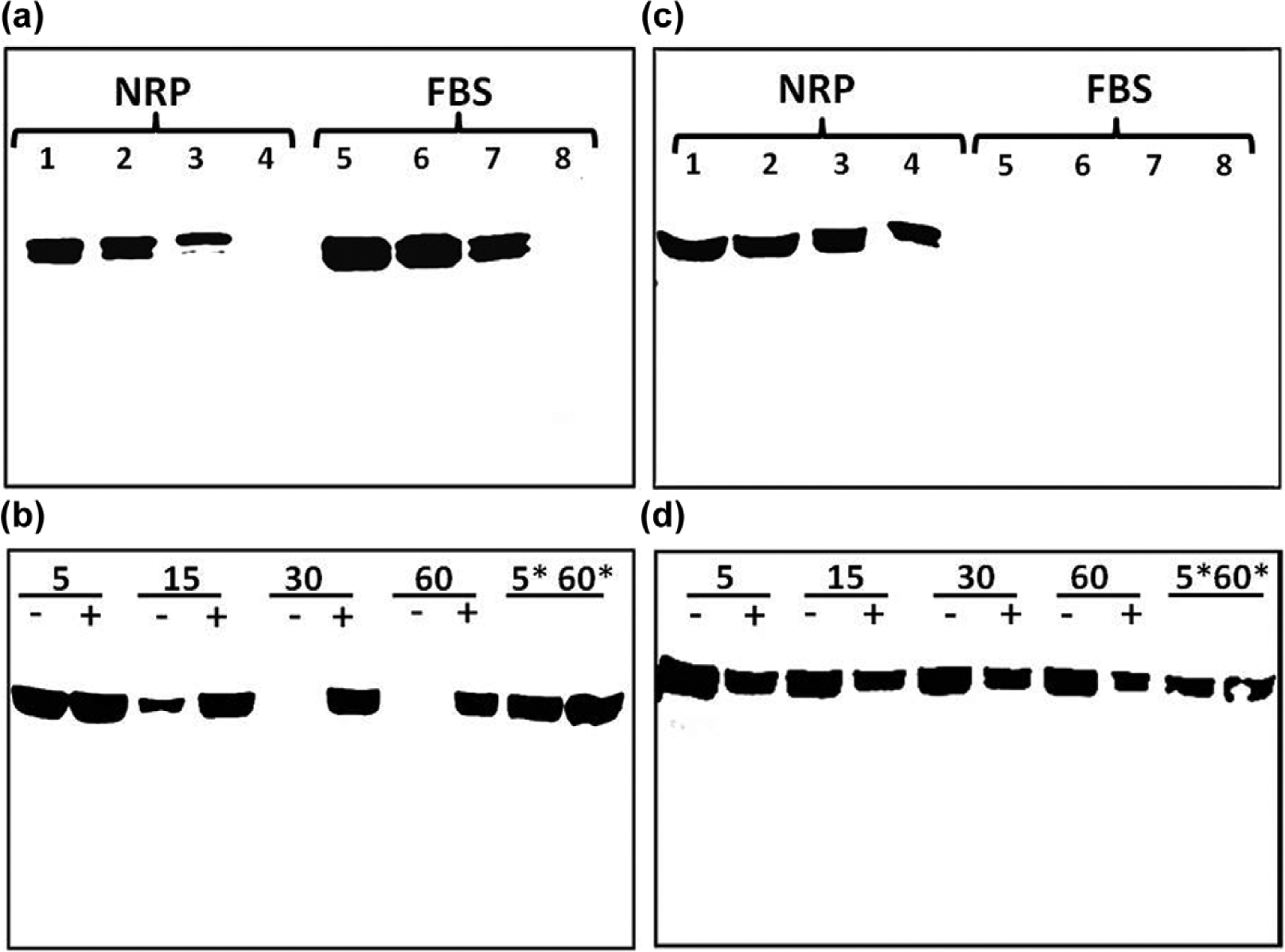
Characterization of antibodies to rat and bovine angiotensinogen (AGT): (a) immunoblot of nephrectomized rat plasma (NRP) and fetal bovine serum (FBS) probed with angiotensin I (Ang I)-AGT (AI-AGT) antibody. The AI-AGT antibody detected AGT bands at 55 and 60 kDa in both NRP and FBS samples; (b) incubation of renin with AGT in NRP for 5–60 min at 37°C or on ice (*5 and *60 min) in the absence (–) or presence (+) of the renin inhibitor aliskerin (10 μM) and probed with AI-AGT antibody; (c) immunoblot of NRP and FBS samples with the AGT antibody directed to the internal sequence of AGT (Int-AGT) distal to Ang I that recognized rat but not bovine AGT; (d) the immunoblot from (b) was stripped and re-probed with the internal sequence of AGT (Int-AGT) antibody revealing no effect of renin exposure on AGT expression.
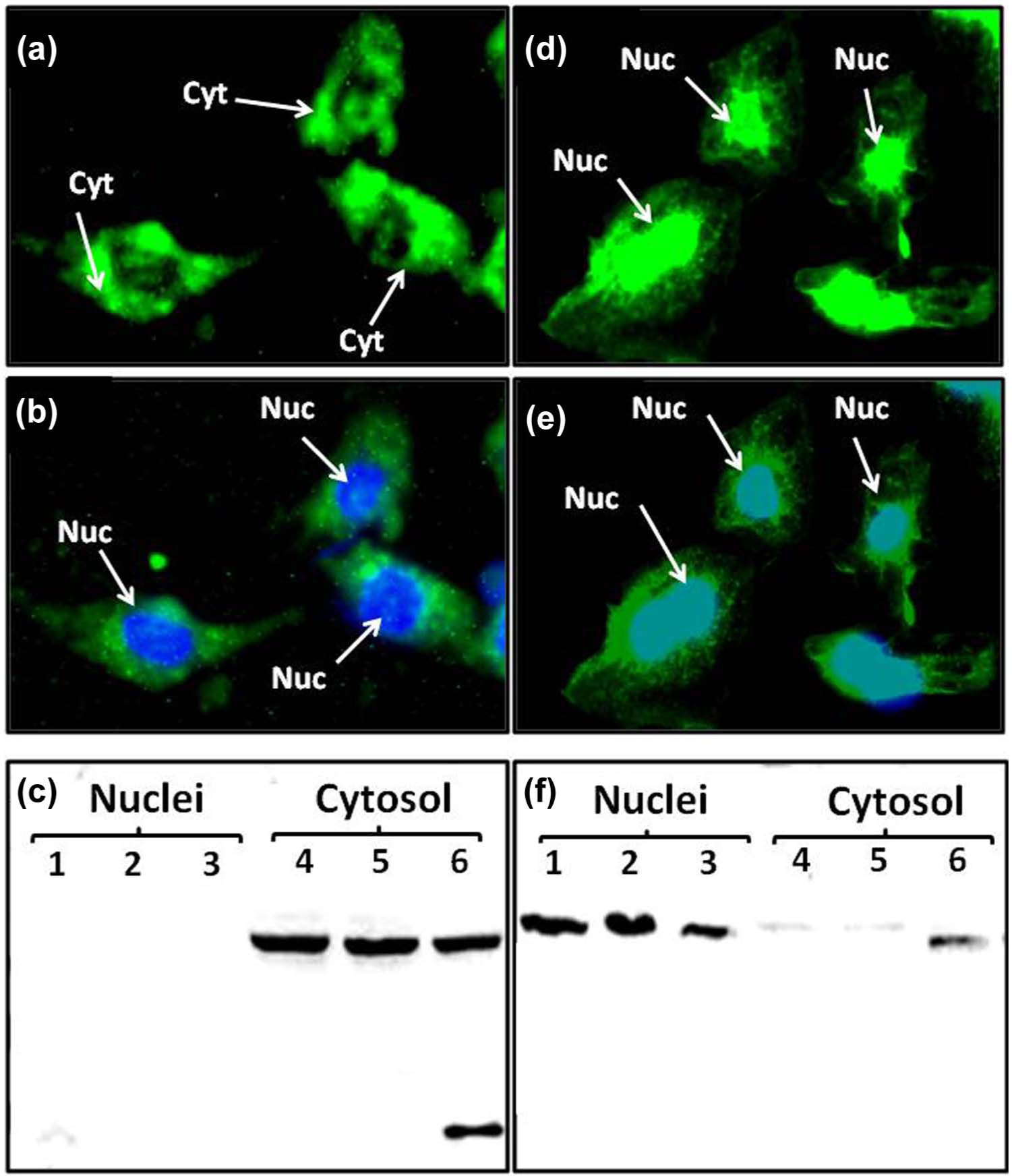
Angiotensinogen (AGT) expression in the NRK-52E cells: (a) immunofluorescent (IMF) staining of NRK-52E cells with the angiotensin I (Ang I) AGT (AI-AGT) antibody reveals predominantly cytosolic (Cyt) staining; (b) identical image of (a) with DAPI nuclear (Nuc) staining in blue; (c) immunoblot of nuclear and cytosolic fractions with AI-AGT antibody reveals a predominant 55 kDa band in cytosolic fractions; (d) IMF staining with internal sequence of AGT (Int-AGT) antibody reveals nuclear associated staining; (e) identical image of (d) with DAPI nuclear (Nuc) staining in blue; (f) immunoblot of nuclear and cytosolic fractions reveals predominant 55 kDa band in the nuclear fraction. IMF staining was representative of three different cell passages. Immunoblot fractions were from three different cell passages.
Renin and PRR
The apparent absence of intact AGT in the nucleus suggested that renin or a renin-like enzyme cleaved AGT either within the nucleus or outside the organelle with subsequent nuclear transport of the precursor. As shown in Figure 3(a), IMF staining for renin revealed the predominant nuclear localization of the enzyme in the cells. Protein immunoblots of the nuclear and cytosolic fractions also revealed a predominant immunoreactive band at ~ 55 kDa in the nuclear fraction and a faint band in the cytosol (Figure 3(b)). We next assessed renin activity in isolated nuclei of the NRK-52E cells by addition of exogenous NRP AGT in the absence or presence of aliskiren. Renin activity averaged 0.77±0.05 nmol Ang I/mg/h and aliskiren reduced activity by 83% (Figure 3(c)). To determine the prorenin levels, nuclear extracts were pretreated with trypsin to convert prorenin to renin and the generation of Ang I from NRP AGT was determined in the presence of the trypsin inhibitor SBTI. Trypsin activation of nuclear prorenin significantly increased enzyme activity approximately three-fold to 1.93±0.23 nmol Ang I/mg protein/h and aliskiren inhibited 93% of the Ang I-forming activity (Figure 3(c)). These findings suggest the presence of both renin and prorenin in the NRK-52E cell nuclei. Since prorenin may preferentially bind to the prorenin receptor to increase catalytic activity and/or induce cell signaling, we stained the cells with an antibody against the prorenin receptor. As shown in Figure 3(d), IMF staining revealed primarily perinuclear staining of the PRR in cells counter-stained with the nuclear marker DAPI shown in blue. In comparison, cells were also stained with an antibody to calnexin, a protein intrinsic to the perinuclear/ endoplasmic reticulum (ER) membrane and the DAPI marker (Figure 3(e)).

Renin and prorenin receptor (PRR) expression in NRK-52E cells: (a) immunofluorescent (IMF) staining of NRK-52E cells with the renin antibody reveals nuclear-associated staining; (b) immunoblot of nuclear and cytosolic fractions with renin antibody reveals a 55 kDa (arrow) band in the nuclear fraction from cells of three different passages. Higher molecular weight bands were evident in the cytosolic fractions. (c) Renin activity in the nuclear fraction of control (CON) or trypsin (TRP) -treated nuclear lysates treated with or without the renin inhibitor aliskerin (ALK). Values are means±standard error of the mean (SEM), *p<0.01 vs CON, #p<0.01 vs TRP; n=3 different passages. (d) IMF staining of PRR reveals perinuclear staining and DAPI nuclear staining (Nuc) in blue; (e) IMF staining of the endoplasmic reticulum (ER) marker calnexin and DAPI nuclear staining (Nuc) in blue. Cyt: cytosolic staining.
Ang peptides
The expression of both AGT and renin within the NRK-52E cell nuclei may suggest a pathway for the intracellular synthesis of Ang peptides. Therefore, we assessed the cell staining and content of Ang II and Ang-(1-7) by two distinct antibodies specific to the C-terminus of each peptide. As shown in Figures 4(a) and 4(b), the affinity-purified antibodies to Ang II and Ang-(1-7) revealed predominantly nuclear staining. Consistent with this pattern of staining, nuclear extracts from four different cell passages contained 59±2 fmol/mg protein of Ang II and 57±22 fmol/mg protein of Ang-(1-7) (Figure 4(c)). We then examined the potential processing enzymes that may contribute to Ang II or Ang-(1-7) within the nucleus. Solubilized nuclear fractions were incubated with 125I-Ang I or 125I-Ang II for 15–120 min at 37○C and the products were analyzed by a HPLC-coupled γ-detector. 25 As shown in the chromatographic panels of Figure 5, Ang-(1-7) was the major peak of Ang I metabolism at 15 and 60 min (Figure 5(a) and 5(b), respectively). Ang I exhibited a half-life of 73 min in the nuclear cell extracts and Ang-(1-7) was the primary product over the 120 min time course (Figure 6). In contrast, there was minimal metabolism of the Ang II in the nuclear extracts with >70% of the peptide intact at 120 min (data not shown).

Expression of angiotensin (Ang) II and Ang-(1-7) in NRK-52E cells. Immunofluorescent staining using affinity-purified antibodies for (a) Ang II and (b) Ang-(1-7) reveal nuclear-associated staining (Nuc). (c) Immunoreactive concentrations of Ang II and Ang-(1-7) in the nuclear fractions of NRK-52E cells. Values are means±standard error of the mean (SEM); n=4.

Conversion of angiotensin (Ang) I to Ang-(1-7) in isolated nuclei from NRK-52E cells. High-performance liquid chromatography (HPLC) analysis of 125I-Ang I metabolism at 37°C revealed primarily Ang-(1-7) at (a) 15 min and (b) 60 min. The neprilysin inhibitor SCH39370 (SCH; 10 μM) or prolyl oligopeptidase inhibitor z-prolyl-prolinal (ZPP; 50 μM) did not inhibit Ang-(1-7) generation (shown in (c) and (d), respectively). The thimet oligopeptidase inhibitor (CPP; 50 μM) and the thiol inhibitor (PCMB; 500 μM) essentially abolished Ang-(1-7) production (shown in (e) and (f), respectively). A minor peak of Ang-(1-4) was detected in panels (b)–(f).
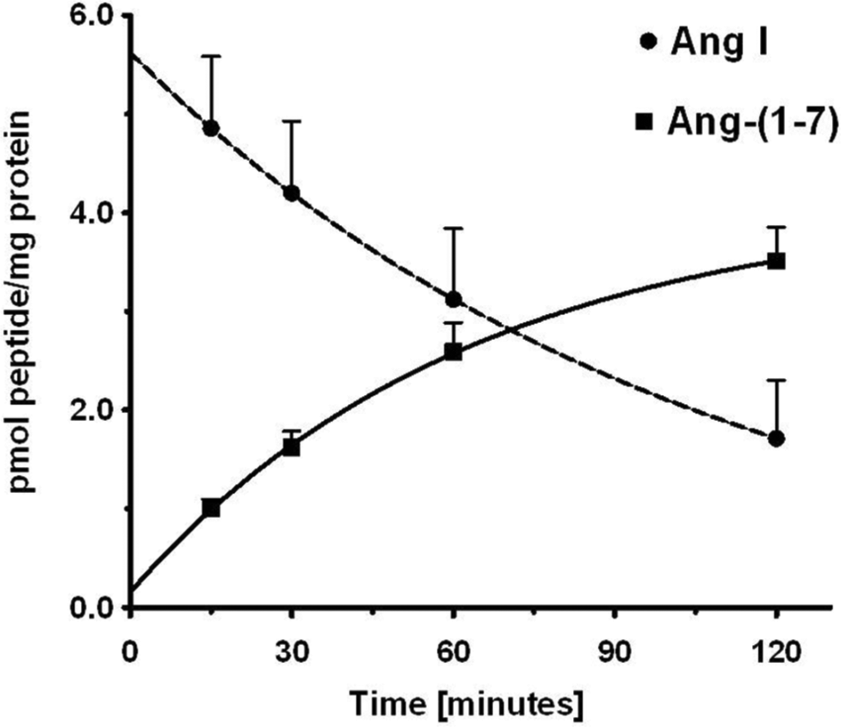
Time course for the disappearance of angiotensin (Ang) I and formation of Ang-(1-7) in isolated nuclei from NRK-52E cells. The conversion of 125I-Ang I to 125I-Ang-(1-7) in nuclei at 37°C was quantitated by high-performance liquid chromatography (HPLC) separation coupled to a γ-counter. Values are means±standard error of the mean (SEM), n=4 of different cell passages. Non-linear decay curve for Ang I and exponential one-phase association of Ang-(1-7) were constructed in GraphPad Prism V.
There are several endopeptidases including neprilysin, prolyl oligopeptidase and thimet oligopeptidase that directly process Ang I to Ang-(1-7). 26 The neprilysin agent SCH39370 and the prolyl oligopeptidase inhibitor ZPP did not block the generation of Ang-(1-7) in the nuclear extract (Figure 5(c) and 5(d), respectively). Both the thimet oligopeptidase inhibitor CPP and the thiol protease inhibitor PCMB markedly reduced the peak of Ang-(1-7) and preserved the peak of Ang I (Figure 5(e) and 5(f), respectively). Evaluation of these inhibitors at the 60 min incubation period revealed that CPP and PCMB inhibited Ang-(1-7) formation by over 80% and suggests that thimet oligopeptidase converts Ang I to Ang-(1-7) in isolated nuclei (Figure 7). We next assessed the Ang-(1-7) and Ang-(1-4)-forming activities at 120 min in the presence of CPP and SCH39370. Again, thimet oligopeptidase accounted for the majority of Ang-(1-7) at this time (Figure 8); however, there remained small peaks of Ang-(1-7) and Ang-(1-4) with CPP (Figure 8(b)). Addition of SCH39370 to CPP appeared to further reduce both Ang-(1-7) and Ang-(1-4) (Figure 8(c)), but the response did not reach statistical significance (Figure 8(d)).

Influence of peptidase inhibitors on the generation of angiotensin (Ang)-(1-7) from Ang I in the isolated nuclear fraction of NRK-52E cells. Peptidase inhibitors were neprilysin inhibitor (SCH), prolyl oligopeptidase inhibitor (ZPP), thimet oligopeptidase inhibitor (CPP) and thiol protease inhibitor (PCMB). All values represent mean±standard error of the mean (SEM), n=4 from different cell passages; *p<0.01 vs control.

Influence of combined thimet oligopeptidase and neprilysin inhibition on angiotensin (Ang)-(1-7) and Ang-(1-4) formation in isolated nuclei NRK-52E cells: (a) metabolism of 125I-Ang I to Ang-(1-7) and Ang-(1-4) following 120 min incubation with nuclei at 37°C; (b) addition of thimet oligopeptidase inhibitor (CPP; 50 µM) blocked the majority of Ang-(1-7) formation; (c) addition of CPP and neprilysin inhibitor SCH39370 (SCH; 10 µM); (d) influence of CPP or CPP/SCH on the formation of Ang-(1-7) and Ang-(1-4) in nuclei at 120 min. All values represent mean±standard error of the mean (SEM), n=3 from different cell passages; *p<0.05 vs control.
Discussion
In the present study we characterized the RAS in the NRK-52E epithelial cell line to facilitate our understanding of this intracellular system. The NRK-52E cells are widely utilized to examine the functional aspects of the Ang II-AT1 receptor pathway, as well as the counter-regulatory actions of the Ang-(1-7)-AT7/Mas axis.27–32 The current findings revealed that NRK-52E cells express multiple components of an intracellular RAS and parallel our previous findings in the isolated tubules of the sheep kidney. 9 In this case, both immunofluorescent staining and western blots demonstrated that NRK-52E cells express AGT, prorenin, PRR and the peptides Ang II and Ang-(1-7) in the nuclear or perinuclear compartments. Activity studies revealed renin-dependent conversion to Ang I; however, prorenin was the more abundant form of the enzyme in a 3:1 ratio in the nuclear fraction. Finally, we demonstrate endopeptidase activity in the nuclear fraction that converts Ang I directly to Ang-(1-7) without the prerequisite formation of Ang II. The identity of the nuclear endopeptidase is likely thimet oligopeptidase based on the sensitivity to both the selective inhibitor CPP and the thiol agent PCMB.
Utilizing two different antibodies to rat AGT, we primarily detected the processed form of AGT (des-Ang I Aogen) in the nucleus. Sherrod et al. detected AGT primarily in the nucleus of human astrocytes and identified a nuclear localization signal in the C-terminal portion of the protein; however, these studies did not determine whether the precursor was processed or intact. 7 In the current study, the presence of the cleaved AGT in the nucleus suggests proteolytic processing potentially by renin which may contribute to local levels of Ang II and Ang-(1-7) detected in the nuclear fraction. Indeed, this may constitute a pathway for the generation and delivery of peptide ligands to nuclear Ang receptors evident in the tubule epithelium, as well as in other tissues. Alternatively, AGT may be processed outside of the nucleus and the des-Ang I form subsequently transported to the nucleus. Apart from its role as the precursor to Ang II or Ang-(1-7), nuclear AGT may potentially contribute to cell signaling and function. Moreover, two studies reveal that both intact and the Ang I-cleaved forms of AGT attenuated angiogenesis.33,34 In this regard, Pan et al. reported high affinity binding sites for AGT that internalized following binding in primary cultures of human proximal tubules. 35 In contrast to the nuclear localization of des-Ang I AGT, the Ang I-directed antibody revealed the protein in both the cytosolic compartment of the NRK-52E cells, as well as in FBS. The circulating RAS is upregulated during pregnancy and the presence of AGT in FBS is not surprising. Since the Int-AGT antibody failed to recognize FBS AGT or detect AGT to the same extent as the AI-AGT antibody in the cell cytosol, it is possible that cytosolic AGT may primarily originate from uptake of AGT in the cell media. Recent studies have questioned the prevailing tenet that tubular AGT solely reflects local synthesis of the protein.36,37 Indeed, the protein transporter megalin may mediate the cellular uptake and stable sequestration of AGT within a subset of renal proximal tubules. 37 However, it is not clear as to the cellular fate of AGT taken up from the tubular fluid and whether this pathway contributes to the generation of Ang peptides. Based on the present findings, the intact form of AGT in the NRK-52E cytosol may localize to a subcellular compartment separate from rat renin. Alternatively, provided the cytosolic form of AGT arises primarily from FBS, the presence of intact AGT may reflect inefficient processing by rat renin due to species differences in the recognition of AGT by renin. Clearly, additional studies are required to elucidate whether the cytosolic from of AGT in the NRK-52E cells reflects the uptake of FBS AGT and the extent that megalin contributes to this process.
The present characterization of the RAS in the NRK-52E cells arose from the demonstration of nuclear renin and AGT in isolated sheep proximal tubules, as well as the expression of Ang receptors on isolated nuclei. 9 The localization of renin to the non-juxtaglomerular (JG) compartments of the kidney is somewhat controversial. Rohrwasser and colleagues originally demonstrated renin in the principal cells of the collecting duct in both the mouse and rat kidney.38,39 Subsequent studies suggest that renin regulation in the collecting duct of the rat kidney differs from that of JG-derived renin.40,41 Immunoreactive renin and mRNA expression were also reported in the proximal tubules; however, the intracellular localization of the protease was not determined.42–44 In this regard, an intracellular or truncated form of renin was identified in the mitochondria which may contribute to local Ang II generation.45–47 The isolated nuclei of the NRK-52E cells contained both active renin and prorenin that converted exogenous AGT to Ang I; this activity was essentially abolished by the renin inhibitor aliskerin. IMF staining also revealed that the prorenin receptor localized to the perinuclear compartment of the NRK-52E cells which is consistent with the findings of Schefe and colleagues. 15 Apart from contributing to activation of the enzyme, the receptor may bind prorenin to stimulate cellular pathways including mitogen-activated protein kinase (MAP) kinase and transforming growth factor beta TGF-β release that are not dependent on the Ang II-AT1 receptor axis.13,14,48,49 The extent that renin or activated prorenin contributes to the intracellular formation of Ang II or Ang-(1-7) in the NRK-52E cells awaits further study. Furthermore, studies are necessary to elucidate whether the PRR contributes to functional signaling pathways in these cells through an interaction with intracellular prorenin.
In addition to AGT and renin, both immunoreactive Ang II and Ang-(1-7) were identified in the isolated nuclear fractions of the NRK-52E cells. Camargo de Andrade et al initially reported IMF staining for both Ang II and Ang-(1-7) in the nucleus of isolated mesangial cells. 10 Consistent with presence of Ang-(1-7) in the nuclei, we find an endopeptidase-like activity that processed Ang I predominantly to Ang-(1-7). The inhibitor profile suggested that the nuclear peptidase is likely thimet oligopeptidase (EC 3.4.24.15). Inhibitors selective for other endopeptidases that form Ang-(1-7) including neprilysin (EC 3.4.24.11) and prolyl-oligopeptidase (EC 3.4.21.26) did not significantly reduce the conversion of Ang I to Ang-(1-7). Thimet oligopeptidase is a zinc-dependent member of the metallopeptidase M3 family, but is sensitive to thiol or cysteine inhibitors such as PCMB and the selective inhibitor CPP.50–52 Interestingly, thimet oligopeptidase was localized to the nucleus of neurons in the rat brain by immunocytochemical staining.53,54 In this regard, Thompson et al. identified the nuclear localization signal sequence for human thimet oligopeptidase. 55 Moreover, the overexpression of intracellular thimet oligopeptidase in human embryonic cells (HEK) cells attenuated Ang II-AT1 receptor signaling as well as the actions of other G protein-coupled receptors.56,57 Blockade of Ang-(1-7) formation from Ang I did not result in the generation of Ang II in the nuclear fraction nor did we find extensive metabolism of Ang II to Ang-(1-7) or any other products. Although these findings suggest that Ang II is stable in the nuclear fraction, they do not identify the pathway for the nuclear formation or sequestration of Ang II and we can only suggest several possibilities for the nuclear expression of Ang II at this time. The peptide may arise from the direct processing of AGT through a non-renin mechanism or an intermediate peptide substrate such as Ang-(1-12).2,26 Alternatively, nuclear Ang II may reflect the trafficking of the Ang II-AT1 receptor complex from the plasma membrane to the nucleus.58–62 Finally, Gonzalez-Villalobos et al. report that uptake of both Ang II and Ang-(1-7) was facilitated by the protein transporter megalin in the opossum kidney (OK) epithelial cell line, an accepted cell model for megalin-dependent uptake of albumin; however, the intracellular fate of the internalized peptides was not evaluated in these studies.63,64
In conclusion, the present study characterized the NRK-52E epithelial cell line as a tubule model for the expression of the intracellular RAS components within the kidney. The kidney is one of the primary targets to specifically block the ACE-Ang II-AT1 receptor axis by various therapeutic approaches including ACE inhibition, AT1 receptor antagonism, and more recently, renin blockade to attenuate blood pressure and renal injury. However, these regimens may not completely prevent the pathological consequences of an activated RAS. This may result from either insufficient drug doses to effectively block the intracellular RAS, low lipophilicity of certain agents that leads to inadequate intracellular concentrations, or an inability to inhibit downstream signaling pathways independent of Ang II generation. 65 Indeed, increasing the dose of candesartan to a level higher than the maximal recommended dose for heart failure and hypertension significantly reduced persistent proteinuria without a further decline in blood pressure. 66 Renin inhibitors such as aliskiren do not block the binding of prorenin to the receptor. 67 Therefore, the proinflammatory and profibrotic responses of an activated RAS may not be completely attenuated by renin inhibitors that would effectively reduce blood pressure. In this regard, introducing membrane permeable nonpeptide agonists for the AT7 and AT2 receptors along with renin inhibition and ARBs may overcome the loss of either AT7 or AT2 receptor activation by renin inhibitors, as well as achieve maximal blockade of the deleterious effects of the AngII-AT1 receptor axis.
Footnotes
Acknowledgements
This present study represents partial fulfillment of the requirements for the degree of Doctorate of Philosophy in the Department of Physiology and Pharmacology, Wake Forest University School of Medicine for Ebaa Alzayadneh. The authors acknowledge Nancy Pirro and Brian Westwood for their technical contribution to these studies.
Conflicts of interest
The authors declare that there are no conflicts of interest.
Funding
This study was supported by the National Institutes of Health (HL-51952, HL-56973) and the Wake Forest Venture Fund.