
Abstract
Lymphangiogenesis is the process of new vessel formation from pre-existing lymphatic vessels. The process mainly involves cell adhesion, migration, and tubule formation of lymphatic endothelial cells. Tumor-induced lymphangiogenesis is an important factor contributing to promotion of tumor growth and cancer metastasis via the lymphatic system. Finding the non-toxic agents that can prevent or inhibit lymphangiogenesis may lead to blocking of lymphatic metastasis. Recently, aspirin, a non-steroidal anti-inflammatory drug (NSAID), has been reported to inhibit in vivo lymphangiogenesis in tumor and incision wound models, but the mechanisms of actions of aspirin on anti-lymphangiogenesis have been less explored. In this study, we aim to explore the mechanism underlying the anti-lymphangiogenic effects of aspirin in primary human dermal lymphatic microvascular endothelial (HMVEC-dLy) cells in vitro. Pretreatment of aspirin at non-toxic dose 0.3 mM significantly suppressed in vitro cord formation, adhesion, and the migration abilities of the HMVEC-dLy cells. Western blotting analysis indicated that aspirin decreased expression of vascular cell adhesion molecule-1 (VCAM-1), at both protein and mRNA levels, and these correlated with the reduction of NF-κB p65 phosphorylation. By using NF-κB inhibitor (BAY-11-7085) and VCAM-1 siRNA, we showed that VCAM-1 expression is downstream of NF-κB activation, and this NF-κB/VCAM-1 signaling pathway controls cord formation, adhesion, and the migration abilities of the HMVEC-dLy cells. In summary, we demonstrate the potential of aspirin as an anti-lymphangiogenic agent, and elucidate its mechanism of action.
Introduction
The lymphatic circulating system is essential for the maintenance of tissue fluid homeostasis, immune cell trafficking and immunosurveillance, as well as absorption of dietary fat and fat-soluble vitamins. Lymphangiogenesis represents new vessel formation from pre-existing lymphatic vessels. Pathological lymphangiogenesis is promoted under some pathological conditions such as inflammation and cancer metastasis. 1 Tumors can induce lymphangiogenesis and use the lymphatic circulatory system to spread out from the primary tumor to distant sites. 2 Owing to the absence of basal lamina and lack of associated pericytes, lymphatic capillaries are the major route for metastasis of some solid tumor types. 3 Lymphangiogenic inducer, such as vascular endothelial growth factors (VEGFs), stimulates the lymphatic endothelial cells (LECs) to proliferate, degrade extracellular matrix, and migrate toward the gradient of stimulus, followed by association and reorganization of the LECs into tube-like structures.4,5 Understanding the mechanisms regulating lymphangiogenesis may be useful in the development of antimetastasis therapy.
Cell adhesion to extracellular matrix (ECM) and cell migration are important processes that are involved in the lymphangiogenesis. 2 Vascular cell adhesion molecule-1 (VCAM-1) has been shown to play essential roles in lymphatic cell adhesion and lymphatic development in zebrafish embryos by interaction with integrin α9 (Itgα9) 6 and is also involved in lymph node metastasis in mice by promoting adhesion of tumor cells to LECs through interaction with integrin α4β1. 7 Our previous microarray analysis found that VCAM-1 is the most upregulated gene in rat LECs during the early stage of in vitro cord formation on Matrigel. 8 However, the molecular mechanisms controlling VCAM-1 expression during lymphangiogenesis are not well understood.
Nuclear factor-kappaB (NF-κB) is a ubiquitous transcription factor which rapidly induces activation in endothelial cells. 9 VCAM-1 is downstream of NF-κB activation which involves protein kinase C (PKC) and p38 mitogen activated protein kinase (p38 MAPK) signaling pathways. 10 NF-κB binds to VCAM-1 promoter and drives expression of the gene 11 ; however, the VCAM-1 expression via NF-κB activation is cell-type specific. 12 Until now, the regulatory mechanism for VCAM-1 expression in lymphangiogenesis is still poorly understood.
Aspirin, an extensively used non-steroidal anti-inflammatory agent (NSAID), exerts its anti-inflammatory, anti-thrombotic, anti-pyrolic and analgesic effects through inhibiting cyclooxygenase (COX) enzymes.13,14 Additionally, aspirin can inhibit NF-κB activation in several models. 15 Recently, several lines of evidence revealed that taking a daily low-dose aspirin could reduce the incidence of certain types of cancer, such as colon cancer, esophageal cancer, breast cancer, and hormone-related cancers.16–19 Low-dose aspirin can inhibit cancer-induced angiogenesis in several models. 20 Additionally, taking high-dose aspirin led to a lower risk of prostate cancer-specific mortality, compared with low-dose users. 21 It is of much interest to use aspirin in cancer prevention and antimetastasis therapy due to its safe long-term treatment, which is a requirement for antimetastatic drugs. However, the dose for use in cancer treatment still needs to be optimized. 22 Interestingly, a recent report showed that daily oral administration of aspirin significantly decreased lymph node metastasis of lung cancer cells and improved the overall survival of the treated mice. 23 However, there is no detailed report on the inhibitory effect of aspirin on lymphangiogenesis and its mechanism.
Here, we report, for the first time: (1) aspirin at non-toxic dosage levels inhibited lymphangiogenesis in vitro; (2) VCAM-1 gene expression was downstream of NF-κB activation which controls lymphangiogenesis in vitro; and (3) anti-lymphangiogenic activity of aspirin was mediated by the inhibition of the NF-κB/VCAM-1 pathway. This finding may be useful for understanding the preventive effect of aspirin on lymphangiogenesis as well as cancer metastasis.
Materials and methods
Materials
Aspirin was purchased from Sigma-Aldrich (St Louis, MO, USA). The compound was dissolved in dimethylsulfoxide (DMSO) to make a stock solution. The final concentration of DMSO was kept below 0.5% throughout the studies. Matrigel was purchased from BD Biosciences (San Diego, CA, USA). Polyclonal antibody specific to VCAM-1 was obtained from Cell Applications (San Diego, CA, USA). Primary antibodies specific to NF-κB p65 and its phosphorylated form (Ser536) were obtained from Cell Signaling Technology (Beverly, MA, USA). Horseradish peroxidase-linked secondary antibodies were obtained from Cell Signaling Technology and Santa Cruz Biotechnology (Santa Cruz, CA, USA). Polyclonal antibody specific to proliferating cell nuclear antigen (PCNA), VCAM-1 siRNA (sc-29519), and control siRNA (sc-37007) were purchased from Santa Cruz Biotechnology. NF-κB inhibitor (BAY 11-7085) was purchased from Calbiochem (San Diego, CA, USA).
Endothelial cells
Primary human dermal lymphatic microvascular endothelial (HMVEC-dLy) cells were obtained from Lonza (Walkersville, MD, USA). The cells were characterized by the expression of lymphatic markers PROX-1, LYVE-1 and Podoplanin using reverse transcription-polymerase chain reaction (RT-PCR). The cells were maintained in EGM-2MV Bullet Kit (Lonza), at 37°C in a humidified atmosphere of 5% CO2 in air, and sub-cultured with Trypsin-EDTA solution every 5 days. Only cells from passages 5 to 15 were used in the study.
Cell proliferation assay
Cell viability after treatment was determined by the WST-8 cell proliferation assay kit (DOJINDO, Kumamoto, Japan). HMVEC-dLy cells were cultured in 96-well plates (1.5×104 cells/well), at 37°C for 24 hours. Then the cells were treated with various concentrations of aspirin, and further incubated for 48 hours. After that, 10 µL WST-8 cell proliferation assay kit was added to each well, and incubated continuously at 37°C for 2 hours, then the absorbance was measured at 450 nm to determine the number of viable cells.
Cord formation on Matrigel
Ninety-six-well plates were coated with 60 µL Matrigel (10 mg/mL) and left to polymerize at 37°C. HMVEC-dLy cells were cultured in medium containing 0.3 mM for 48 hours, then harvested by trypsinization and the cell suspension seeded into the Matrigel-coated wells (8×103 cells/well), and further incubated at 37°C. At each time point, cells were fixed with a 4% paraformaldehyde fixative and stained using Mayer’s hematoxylin (Muto Pure Chemical, Tokyo, Japan). The characteristics of the cord network were photographed and the total length of cords was measured using a software Angiogenesis Image Analyzer (Kurabo, Osaka, Japan) and calculated as a percentage compared to control.
Cell adhesion assay
HMVEC-dLy cells were cultured in medium containing 0.3 mM for 48 hours, then harvested by trypsinization and the cell suspension seeded into a 96-well plate (2×104 cells/well), which had been pre-coated with 100 µg/mL Matrigel, and further incubated at 37°C for 30 minutes. Unattached cells were washed out from wells with phosphate buffered saline (PBS). The remaining attached cells were fixed with a 4% paraformaldehyde fixative for 30 minutes and stained with 0.5% crystal violet in 20% methanol for 5 minutes. After washing and air drying, the total number of attached cells per well was counted and calculated as a percentage compared to control.
Cell migration assay
This assay was performed in Falcon cell culture PET inserts with a pore size of 8 µm in a 24-well format (no. 353097; Falcon, Franklin Lakes, NJ, USA). Briefly, the lower surface of the filters was coated with 5 µg/mL Matrigel. HMVEC-dLy cells were cultured in medium containing 0.3 mM for 48 hours, then harvested by trypsinization and the cell suspension was added into the upper compartment of the chamber (1.5×103 cells/100 µL), while the lower compartment was filled with 600 µL normal culture medium. The cells in chambers were incubated for 24 hours at 37°C. Non-migrated cells were removed with cotton swabs. The migrated cells attached on the filter were fixed with methanol, stained with Mayer’s hematoxylin and 1% eosin Y solution (Muto Pure Chemical), and photographed. The total number of migrated cells per culture insert was counted and calculated as a percentage compared to control.
Reverse transcription-polymerase chain reaction (RT-PCR) and quantitative RT-PCR (qRT-PCR)
Total RNA was extracted from the cells cultured on Matrigel using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Subsequently, the amount of 2 µg of total RNA was reverse transcribed into cDNA. The PCR products for the reverse transcription-polymerase chain reaction (RT-PCR) were electrophoresed on 1.5% agarose gels and stained with SYBER Green I Nucleic Acid Gel Staining (Lonza, Rockland, ME, USA). The PCR reaction for quantitative RT-PCR (qRT-PCR) was performed by using the StepOnePlus™ (Applied Biosystems, Waltham, MA, USA) according to the manufacturer’s instructions. GAPDH was used as an internal control. The relative quantification of mRNA expression was calculated as a ratio of the target gene to GAPDH. The primer sequences were as follows: VCAM-1 sense, 5′-CGTCTTGGTCAGCCCTTCCT-3′, and antisense, 5′-ACATTCATATACTCCCGCATCCTTC-3′; LYVE-1 sense, 5′-GTGCTTCAGCCTGGTGGTG-3′, and antisense, 5′-GCTTGGACTCTTGGACTCTTC-3′; PROX-1 sense, 5′-TTCAGATGGAGAAGTACGCA-3′, and antisense, 5′- GGACTGCTACTCTTCATACA-3′; Podoplanin sense, 5′-CAACAACTCAACGGGAACGATG-3′, and antisense, 5′-GAGCTCAGGGACAGGGCACAGA-3′; GAPDH sense, 5′-AGCCACATCGCTCAGACAC-3′, and antisense, 5′-GCCCAATACGACCAAATCC-3′.
Western blotting analysis
To determine protein levels after seeding the cells on a Matrigel-coated dish, western blotting was performed. Cells were washed with PBS, incubated with RIPA lysis buffer (Santa Cruz Biotechnology, Santa Cruz, CA, USA) and then scraped. The collected samples were vigorously vortexed, and centrifuged at 14,000 rpm for 10 minutes, then the supernatant was collected. For western blotting analysis, the lysate in equal amount was resuspended in loading buffer, boiled for 3 minutes, separated on 7.5–10% SDS-polyacrylamide gels, and transferred to a polyvinylidene difluoride (PVDF) membrane. The membrane was incubated with specific antibodies. The bands were detected by using the immunochemiluminescence technique. PCNA was used as loading controls.
siRNA transfection
Proliferating HMVEC-dLy was transfected with control siRNA or siRNA against VCAM-1 (Santa Cruz Biotechnology) at a final concentration of 6 nM using Lipofectamine RNAiMAX reagents (Invitrogen). After transfection, the cells were grown for 18 hours at 37°C in 5% CO2 and trypsinized. The transfected cells were seeded on Matrigel-coated dishes and then employed for cord formation assay, western blotting analysis, and qRT-PCR.
Statistical analysis
Statistical significance was assessed by one-way ANOVA, with Dunnett’s test and by Student’s t-test. P-values < 0.05 were considered to be significant.
Results
Effect of aspirin on HMVEC-dLy cell growth
Aspirin has been reported to inhibit lymphangiogenesis in several tumor-induced lymphangiogenesis models, 3 but the mechanisms underlying this anti-lymphangiogenic effect have not been investigated. Thus, the non-cytotoxic concentration ranges of aspirin were first explored, in order to find the concentrations that can inhibit lymphangiogenesis without affecting proliferation of LECs. HMVEC-dLy cells which were characterized by the expression of lymphatic markers PROX-1, LYVE-1 and Podoplanin using RT-PCR (Figure 1), were treated in culture medium containing aspirin at various concentrations for 48 hours. The results showed that aspirin concentrations lower than 0.31 mM did not affect proliferation of the cells (Figure 2). Therefore, the highest non-toxic concentration of aspirin (0.3 mM) was selected for further studies.
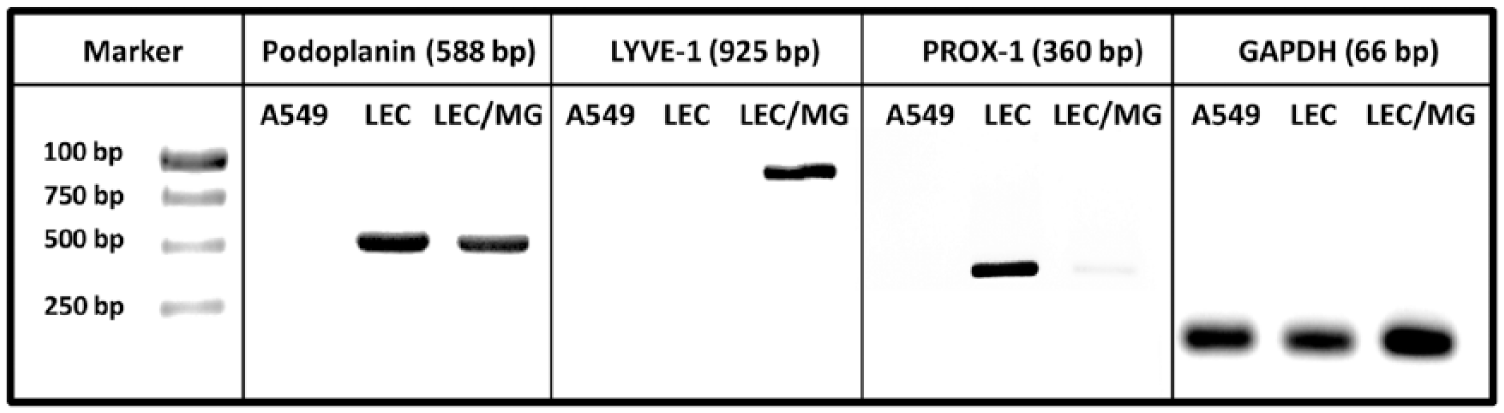
Cell characterization. The HMVEC-dLy cells were confirmed by the expression of lymphatic markers LYVE-1, PROX-1 and Podoplanin using RT-PCR after culture with (LEC/MG) and without Matrigel (LEC). GAPDH was employed as the loading control. The human lung carcinoma (A549) was used as a negative control.
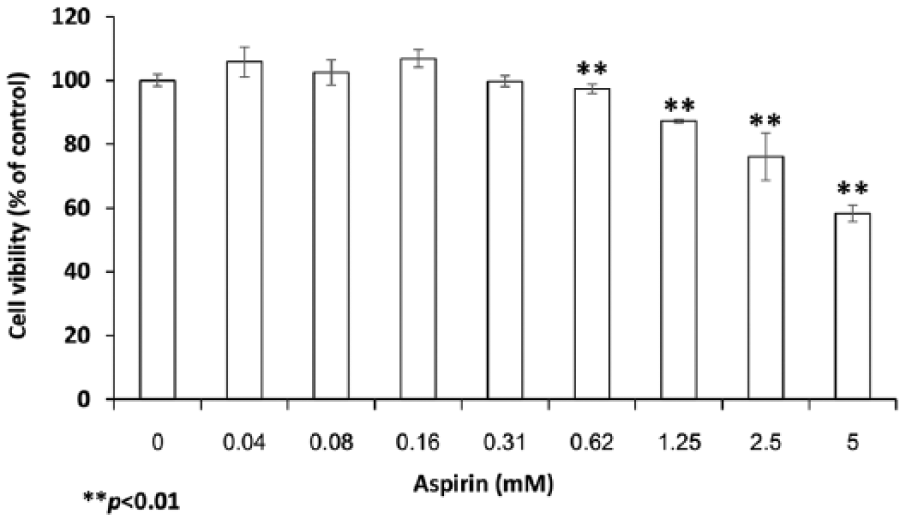
The effect of aspirin on growth of HMVEC-dLy cells. HMVEC-dLy cells were incubated with 0–5 mM aspirin in a 96-well plate for 48 hours. After that, cell viability was determined by WST-8 assay. Data are shown with the mean ± SD (**p < 0.01) compared with control (0.5% DMSO).
Aspirin suppresses cord formation of HMVEC-dLy cells
To investigate the direct effect of aspirin on lymphangiogenesis in vitro, the cord formation assay was performed. The cells were exposed to aspirin at 0.3 mM (0.03% DMSO) for 48 hours, after which the cells were trypsinized and then seeded onto Matrigel. Cord formation of HMVEC-dLy at 2, 4, 6, and 8 hours was observed, and photographed (Figure 3A). Total cord length was measured. The aspirin-treated cells could not form cord networks as well as the control group (Figure 3B).
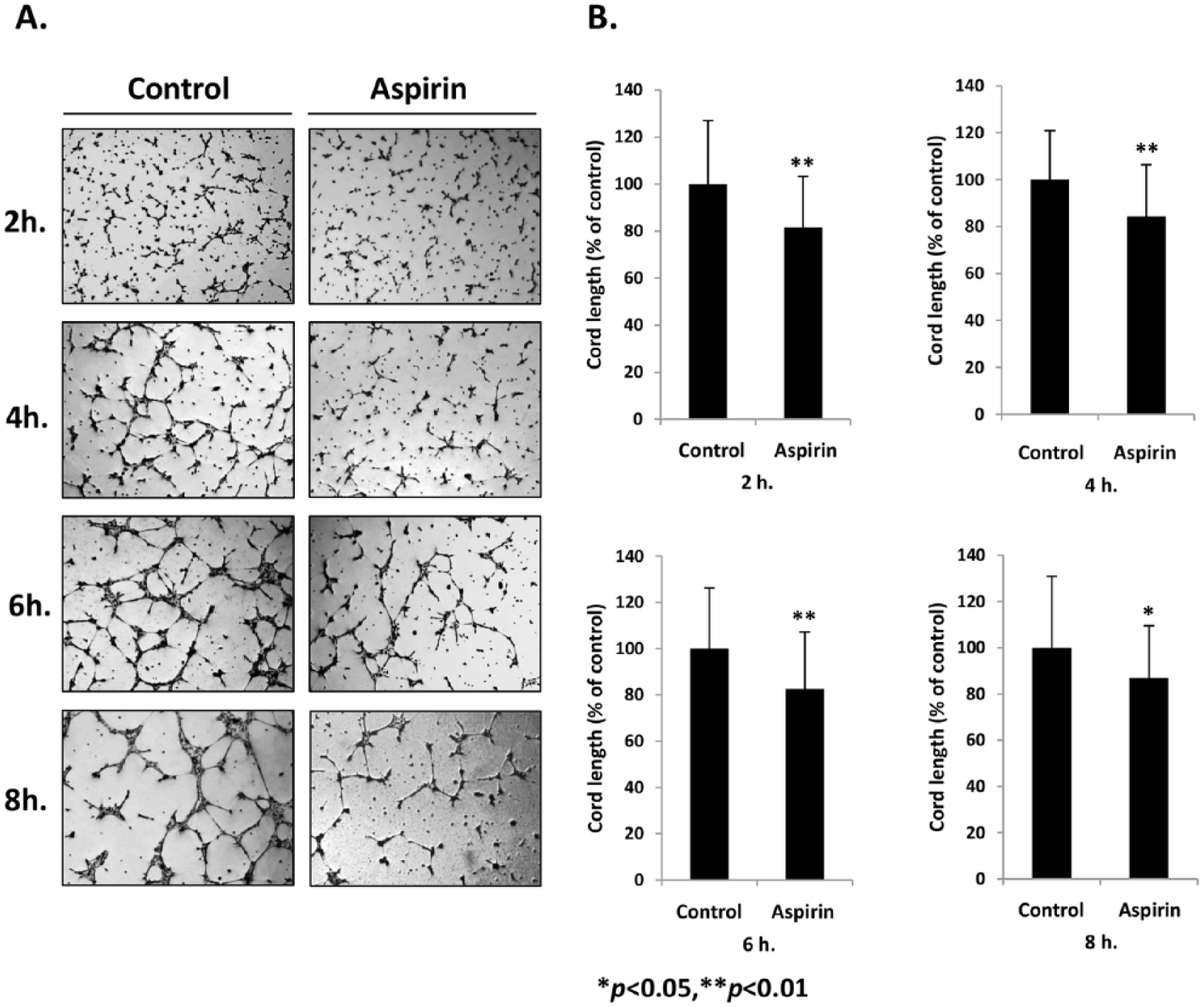
The effect of aspirin on cord formation of HMVEC-dLy cells. The cells were cultured in the presence of 0.3 mM aspirin for 48 hours, then seeded on Matrigel and further cultured with normal culture medium. At the indicated time, photographs were taken (A). Total cord length was measured using image analysis software, and presented as percentage compared to control (B). Data are shown as mean ± SD of three independent experiments; *p < 0.05 and **p < 0.01 compared with control (0.03% DMSO).
Aspirin suppresses adhesion and migration of HMVEC-dLy cells
We further examined whether aspirin affected cell-ECM adhesion and the cell migration abilities of LECs. The HMVEC-dLy cells were cultured in medium containing 0.3 mM aspirin for 48 hours, and then subjected to cell adhesion and migration assays in normal culture medium. The results showed that aspirin significantly decreased cell adhesion and cell migration abilities by 24.2% and 24.5% inhibition, respectively, compared with control (0.03% DMSO) (*p<0.05) (Figure 4). The results suggested that the in vitro anti-lymphangiogenic effect of aspirin involved a decrease in cell adhesion and the migration abilities of LECs.
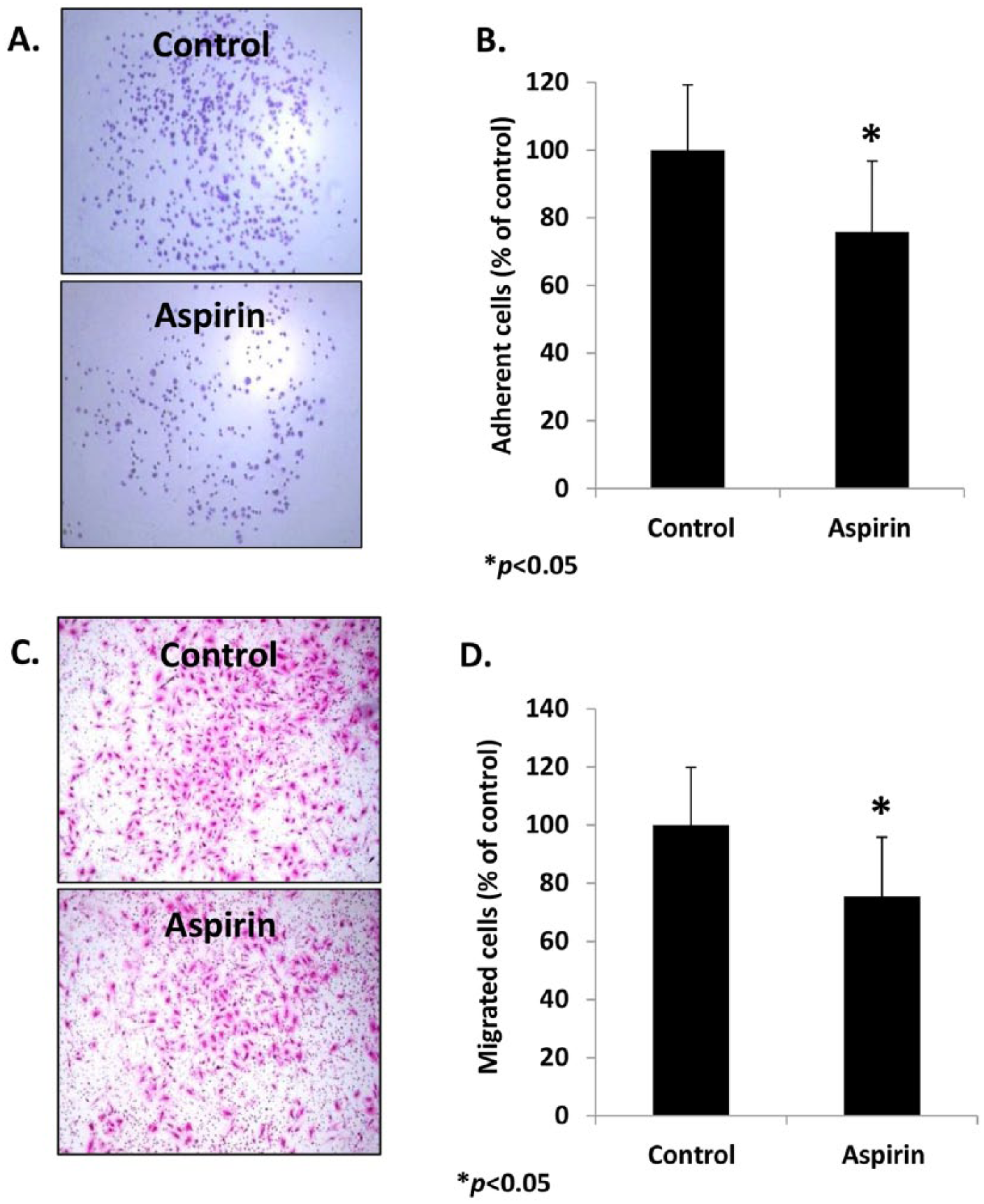
The effect of aspirin on ECM-cell adhesion and migration of HMVEC-dLy cells. The cells were cultured in the presence of 0.3 mM aspirin for 48 hours, and then subjected to cell adhesion and cell migration (Transwell) assays. Photographs of attached cells (A) and migrated cells (C) were taken and the number of cells was counted. Percentage of cell adhesion (B) and cell migration (D) are shown as mean ± SD of three independent experiments; *p < 0.05 compared with control (0.03% DMSO). (Note: Figure is in color online.)
Aspirin suppresses VCAM-1 expression in HMVEC-dLy cells during cord formation
VCAM-1 is known to play roles in cell adhesion and migration of LECs, as well as lymphatic development.6,24 Moreover, VCAM-1 was also found to be the most upregulated gene during early stages of cord formation in rat LECs on Matrigel. 8 The effect of aspirin on VCAM-1 expression in LECs was explored in HMVEC-dLy cells during cord formation. After culturing cells in medium containing 0.3 mM aspirin for 48 hours, the cells were harvested and seeded onto Matrigel. At 4 hours after seeding, the VCAM-1 protein levels in aspirin-treated cells were markedly decreased compared with control (0.03% DMSO) (Figure 5A). We further investigated whether the decreased protein levels were due to downregulation of VCAM-1 gene expression; the qRT-PCR analysis revealed that the VCAM-1 mRNA level in aspirin-treated cells was significantly reduced by 0.64-fold compared with the control (0.03% DMSO) (Figure 5B). These results suggest that aspirin treatment could downregulate VCAM-1 gene expression during cord formation of HMVEC-dLy cells.
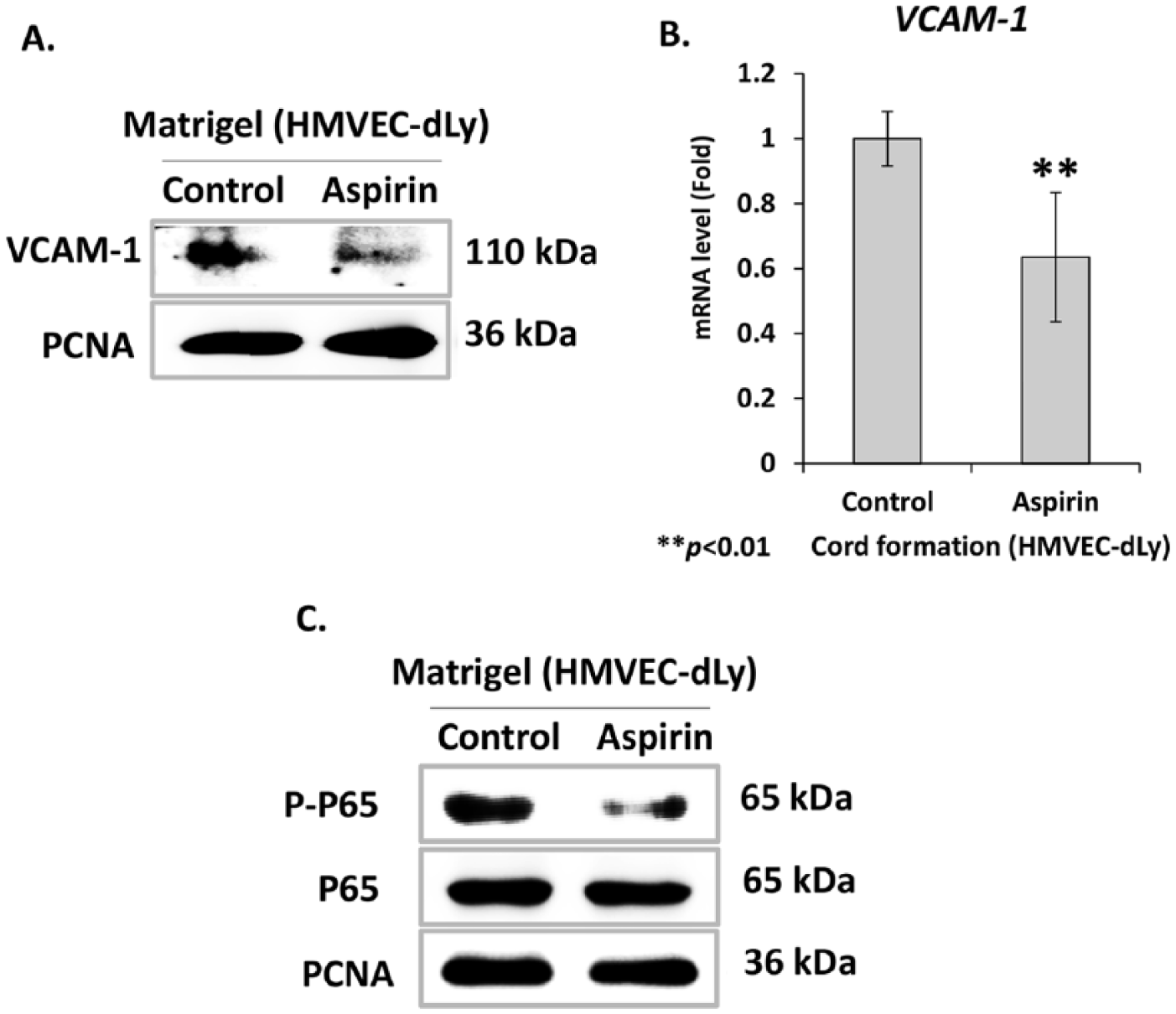
The effect of aspirin on VCAM-1 expression and in HMVEC-dLy cells during cord formation. Cells were cultured in the presence of 0.3 mM aspirin for 48 hours, then seeded on Matrigel and further cultured for 4 hours with normal culture medium. Expression levels of VCAM-1 protein (A) and mRNA (B) were detected by using western blotting analysis and qRT-PCR, respectively. Activation of NF-κB signaling (C) was detected by using western blotting analysis. Phosphorylated p65 (P-p65) is the active form of NF-κB. PCNA was used as a loading control. Data are shown as mean ± SD of three independent experiments; **p < 0.01 compared with control (0.03% DMSO).
Aspirin inhibits NF-κB activation in HMVEC-dLy cells during cord formation
It has been reported that VCAM-1 is an NF-κB dependent gene,11,25 and aspirin can inhibit the function of NF-κB by interfering with the pathway that leads to inhibitor of kappa B (I-κB) degradation and phosphorylation of NF-κB p65 subunits.15,26 It is therefore possible that the decrease of VCAM-1 expression by aspirin in HMVEC-dLy cells may result from blocking of NF-κB activation during cord formation. Thus, we determined the levels of NF-κB p65 phosphorylation in the aspirin-treated cells during cord formation. The cells were treated with 0.3 mM aspirin for 48 hours, then seeded and cultured on Matrigel for 4 hours. The result showed a dramatic decrease in levels of NF-κB p65 phosphorylation in the aspirin-treated cells compared with control (0.03% DMSO) (Figure 5C). The results indicated a correlation between the inhibition of NF-κB activation and downregulation of VCAM-1 gene expression in aspirin-treated HMVEC-dLy cells.
Cord formation, adhesion and migration of HMVEC-dLy cells is controlled by NF-κB/VCAM-1 pathway
We used siVCAM-1 transfection and NF-κB inhibitor (BAY 11-7085) to confirm the role of the NF-κB/VCAM-1 pathway in lymphangiogenesis of HMVEC-dLy cells. Firstly, VCAM-1 expression in HMVEC-dLy cells was silenced by using the siRNA technique. After the transfected cells were cultured on Matrigel for 4 hours, mRNA and protein levels of VCAM-1 in siVCAM-1-transfected cells were markedly reduced (Figures 6A and 6B), while NF-κB p65 phosphorylation level was not affected (Figure 6C), suggesting that VCAM-1 expression is downstream of NF-κB activation. The siVCAM-1-transfected cells showed significantly reduced abilities to form cord networks, adhesion and migration by 62.8%, 60.0% and 38.1%, respectively, compared with siControl-transfected cells (Figures 6D, 6F and 6G) without the reduction of cell viability (Figure 6E). These results confirm the important role of VCAM-1 in lymphangiogenesis of HMVEC-dLy cells.
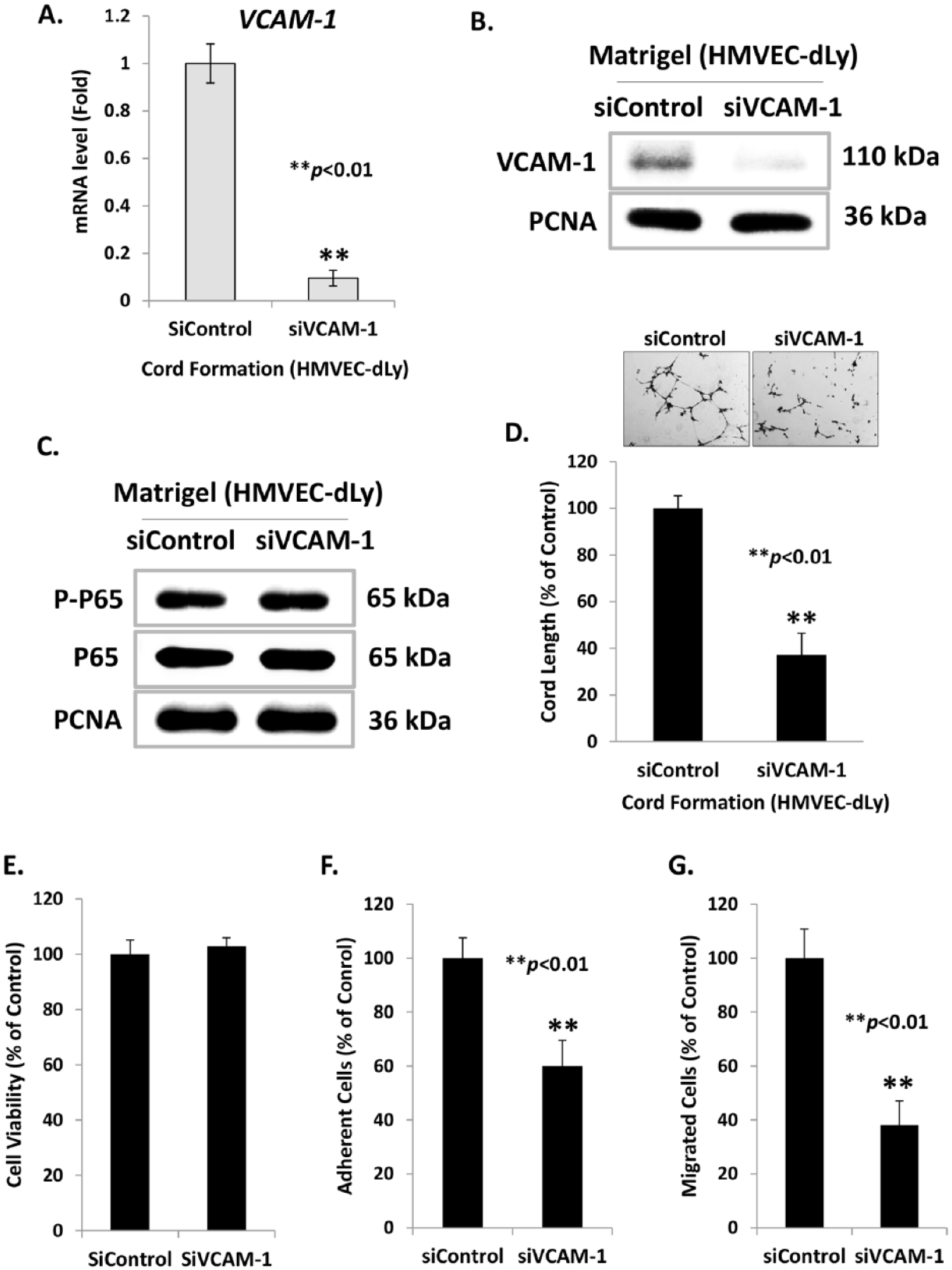
The effect of VCAM-1 silencing on the NF-κB activation and cord formation ability of HMVEC-dLy cells. After siRNA transfection, the transfected cells were cultured on Matrigel for 4 hours. Then the cells were subjected to extract mRNA and protein and determined the levels of VCAM-1 mRNA by qRT-PCR (A), VCAM-1 protein (B) and NF-κB (phosphorylated and total forms of p65) protein by western blotting analysis (C), cord formation assay (D), proliferation assay (E), adhesion assay (F), and migration assay (G). Data are shown as mean ± SD of three independent experiments; **p < 0.01 compared with control siRNA group.
To further investigate the possible involvement of NF-κB in lymphangiogenesis of HMVEC-dLy cells, the cells were treated with an inhibitor of NF-κB activation (BAY 11-7085) for 1 hour before culturing on Matrigel for 4 hours. The results showed that NF-κB inhibitor treatment successfully decreased the level of phosphorylated NF-κB p65 protein compared with control (0.005% DMSO) (Figure 7A), and this suppression correlated with the significant reduction of VCAM-1 mRNA and protein levels compared with control (0.005% DMSO) (Figures 7B and 7C). Finally, the NF-κB inhibitor also significantly suppressed cord formation, adhesion and the migration abilities of HMVEC-dLy cells by 52.4%, 55.1% and 24.1% inhibition, respectively, compared with control (0.005% DMSO) (Figures 7D, 7F and 7G) without the reduction of cell viability (Figure 7E). Taken together, these results revealed that NF-κB is an upstream regulator of VCAM-1 gene expression, and also confirmed the important role of the NF-κB/VCAM-1 pathway in lymphangiogenesis.
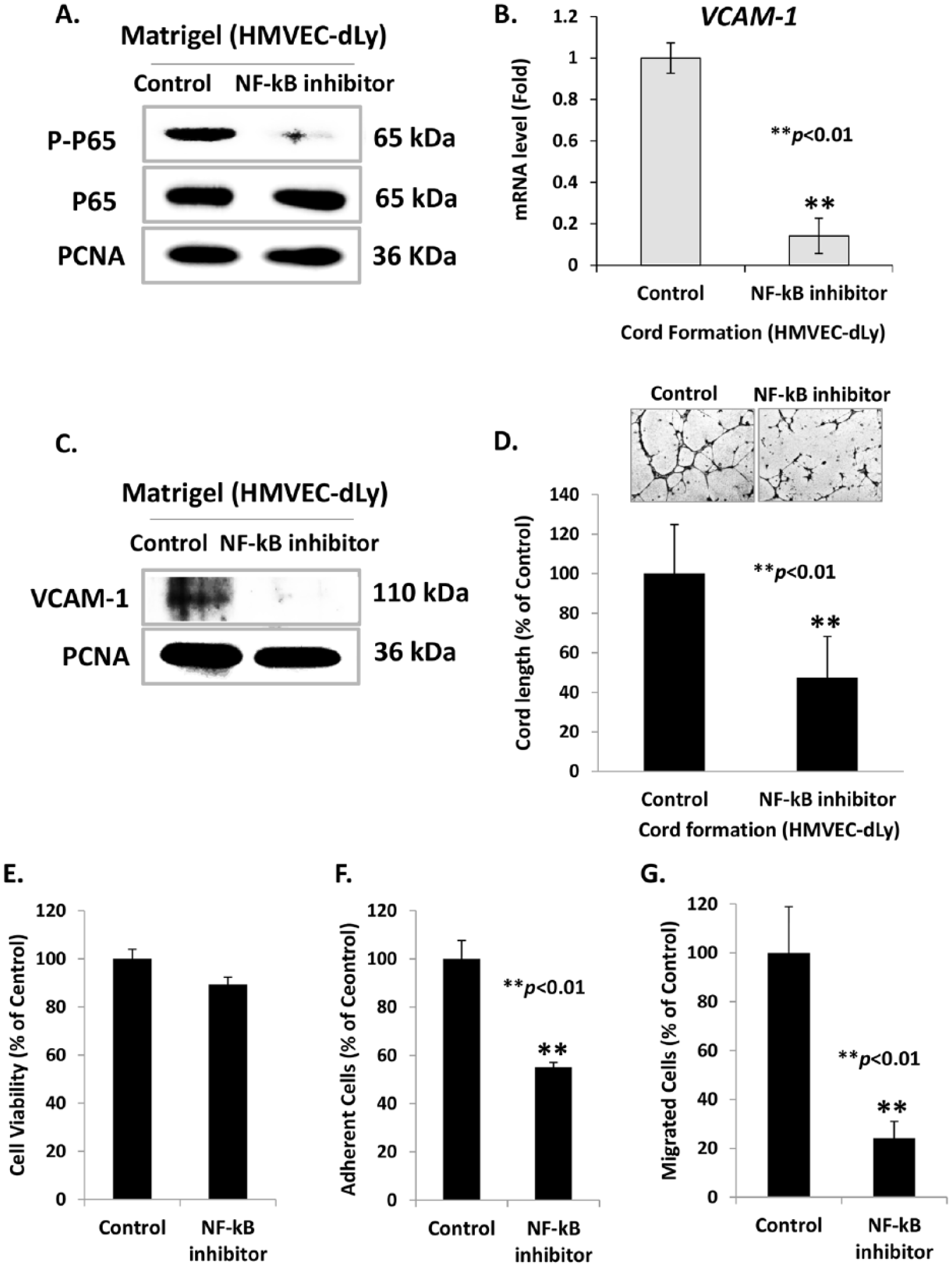
The effect of NF-κB specific inhibitor on VCAM-1 expression and cord formation ability of HMVEC-dLy cells. The cells were pretreated with 5 µM of NF-κB inhibitor (BAY 11-7085) for 1 hour, then seeded on Matrigel and cultured for 4 hours. Activation of NF-κB signaling, P-p65 levels (A) and VCAM-1 protein levels (C) were detected by western blotting analysis. VCAM-1 mRNA levels were determined by qRT-PCR (B). The cells were also subjected to cord formation assay (D), proliferation assay (E), adhesion assay (F), and migration assay (G). Data are shown as mean ± SD of three independent experiments; **p < 0.01 compared with control (0.005% DMSO).
Discussion
Tumor-induced lymphangiogenesis is an important phenomenon that supports growth and metastasis of solid tumors. 4 As far as we know, this is the first report showing the anti-lymphangiogenic effect of aspirin and its mechanism of action in human LECs.
The in vivo anti-lymphangiogenic effect of aspirin has been demonstrated. Aspirin inhibited lymphatic regeneration in a rat model of surgical wound incision. 27 Daily oral administration of aspirin could suppress growth of S180 sarcoma tumors in mice, as well as decrease the lymphatic vessel density in the tumors, and these effects correlate with the reduction of VEGF-C protein levels in the tumors of aspirin-treated mice compared with control mice. 28 VEGF-C is a lymphangiogenic inducer directly acting on LECs. Cancer cells can secrete VEGF-C to promote lymphangiogenesis as well as lymphatic metastasis. 29 In the latter model, the mechanism of action suggested was inhibition of VEGF-C production in the tumors by aspirin, which might involve the COX-2 inhibitory effect of aspirin. 28
In this study, we investigated the possible mechanisms of aspirin by starting from the upregulation of VCAM-1 during cord formation of LECs. VCAM-1 is a member of the immunoglobulin gene superfamily and has been described as a cytokine-inducible endothelial adhesion molecule. 30 We previously reported microarray analysis data indicating the VCAM-1 gene was the most upregulated gene during the early stage of lymphangiogenesis of rat LECs (TR-LE cell line) on Matrigel. 8 Similarly, we also found increased VCAM-1 expression in HMVEC-dLy cells during cord formation on Matrigel (data not shown). Our results indicated that VCAM-1 expression was important for the process of lymphangiogenesis in vitro, and that VCAM-1 induction required activation of the NF-κB signaling pathway. The key regulating step in NF-κB signaling activation is the dissociation of the cytosolic I-κB/NF-κB complex to release NF-κB proteins which further translocate to the nucleus. Binding of the activated NF-κB to the VCAM-1 promoter region is sufficient to drive VCAM-1 gene expression.11,25 Aspirin suppresses activation of NF-κB signaling by inhibiting IKK-β activity, 31 which is necessary for both I-κB degradation and NF-κB p65 phosphorylation. 32 The effect of aspirin on the expression of adhesion molecules seems to be selective suppression of VCAM-1 expression in HMVEC-dLy. We have also tried to see the effect of aspirin on the expression of vascular endothelial-cadherin (VE-cadherin), the major endothelial-specific cell adhesion molecule that plays an important role in vascular morphogenesis and growth control 33 in HMVEC-dLy, and found that aspirin did not change the amount of VE-cadherin protein (data not shown). In addition, it has been reported that aspirin inhibited NF-κB-dependent E-selectin and VCAM-1 induction by tumor necrosis factor-α (TNF-α) but did not have an effect on intercellular adhesion molecule-1 (ICAM-1) in human umbilical vein endothelial cells (HUVECs).26,34 Therefore, the mechanism of aspirin on the selective inhibition of adhesion molecules of HMVEC-dLy cells will be further studied. Taken together, this is consistent with our results, showing that aspirin decreases VCAM-1 expression through inhibition of NF-κB p65 phosphorylation, resulting in suppression of cord formation of LECs. This mechanism may underlie the anti-lymphangiogenic activity of aspirin as depicted in Figure 8.
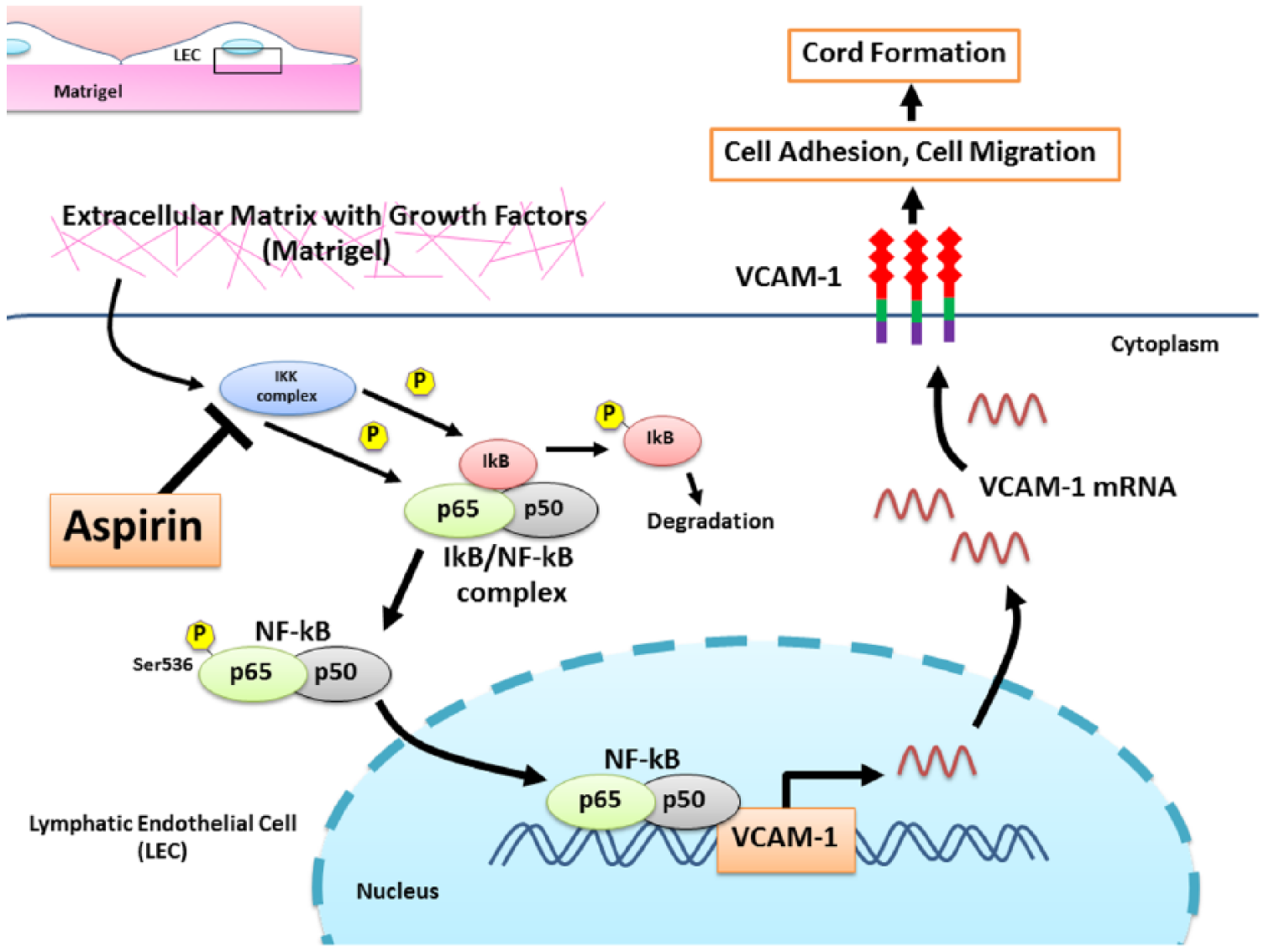
A proposed mechanism underlying anti-lymphangiogenic activity of aspirin in HMVEC-dLy cells. When the cells receive signals from its microenvironment, such as cytokines and growth factors presented in Matrigel, NF-κB signaling is activated and the active NF-κB proteins translocate into the nucleus where they upregulate expression of the VCAM-1 gene. The increased VCAM-1 levels will enhance the cord formation ability of the cells. Aspirin suppresses in vitro lymphangiogenesis by inhibiting NF-κB activation, thus resulting in decreased VCAM-1 expression and reduced cord formation, adhesion and migration abilities of the cells. HMVEC-dLy, human dermal lymphatic microvascular endothelial cells; NF-κB, nuclear factor-kappaB; VCAM-1, vascular cell adhesion molecule-1. (Note: Figure is in color online.)
Conclusion
We show the potential of aspirin in inhibiting lymphangiogenesis of human LECs, and also elucidated its mechanism of action which occurs through inhibition of the NF-κB/VCAM-1 signaling pathway. This finding suggests the additional benefit of aspirin in suppression of pathological lymphangiogenesis, such as inhibiting tumor-induced lymphangiogenesis.
Footnotes
Acknowledgements
We acknowledge the Laboratory of Biochemistry, Chulabhorn Research Institute, Thailand, the Research Center for Drug Discovery and Development, Faculty of Pharmacy, Srinakharinwirot University, Thailand, and the Department of Kampo Diagnostics, University of Toyama, Japan for providing various reagents, facilities and equipment.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: this work was fully funded by the Thailand Research Fund under grant number MRG5980210.