
Abstract
Antenatal corticosteroid exposure reduces renal function and alters the intrarenal renin-angiotensin system to favor angiotensin activation of angiotensin type 1 receptor (AT1R) mediated responses in ovine offspring. This study aimed to assess whether antenatal steroid exposure would affect renal responses to the direct intrarenal infusion of angiotensin-(1–7) in rams and the angiotensin receptors involved in mediating responses to the peptide.
Adult, uninephrectomized rams exposed to either betamethasone or vehicle before birth received intrarenal angiotensin-(1–7) infusions (1 ng/kg/min) alone or in combination with antagonists to angiotensin receptors for 3 h. Basal sodium excretion (UNa) was significantly lower and mean arterial pressure was significantly higher in betamethasone- compared to the vehicle-treated sheep. Angiotensin-(1–7) decreased UNa more in betamethasone- than in vehicle-treated sheep. Candesartan reversed the response to angiotensin-(1–7) but D-Ala7-angiotensin-(1–7) did not. Angiotensin-(1–7) infusion decreased effective renal plasma flow in both groups to a similar extent and the response was reversed by candesartan, but was not blocked by D-Ala7-angiotensin-(1–7). Glomerular filtration rate increased significantly in both groups after 3 h infusion of angiotensin-(1–7) plus candesartan. These results suggest that antenatal exposure to a clinically relevant dose of betamethasone impairs renal function in rams. Moreover, angiotensin-(1–7) appears capable of activating the AT1R in uninephrectomized rams.
Keywords
Introduction
Glucocorticoid administration for the prevention of complications related to prematurity has been standard care for women with preterm labor before 35 weeks gestation since 1994. 1 A single course of antenatal betamethasone accelerates fetal lung maturation and enhances the survival of preterm infants. However, this regimen has been associated with higher blood pressure in adolescence and decreased insulin sensitivity and renal function in adulthood.2,3
Studies using sheep experimental models demonstrate that prenatal glucocorticoid exposure results in elevated blood pressure in adulthood.4,5 The reasons for this increase are not fully known: however, the mechanisms are likely to involve a reduction in nephron number6,7 and alterations in the intrarenal renin-angiotensin system (RAS) and renal function.5,8,9
The RAS plays an essential role in controlling sodium and water excretion. While angiotensin II (ANG II) is considered the predominant RAS mediator,10–12 angiotensin -(1–7) (ANG- (1–7)) can produce a number of physiological effects, including natriuresis, diuresis, vasodilation, and the release of vasopressin and prostaglandins11,13,14 Both ANG II and ANG-(1–7) exert their effects by binding to specific receptors: angiotensin type 1 receptor (AT1R) and angiotensin type 2 receptor (AT2R) for ANG II and the Mas receptor for ANG-(1–7).15–18 We have speculated, based upon the results of several studies, that one of the global effects of antenatal betamethasone exposure is a shift in the balance of ANG II to ANG-(1–7) favoring responses mediated by activation of the AT1R pathway by several means including alterations in receptor expression.19–21
At this point, however, there is limited information on the effects of antenatal betamethasone exposure on renal responses to ANG-(1–7). Based upon results of studies with systemic infusions of ANG-(1–7) in our model,19,22 we hypothesized that responses to direct infusions of ANG-(1–7) into the kidney would be attenuated in adult sheep that had been exposed to betamethasone prenatally. To test this hypothesis we infused a low dose of ANG-(1–7) directly into the renal artery in a group of 12 adult male uninephrectomized sheep of which half were exposed to betamethasone and the remainder to vehicle at 0.6 gestation and measured aspects of renal function.
Materials and methods
Animal preparation
All procedures were approved by the Institutional Animal Care and Use Committee of Wake Forest University School of Medicine. Time-dated pregnant ewes were randomly assigned to receive either two 0.17 mg×kg-1 intramuscular injections of a 1:1 mixture of betamethasone acetate and betamethasone phosphate (Celestone Soluspan, Schering, Kenilworth, New Jersey, USA) or vehicle alone which contained 3.4 mg of monobasic sodium phosphate, 7.1 mg of dibasic sodium phosphate, 0.1 mg of sodium ethylenediaminetetra-acetic acid, and 0.2 mg of benzalkonium chlorine per ml given 24 h apart at days 80 and 81 of gestation. The ewes were allowed to deliver naturally at term (term is approximately 145 days in our flock).
Male offspring (five or six in each group) were transferred to the laboratory at 12–18 months of age. The research team was blinded to the sheep allocation. The temperature in the laboratory facilities and in the sheep pens was kept stable at 20±2°C, and the humidity at approximately 35–40%. Sheep were fed a standard commercial diet (PMI Rumilab® Brentwood, Missouri, USA) containing 0.75% NaCl and had ad libitum access to clean tap water. Twelve hour day/night light cycles were maintained throughout the experiment.
Surgical procedure
A midline suprapubic laparotomy was made in the lower abdomen to expose the bladder. A 14 French silicone Foley catheter (Bard Medical, Covington, Georgia, USA) was placed in the bladder and exteriorized through the left flank. Two polyvinyl catheters were placed in the left carotid artery and two catheters were inserted into the left external jugular vein. These were tunneled subcutaneously and emerged through a left anterior neck incision. All vascular catheters were fastened to their corresponding vessels and the distal vessels were all ligated.
A paravertebral left or right flank incision was made to expose the corresponding renal artery. A modified, Teflon coated, 22 gauge endovascular catheter 25 mm long and with a 0.9 mm external diameter was placed into the left or right renal artery in a non occlusive fashion and attached to a Tygon catheter. The Tygon catheter was tunneled a short distance to emerge adjacent to the spine. We then removed the contralateral kidney.
All the animals received daily intravenous doses of ampicillin 1000 mg, gentamicin 80 mg and ketorolac 100 mg on the day of surgery and two days thereafter. All vascular catheters were flushed with a heparin solution on a daily basis to confirm patency. During the post-operative period and throughout the experimental period, each pair of animals was housed together in individual carts with free access to food and water.
Protocol of experiments
After allowing five days for recovery, sheep underwent a series of experiments to evaluate the response to a 3 h infusion of ANG-(1–7) alone or in combination with specific antagonists. The sequence of infused substances was: (a) saline (1 ng/kg/min); (b) ANG-(1–7) (1 ng/kg/min, Bachem BioScience Inc., Torrance, California, USA); (c) ANG-(1–7) and D-Ala7-angiotensin-(1–7) (D-Ala-ANG-(1–7)) (10 ng/kg/min, Bachem BioScience Inc., Torrance, California, USA); (d) D-Ala-ANG-(1–7) (10 ng/kg/min, Bachem BioScience Inc., Torrance, California, USA); (e) ANG-(1–7) and candesartan (AT1R antagonist, 0.3 mg/kg by bolus injection via renal artery, Astra Zeneca, Möindal, Sweden) and (f) candesartan alone (AT1R antagonist, 0.3 mg/kg by bolus injection via renal artery, Astra Zeneca, Möindal, Sweden). All drugs were infused directly into the renal artery with syringes mounted on a multichannel pump. The dose for ANG-(1–7) and candesartan were obtained from our previous work documenting ANG II effects during intrarenal infusions in sheep. 23 The dose of ANG-(1–7) was equivalent to ANG II because part of our long-term goal is to determine if antenatal betamethasone alters the relative efficacy of these two peptides in the kidney. The dose of D-Ala-ANG-(1–7) was obtained from a previous study in which D-Ala-ANG-(1–7) at a 10-fold higher dose than ANG-(1–7) was able to abolish the effect of ANG-(1–7). 24 We didn’t evaluate higher doses of D-Ala-ANG-(1–7) because they can potentially elicit nonspecific effects. 13 Forty-eight to 72 h were allowed between experiments for adequate clearance of previously infused drugs. Not all of the animals received all of the treatments due to occasional catheter failure.
Blood and urine samples were serially obtained prior to initiating all infusions, subsequent to initiating infusions and over the course of the following 3 h.
To guarantee adequate hydration status we administered 2000 ml of normal saline intravenously the afternoon prior to beginning each experiment. After all experiments had concluded, the animals were euthanized with 10 ml Beuthanasia D® (390 mg pentobarbital sodium and 50 mg phenytoin sodium per ml) intravenously, after which organ harvesting was performed.
Blood and urine samples
Timed samples of blood and urine were collected 1 h prior to beginning any infusion and at the end of the infusions. Urine samples were collected in plain tubes. All blood and urine samples were centrifuged at 3000 rpm, 4ºC, for 10 min immediately after collection, aliquoted and stored at −20°C for subsequent analysis. Urine volumes were recorded.
Blood pressure measurement
Mean arterial blood pressure was monitored continuously throughout each experiment with a Cobe transducer connected to a DigiMed analyzer, which digitized the arterial blood pressure signal that was then recorded on a computer. Pressure was sampled at 100 Hz and averaged over 1 min intervals. The reported blood pressure values are the average blood pressure over 1 h prior to the time point reported.
Urinary concentration of electrolytes
Urinary concentrations of electrolytes were determined using a Medical Easy-Lyte instrument (Bedford, Massachusetts, USA). Prior to measuring the samples, the machine underwent the manufacturer recommended procedures which included a cleaning cycle for the probe, analyzer calibration and use of standardized control samples. The instrument’s margin of error for urinary electrolytes is 5%. All results are reported as mmol/l.
Changes of fractional excretion of sodium (FENa)
Changes of FENa were determined by subtracting the value of FENa at 3 h from that at baseline. The following formula was used for FENa: FENa=sodium excretion/(plasma sodium concentration×glomerular filtration rate (GFR)).
GFR measurement
GFR was measured using an anthrone method and a colorimetric assay for determining the concentration of plasma inulin as previously described.25–27 The following formula was used for GFR (ml/min)=rate of inulin infusion (mg/min)/plasma inulin (mg/ml).
Effective renal plasma flow (ERPF)
Plasma para-aminohippurate (PAH) was measured using a previously described colorimetric assay. 28 Renal PAH excretion was used to derive effective plasma flow rate (ERPF) according to the following formula: ERPF= rate of PAH infusion (mg/min)/plasma PAH (mg/ml).
Plasma ANG-(1–7)
Blood for measurement of ANG-(1–7) was collected into chilled tubes containing a mixture of 25 mmol/l ethylene-diamine-tetraacetic acid (Sigma Chemical Co., St Louis, Missouri, USA), 0.44 mmol/l 1,20-orthophenanthrolene monohydrate, 1 mmol/l Na+ para-chloromercuribenzoate, and 3 μmol/l of WFML (rat renin inhibitor: acetyl-His-Pro-Phe-Val-Statine-Leu-Phe). The concentration of ANG-(1–7) was measured by radioimmunoassay described previously.29,30
Statistical analysis
The study data were tabulated and graphed using Prism software (Graphpad Software, La Jolla, California, USA). Statistical analysis was accomplished with the use of t-test and two-way analysis of variance for repeated measurements. Means between the betamethasone- and vehicle-exposed sheep were compared. A p-value of <0.05 was considered statistically significant. The data are reported as means±standard error of the mean (SEM). Except for blood pressure all other measurements were normalized by body weight.
Results
Body weights and remaining kidney weights
The mean body weights at the time of experiments were no different between vehicle- and betamethasone-treated sheep. At the time of necropsy the mean weight of the remaining kidney in betamethasone-treated sheep was similar to that in vehicle-treated animals (Table 1).
Body weight, kidney weight and ratio of kidney weight to body weight.

Basal sodium excretion
Basal sodium excretion was lower in betamethasone compared to vehicle-treated sheep (0.33±0.02 mmol/h/kg vs 0.53±0.06 mmol/h/kg, p<0.05).
Plasma ANG-(1–7) concentration
We measured plasma ANG-(1–7) in a subset of both groups of sheep following intrarenal infusion of the peptide. Plasma levels of ANG-(1–7) did not increase significantly as compared to baseline in either the vehicle- or betamethasone-treated groups (7±1 pg/ml to 9±5 pg/ml in vehicle sheep, n=3; 7±2 pg/ml to 8±3 pg/ml in betamethasone sheep, n=4).
Sodium excretion
Sodium excretion was unchanged in betamethasone- and vehicle-treated sheep during 3 h intrarenal infusion of either saline, D-Ala-ANG-(1–7), or candesartan (data not shown). Infusion of ANG-(1–7) alone decreased sodium excretion in both groups (F=8.8, p=0.02) and there was a significant effect of betamethasone (F=5.6, p=0.04) (Figure 1(A)). This antinatriuretic response was not blocked by the ANG-(1–7) receptor antagonist D-Ala-ANG-(1–7) (Figure 1(B)): however, a significant increase in sodium excretion in both groups of sheep was apparent during infusion of ANG-(1–7) with the AT1R antagonist candesartan. There was a significant effect of ANG-(1–7)/candesartan infusion (F=18.1, p=0.003) and betamethasone (F=8.8, p=0.02) (Figure 1(C)).

Sodium excretion before and after 3 h of intrarenal infusion of (A) angiotensin-(1–7) (ANG-(1–7)) alone, (B) ANG-(1–7) with D-Ala7-ANG-(1–7) (D-Ala), or (C) ANG-(1–7) with candesartan in uninephrectomized vehicle- and betamethasone-treated sheep. A statistically significant difference within groups after infusion (*p<0.05 or φp<0.01). #indicates a statistically significant difference between groups after infusion (p<0.05).
Changes of FENa
Changes in the FENa in both groups are shown in (Figure 2). There was an overall effect of the treatment on FENa (F=7.4, p<0.01). There was a significant effect in both vehicle- and betamethasone-treated sheep (p<0.05, Bonferroni post-tests) between infusion of ANG-(1–7) with D-Ala-ANG-(1–7) and infusion of ANG-(1–7) with candesartan.

Changes in the fractional excretion of sodium (FENa) after 3 h of intrarenal infusion of angiotensin-(1–7) (ANG-(1–7)) alone, ANG-(1–7) with D-Ala7-ANG-(1–7) (D-Ala), or ANG-(1–7) with candesartan in uninephrectomized vehicle- and betamethasone-treated sheep. (F=7.4, p<0.01 with treatment, Bonferroni post-test, *p<0.05).
Mean arterial pressure (MAP)
The betamethasone exposed animals had higher resting MAPs than did the vehicle-treated sheep (83.5±1.9 mm Hg vs 76.9±1.4 mm Hg, p<0.05). MAP did not change significantly during any of the intrarenal infusions (data not shown).
ERPF
Saline, D-Ala-ANG-(1–7), and candesartan did not alter ERPF significantly in either group of sheep (data not shown). However, infusion of ANG-(1–7) decreased ERPF in both groups (F=12.8, p=0.004) to a similar extent (Figure 3(A)). Infusion of ANG-(1–7) and D-Ala-ANG-(1–7) also decreased ERPF significantly in both groups (F=7.0, p=0.02) with no significant difference in the response between vehicle- and betamethasone-treated sheep (Figure 3(B)). In contrast, ERPF increased similarly in both groups of sheep after 3 h infusion of ANG-(1–7) and candesartan (F=44.4, p=0.0002) (Figure 3(C)).

Effective renal plasma flow (ERPF) before and after 3 h of intrarenal infusion of (A) angiotensin-(1–7) (ANG-(1–7)) alone, (B) ANG-(1–7) with D-Ala7-ANG-(1–7) (D-Ala), or (C) ANG-(1–7) with candesartan in uninephrectomized vehicle- and betamethasone- treated sheep. A statistically significant difference within groups after infusion (*p<0.05 or φp<0.01).
Glomerular filtration rate (GFR)
After 3 h infusion of saline, D-Ala-ANG-(1–7), or candesartan, GFR was unchanged (data not shown). GFR tended to decrease after ANG-(1–7) infusion with or without D-Ala-ANG-(1–7), but the difference did not reach statistical significance (Figure 4(A) and (B)). However, GFR increased to about the same level in both groups of sheep following the 3 h infusion of ANG-(1–7) and candesartan (F=7.4, p=0.03) (Figure 4(C)).
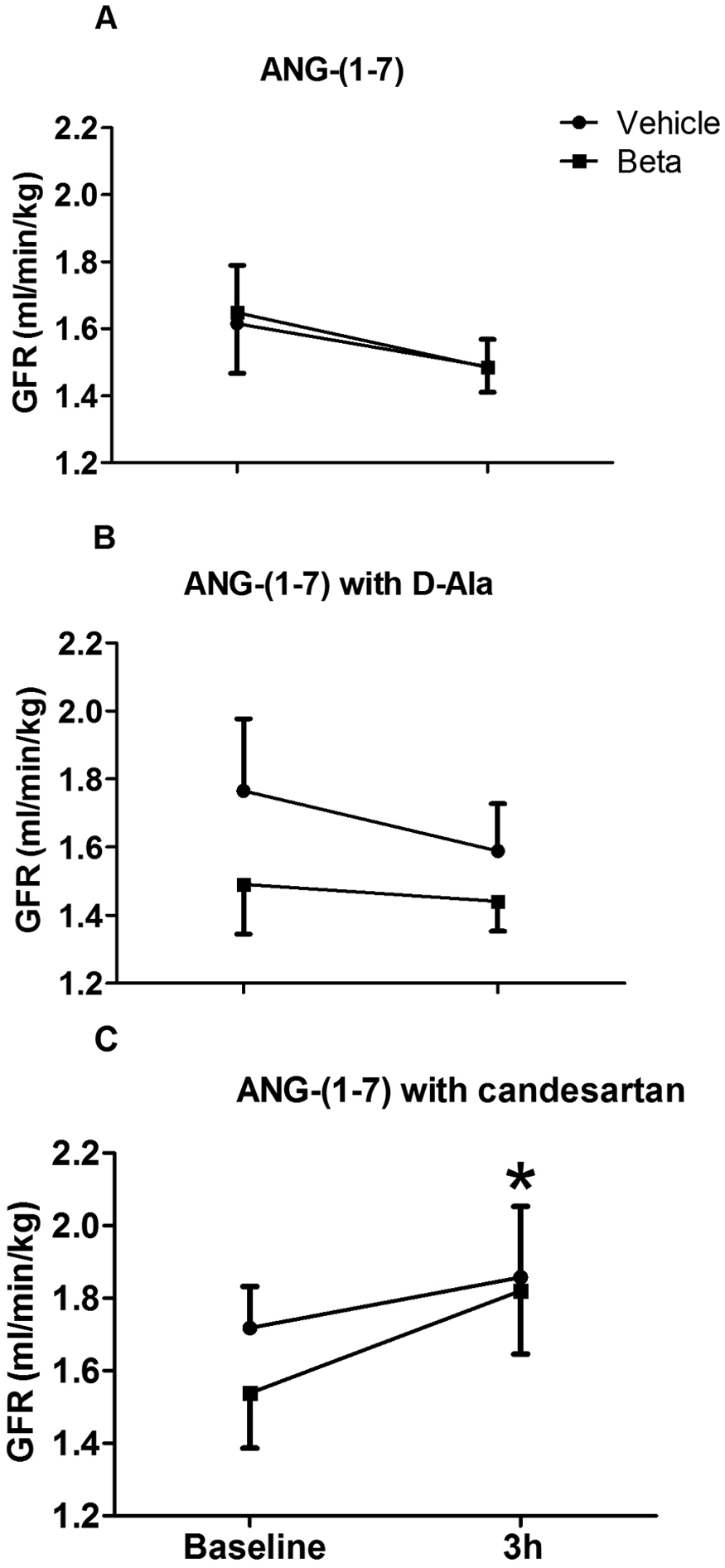
Glomerular filtration rate (GFR) before and after 3 h of intrarenal infusion of (A) angiotensin-(1–7) (ANG-(1–7)) alone, (B) ANG-(1–7) with D-Ala7-ANG-(1–7) (D-Ala), or (C) ANG-(1–7) with candesartan in uninephrectomized vehicle- and betamethasone-treated sheep. *indicates a statistically significant difference within groups after infusion (p<0.05).
Discussion
The present experiments determined whether antenatal betamethasone treatment would reduce the renal responses to direct intrarenal infusions of ANG-(1–7), a heptapeptide thought to counterbalance the effects of the ANG II-AT1R axis. 31 We found that antenatal betamethasone exposure magnified an inhibitory effect of ANG-(1–7) on sodium excretion and that this response was not blocked by D-Ala-ANG-(1–7), a selective antagonist for the Mas receptor for which ANG-(1–7) is a naturally occurring ligand. 32 However, simultaneous intrarenal administration of ANG-(1–7) and the AT1R antagonist candesartan, converted the antinatriuresis to a natriuretic response with higher levels of sodium excretion in the vehicle- than in the betamethasone-treated sheep. Moreover, a reduction in renal plasma flow was observed in both groups along with a tendency for a reduction in GFR following ANG-(1–7) infusion. These responses were also reversed by candesartan but not D-Ala-ANG-(1–7). Considered together, the data suggest that antenatal betamethasone exposure alters the renal responses to ANG-(1–7) and that unilateral nephrectomy modulates these responses independent of any effects induced by antenatal steroid treatment. We chose intrarenal infusion of ANG-(1–7) in an attempt to influence renal function directly and avoid any confounding systemic effects, such as changes in blood pressure, resulting from significant elevations in the peripheral plasma levels of the peptide. Indeed, the intrarenal infusion protocol did not alter circulating levels of ANG-(1–7) nor influence blood pressure.
The data in the literature on the effects of ANG-(1–7) on sodium excretion are conflicting. Multiple reports indicate the peptide increases sodium excretion and /or urine flow in rodents,33–35 dogs 36 and sheep.19,22 Consistent with these findings are in vitro studies that ANG-(1–7) acts on the proximal tubule to inhibit sodium uptake. 32 In contrast, there are several observations that ANG-(1–7) has antidiuretic/antinatriuretic effects in vivo and in vitro. 32 There is reasonable agreement that ANG-(1–7) is a naturally occurring ligand for the Mas receptor and thus many of its effects are mediated via binding to that receptor, 37 but ANG-(1–7) may also interact with AT1 and AT2 receptors. 37 In our studies the inhibition of sodium excretion does not appear to be associated with activation of the Mas receptor as the response was not blocked by D-Ala-ANG-(1–7). The fact that the antinatriuretic effect was converted to a natriuresis by the addition of candesartan implies some interaction with the AT1 receptor which may be opposed by either the Mas or AT2 receptors. This idea is further supported by the changes in the FENa following the different infusions in both groups. Thus, in the presence of AT1R blockade, the ANG-(1–7) induced response converted from a decrease to an increase in FENa. Whether the latter is blocked by D-Ala-ANG-(1–7) or an AT2R antagonist awaits further study, but complex interactions among these receptors and ANG-(1–7) have been noted in studies on the cardiac effects of the peptide.38,39
While we do not have data for sodium excretion before uninephrectomy in this cohort, comparisons with the intact sheep from a different cohort in our previous study 22 reveal some changes in sodium excretion. Specifically, basal sodium excretion in these vehicle-treated uninephrectomized sheep was about twice that found earlier in vehicle-treated intact males. 22 This is consistent with the study by Shirley et al. which showed that excretion rates of sodium were approximately 2-fold higher in the remaining kidney of uninephrectomized rats. 40 Uninephectomy of adult animal results in compensatory renal growth, but does not involve formation of new nephrons. The significant decrease in basal sodium excretion in uninephrectomized, betamethasone-treated sheep (compared to vehicle) strongly suggests that the impaired sodium excretion in this model is directly associated with the antenatal betamethasone exposure while the decrease in sodium excretion caused by ANG-(1–7) seems related to the additional loss of nephrons from the unilateral nephrectomy. Uninephectomy immediately stimulates the growth and function of the remaining kidney. 41 There are differential mechanisms initiating the early compensatory renal growth response and functional adaptation, 42 including a number of endocrine, biochemical, and molecular changes,43,44 which result in glomerular hyperfiltration, renal hypertrophy, and compensatory changes of transporters. 42 Regarding the RAS, ANG II and increased AT1R may mediate the growth of the remaining kidney.45,46 It is unclear whether or not ANG-(1–7) is involved in these changes. However, it is certainly possible that any of the adaptations mentioned above could modulate the responses to betamethasone and ANG-(1–7).
The role of ANG-(1–7) in the regulation of renal plasma flow is not clear. It has been reported that ANG-(1–7) causes afferent arteriolar vasodilatation and, even if devoid of any vasodilator actions by itself, it at least antagonizes the renal vasoconstrictor effects of ANG II. 47 Others, however, have found that ANG-(1–7) does not exert vasodilator and/or ANG II opposing actions in the renal circulation. 48 Although it is difficult to reconcile such contradictory findings, based on their comprehensive analysis of studies regarding the physiologic and pathophysiologic roles of ANG-(1–7), Santos et al. 49 have suggested that the overall activity of the RAS, as it results from experimental manipulations, markedly influences MAP and renal functional responses to ANG-(1–7). Antenatal betamethasone exposure can alter the balance between angiotensin-converting enzyme (ACE) and ACE2 expression implying that the overall activity of the RAS may be changed in the animals in the present study. 50
The intrarenal infusion of ANG-(1–7) suppressed renal plasma flow in both vehicle- and antenatal betamethasone-treated sheep to a similar extent in the absence of changes in MAP. This suppression may have been the result of vasoconstriction via AT1 receptor activation because candesartan, not D-Ala-ANG-(1–7), reversed the suppression.
We have previously reported increases in arterial blood pressure in sheep exposed to betamethasone before birth.8,22,51,52 Consistent with these results, the MAP in antenatal betamethasone-treated sheep is significantly higher than in vehicle-exposed sheep in the present study. The mean difference is similar to that previously reported 51 and would not be accounted for by unilateral nephrectomy in our study. None of the infusions changed MAP in these studies, suggesting that their effects were confined to actions directly on the kidney. Thus, for example, intravenous administration of candesartan lowers MAP in betamethasone-exposed sheep, 52 but our intrarenal infusion had no effect on MAP. Absence of peripheral effects is also supported by the fact that we did not find a significant increase in the plasma levels of ANG-(1–7) during the intrarenal infusions of the peptide.
Basal GFR tended to decrease in antenatal betamethasone-treated sheep but the reduction was not significant. Although the GFR did not significantly change during intrarenal infusion of ANG-(1–7) alone or with D-Ala-ANG-(1–7), renal plasma flow (RPF) decreased. Thus, the decrease in RPF was perhaps caused by contraction of both afferent and efferent arterioles. The GFR increased significantly in both groups during intrarenal infusion of ANG-(1–7) with candesartan. The simultaneous increase in RPF may have contributed to this increase.
GFR values in the present study are different than those we found in intact sheep at 1–2 years of age. In both the vehicle- and betamethasone-treated uninephrectomized animals, absolute GFR values are lower than the corresponding values in intact sheep. 22 The decline in GFR produced by uninephrectomy in these adult sheep agrees with data from other animal studies 53 and with observations in human living kidney donors.54–56 However, single nephron GFR must be increased after the unilateral nephrectomy because the reduction in nephron number was 50% while the decline in GFR was, at most, 25%. It is possible that the increase in single nephron GFR could partially explain the increased sodium excretion in these animals.
In conclusion, the present findings indicate that prenatal exposure to a clinically relevant dose of betamethasone modulates renal responses to ANG-(1–7) in uninephrectomized, adult sheep.
Footnotes
Acknowledgements
The authors would like to thank David Jones and Eric Lesane for providing invaluable support in caring for sheep and assisting with operation and anesthesia, and to thank Xiurong Sun for participating in sample collection and measurement.
Conflicts of interest
None declared.
Funding
This work was supported by National Institutes of Health, Grants HD-47584 and HD-17644.