
Abstract
Previously, we created an experimental murine model for the induction of vulnerable plaque (VP). Although this murine model offers the opportunity to study the different molecular biological pathways that regulate plaque destabilization, the size of the animals severely limits the use of the model for in vivo diagnostics and percutaneous interventions. This study aimed to create a VP model in the rabbit, based on the murine model, to aid the assessment and development of novel diagnostic and interventional tools. New Zealand white rabbits were fed on a 2% cholesterol diet. After 1 week, a shear stress-altering device was implanted around the right carotid artery. Twelve weeks after cast placement, the carotid artery was isolated and processed for (immuno-)histological analysis to evaluate the presence of a VP phenotype. Atherosclerotic plaques with high lipid and macrophage content, low vascular smooth muscle cell content and intimal neovascularization were located upstream and downstream of the cast. The plaques lacked a significant necrotic core. In conclusion, we were able to create atherosclerotic plaques with a phenotype beyond that of a fatty streak, with a high percentage of lipids and macrophages, a thick cap with some vascular smooth muscle cells and neovascularization. However, as there was only a small necrotic core, the overall phenotype seems less vulnerable as compared to the thin fibrous cap atheroma in patients.
Introduction
Wall shear stress is a hemodynamic force that plays an important role in the induction and preservation of endothelial cell homeostasis during vascular development and in the mature vascular system. In straight arterial segments where unidirectional laminar flow prevails, average shear stress levels are actively maintained to trigger an atheroprotective expression profile in endothelial cells. In contrast, initial areas of atherosclerosis development correlate strongly with specific sites in the vascular structure where the endothelium is constantly exposed to relatively low and oscillatory shear stress.1 –4
Previously, we have shown in an in vivo study that lowering shear stress by placement of a tapered perivascular cast around the common carotid artery of an apolipoprotein E-deficient (ApoE–/–) mouse promotes atherosclerotic plaque development in the upstream and downstream vessel segment. 5 Upstream, these lesions showed characteristics of a human vulnerable plaque, the culprit atherosclerotic lesions that are associated with rupture in patients and are defined by a distinct morphology, which include the presence of a thin fibrous cap, large necrotic core and high inflammatory state. In contrast, in the downstream vessel region where oscillatory shear stress was induced, atherosclerotic lesions with a more stable phenotype were formed with a thick fibrous cap, neglectable necrotic core, and low infiltration of inflammatory cells. This murine cast model offered the ideal setting to study different molecular biological pathways that regulate plaque formation and destabilization, as the effect of interventions on the two specific types of plaques could be compared within one single carotid artery. Subsequent studies indeed provided important insights in the pathogenesis of the disease.6 –9 However, the small size of the animals severely limits the use of this model for in vivo diagnostics and percutaneous interventions.
Currently, rabbits are widely used to study atherosclerosis as they are sensitive to a high cholesterol diet, leading up to the accumulation of high levels of total plasma cholesterol and the subsequent development of atherosclerotic lesions.10,11 Furthermore, it has been shown that rabbits can be used for in vivo intravascular evaluation of atherosclerotic plaques; for example, by intravascular ultrasound (IVUS) or optical coherence tomography. 12 However, the type of atherosclerotic lesions that these rabbits develop are usually early fatty-streak-like plaques, containing high levels of macrophage-derived foam cells, but with no clear definition of a fibrous cap or necrotic core, and thus not resembling the complex vulnerable lesions found in humans. 13 This makes it very difficult to extrapolate the findings obtained in rabbits to the human condition. Low shear stress could provide the crucial pro-atherogenic trigger to initiate and promote the growth of an advanced lesion in the rabbit animal model, which in morphology and complexity would better represent advanced human lesions.
Here we aim to induce complex atherosclerotic lesion development in the New Zealand white rabbit by local shear stress reduction using our cast device in combination with a high cholesterol diet. This advanced plaque model in rabbits could provide a useful platform for preclinical validation of new intravascular diagnostic applications and interventional therapies for vulnerable plaque.
Materials and methods
Animal operations
All animal experiments were performed according to institutional and national regulations and approved by the Institutional Review Board on Animal Experiments of the Erasmus University Medical Center. We used 16 male New Zealand white rabbits (3–3.5 kg; Charles River, United Kingdom) divided into two groups of eight rabbits. In the first group, the effect of shear stress alteration on atherosclerotic plaque development was evaluated, while in the second group the effect of combined shear stress alteration and endothelial damage on atherosclerotic plaque development was studied. After arrival, the rabbits were individually housed and fed a normal chow diet (Arie Blok, The Netherlands) ad libitum with free access to water. After 2 weeks of acclimatization, the rabbits were fed a Western diet, containing 2% (w/w) cholesterol (Arie Blok). Two weeks after the start of the Western diet, a flow-altering cast (Figure 1) was surgically implanted around the right common carotid artery. Before surgery, animals were anesthetized with an intramuscular injection of ketamine hydrochloride (25 mg/kg, 100 mg/ml) and a subcutaneous injection of Domitor® (0.5 mg/kg, 1 mg/ml). The right common carotid artery was exposed and both halves of the cast were placed around the vessel and fixed with two sutures. In the second group, we inflicted endothelial damage by external manipulation and stretching of the carotid artery before cast placement. The wound was closed and the rabbits were allowed to recover. Rabbits were given 10 ml Ringer’s lactate, 10 ml 10% glucose solution and Temgesic® (0.05 mg/kg) subcutaneously for postoperative rehydration and analgesic. Twelve weeks after cast placement, rabbits were euthanized using an overdose of pentobarbital and the right common carotid artery was flushed and taken out, snap-frozen in liquid nitrogen and stored in −80°C until further analysis.
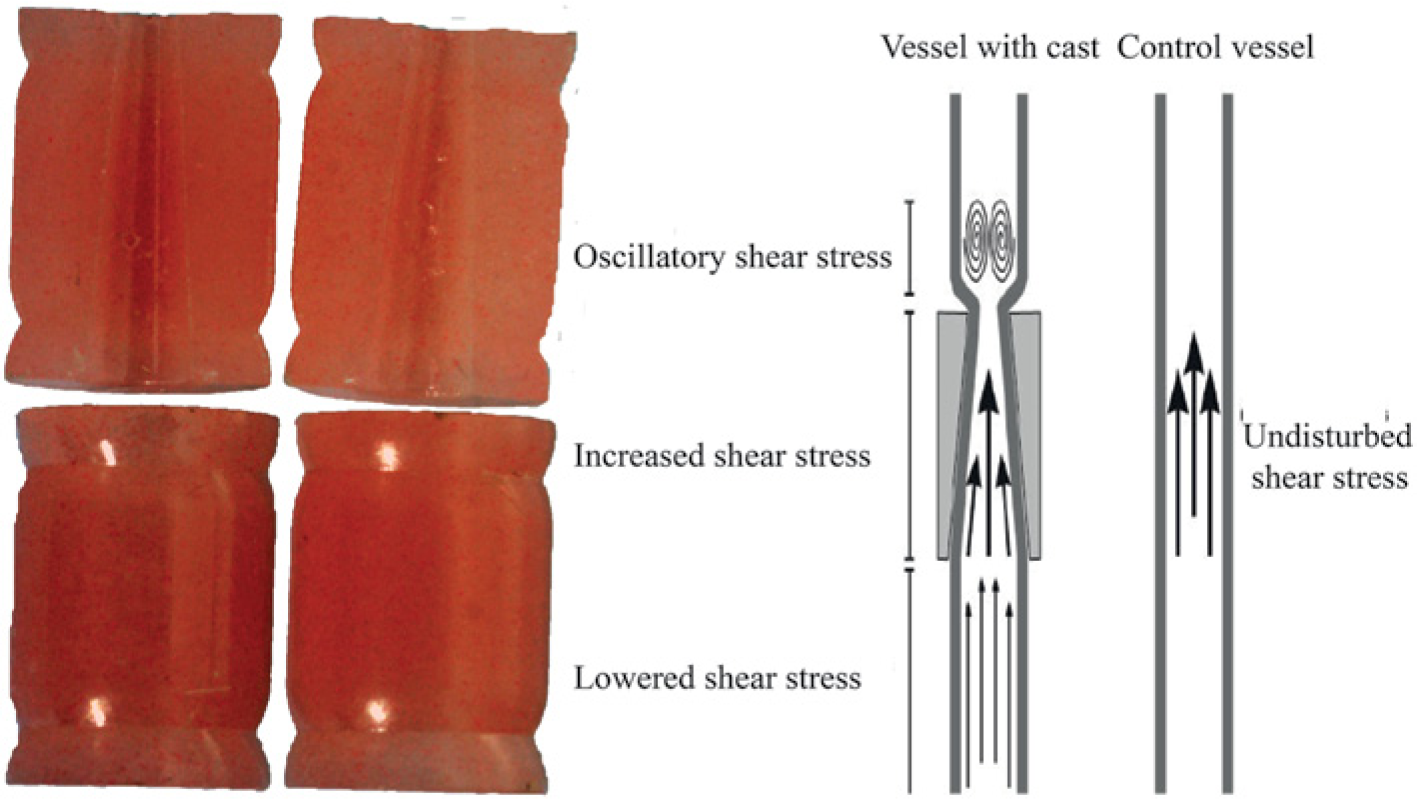
Example of cast and its schematic influence on shear stress.
Shear stress-altering cast device
To induce standardized alterations in shear stress, we up-scaled the existing murine cast to fit the rabbit carotid artery. Casts with different inner diameters were used, based on the initial diameter of the carotid artery as measured with ultrasound. The upstream inner diameter is 2.5, 2 or 1.5 mm (non-constrictive) and gradually declines to 1.25, 1 and 0.75 mm at the downstream side of the cast (constrictive), respectively, creating a 50% tapering. It has been shown that this tapering induces alterations in shear stress in the mouse with a region of low shear stress upstream of the cast, high shear stress in the cast and oscillatory shear stress downstream of the cast. 5
Doppler measurement
In order to validate that the rabbit cast could induce shear stress alterations, pulsed-wave Doppler velocities and the M-mode luminal diameter were measured in naive carotid arteries and after cast placement in 16 different New Zealand white rabbits. The ultrasound measurements were performed with the Vevo 2100 system (Visualsonics Inc., Canada) using a 40-MHz center frequency linear interfaced array transducer (MS550D). Briefly, the carotid artery was dissected and the pulsed-wave Doppler assessment was measured at a max angle of 65°. The luminal diameter was measured using M-mode (Figure 2). After cast placement, Doppler velocity measurements and M-mode were repeated upstream and downstream of the cast to determine whether cast placement induced shear stress alterations upstream and/or downstream of the cast. Flow through the artery was estimated using a Poiseuille profile and computed as:
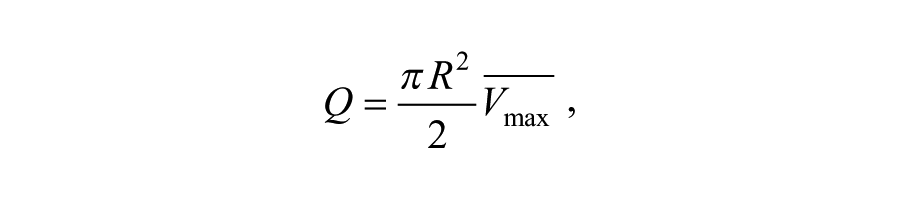
where Q is the flow, R is the time-average vessel diameter, and Vmax is the time-average peak velocity, measured in the center of the lumen. Consequently, shear stress was calculated as:
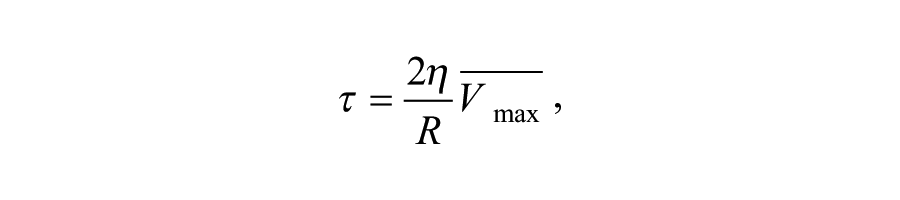
where τ is the wall shear stress and η is the blood viscosity, set as 3.10−3 Pa. Both Q and τ are determined prior to cast placement and are proximal to the cast after cast placement.
Cholesterol measurement
Blood samples were collected from the middle ear artery at baseline, cast placement and at sacrifice. Serum samples were stored at −80°C until further use. Total cholesterol, low-density lipoprotein (LDL), very low-density lipoprotein (VLDL) and high-density lipoprotein (HDL) were measured using Cobas® determination kits and measured using a Roche/Hitachi analyzer (Roche Diagnostics, Indianapolis, USA).
Histology and immunohistochemistry
The carotid artery region was serially sectioned in 6 µm cryosections. Histological and immunohistochemical analysis was performed on 72 µm intervals, covering the whole region upstream and downstream of the cast. In addition to routine hematoxylin-eosin (HE) staining, different immunohistochemical stainings were performed to evaluate plaque stability. Sections were stained for vascular smooth muscle cells (VSMCs) (anti-α-actin; DAKO, The Netherlands), macrophages (anti-RAM11 antibody; DAKO) and endothelial cells (anti-PECAM1 antibody; DAKO), followed by a biotinylated secondary antibody. We used 3,3’-diamino-benzidine as an enzyme substrate for horse radish peroxidase and the signal was visualized using bright field microscopy. Lipid deposition was analyzed using oil-red-O staining (Sigma, The Netherlands) and visualized by bright field microscopy.
Quantification and statistical analysis
Data analysis was performed using an automated commercial image analysis system (Impak C; Clemex Technologies, Canada) to evaluate differences in plaque phenotype upstream and downstream of the cast and between the two groups. HE staining was used to measure the intima/media ratio and percentage of luminal stenosis, α-actin staining to evaluate VSMC content, CD31 staining to evaluate neovascularization and RAM11 staining to evaluate macrophage content. Multiple groups were compared with one-way ANOVA and a subsequent Student-Newman-Keuls multiple comparisons test. Two-tailed Student’s t-tests were used to compare individual groups. Data are presented as mean ± SD; p-values < 0.05 were considered significantly different.
Results
Low shear stress and shear stress oscillations both induce plaque formation in cholesterol-fed New Zealand white rabbits
We first investigated the effect of the Western diet on the plasma lipid profile of the rabbits. The results are summarized in Table 1. After 2 weeks of Western diet, total plasma cholesterol, LDL-cholesterol and VLDL-cholesterol levels showed a significant increase that was maintained until the time of sacrifice at 12 weeks. There was also a significant increase in HDL-cholesterol after 2 and 12 weeks of Western diet.
Rabbit lipid profile.
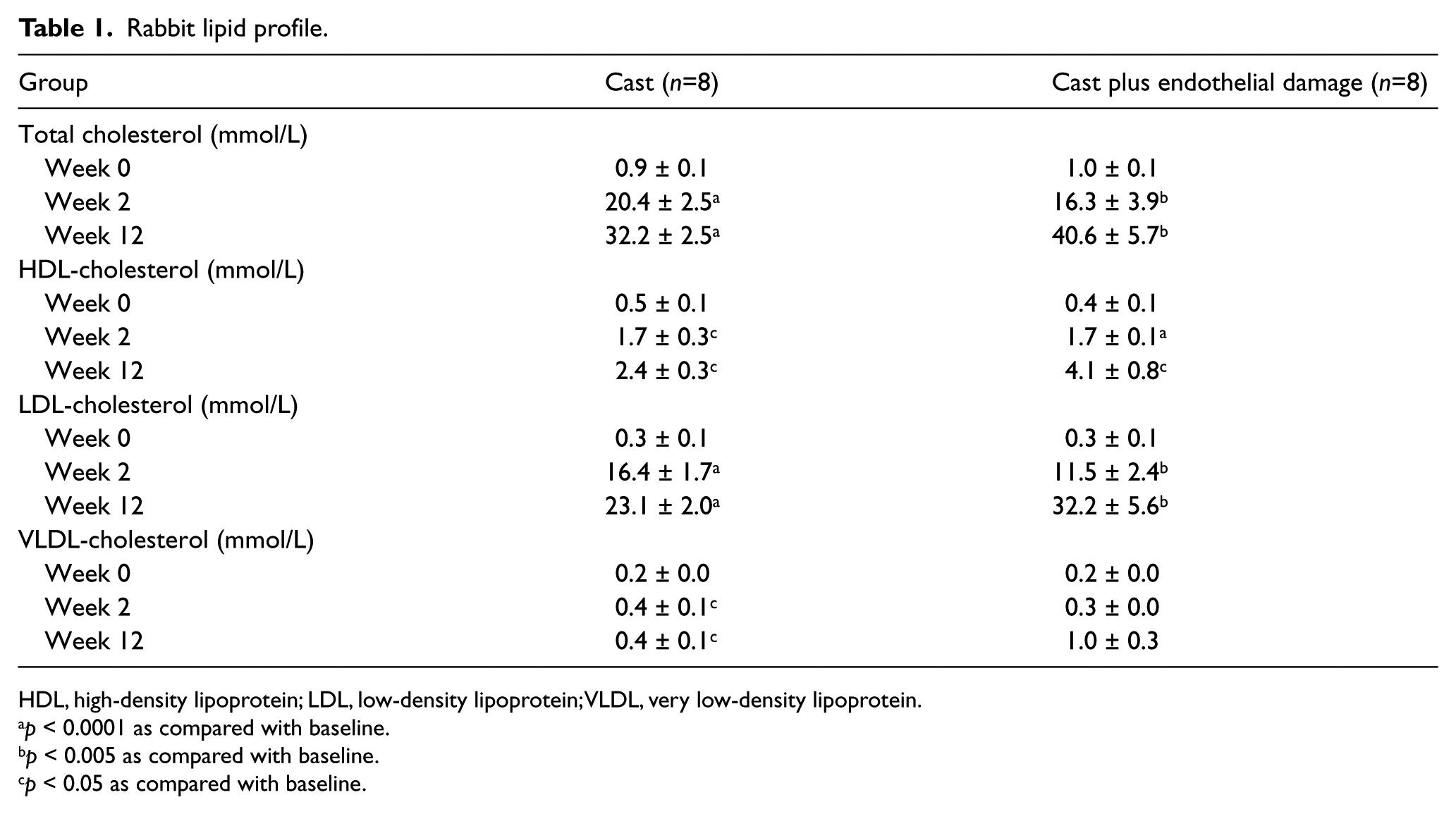
HDL, high-density lipoprotein; LDL, low-density lipoprotein; VLDL, very low-density lipoprotein.
p < 0.0001 as compared with baseline.
p < 0.005 as compared with baseline.
p < 0.05 as compared with baseline.
Doppler velocity measurements of blood flow assessed the effect of the perivascular cast on shear stress in the straight rabbit carotid artery. Before cast placement, the time-averaged peak blood velocity was 527 ± 215 mm/s. Doppler velocity measurements close to the vessel wall showed a symmetric profile, while the center velocity was 30–40% higher than at the vessel wall. As the velocity at the wall cannot be accurately measured, a parabolic profile is assumed for flow computation. The average (± SD) flow before cast placement was 29.4 ± 9.0 ml/min and the shear stress at the wall 4.3 ± 2.3 Pa. After cast placement, the downstream segment registered flow-reversal near the vessel wall consistent with the occurrence of oscillatory shear stress (Figure 2). Upstream of the cast, the time-average peak blood velocity at the center was significantly reduced to 320 ± 176 mm/s (p=0.02). The flow reduced to 19.7 ± 13.7 ml/min (p=0.02) and the shear stress reduced to 2.4 ± 1.2 Pa (p=0.03). The lumen radius before cast placement and proximal to the cast after cast placement remained unchanged (0.8 ± 0.1 mm for both cases). Combined, these data indicate that shear stress levels in the upstream region from the cast were significantly reduced, while oscillatory shear stress was successfully induced in the downstream region.
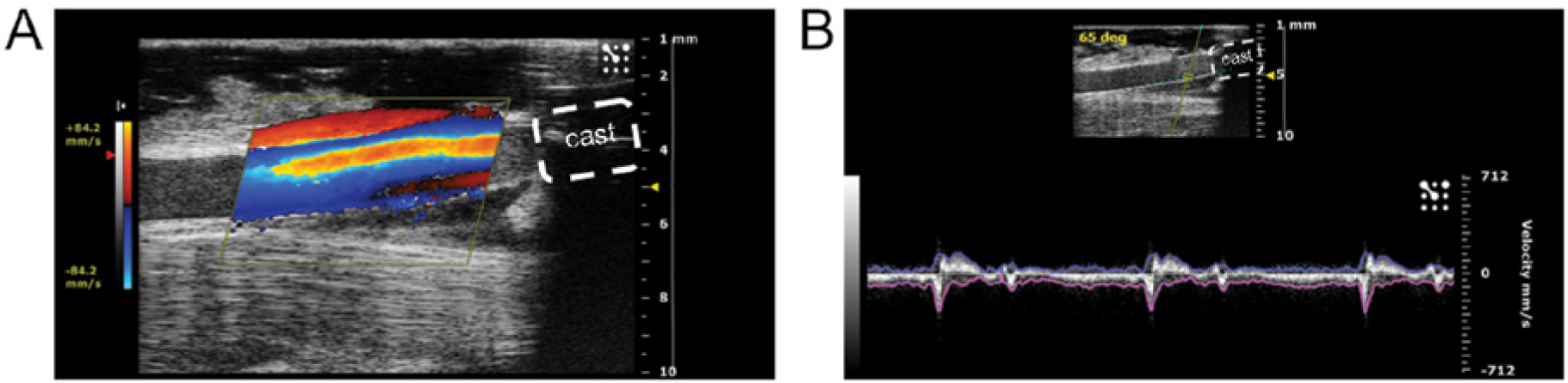
Doppler measurements of the rabbit carotid artery. (A) Color flow in the carotid artery after cast placement, downstream of the cast. (B) Pulse wave velocity measurement downstream of the cast, near the vessel wall, showing flow reversal.
To examine the effect of cast placement on atherosclerotic lesion size in hypercholesterolemic rabbits, we sacrificed the animals after they had been exposed to shear stress alteration for 12 weeks. The upstream and downstream carotid regions with low and oscillatory shear stress were assessed by histological evaluation for atherosclerotic lesion growth. Low and oscillatory shear stress both triggered atherogenesis in the upstream and downstream vessel regions in 50% (4/8) of the rabbits. In 12.5% (1/8) of the rabbits, atherosclerotic plaque developed only upstream of the cast, while in 25% (2/8) of the rabbits no significant lesions were detected (results summarized in Table 2). These results were independent of cholesterol levels. Figure 3 shows the intima/media ratio upstream and downstream and representative pictures of HE staining. The lesion size between the upstream and downstream regions was not significantly different (0.25 ± 0.09 and 0.24 ± 0.11, respectively; p>0.05).
Effect of cast placement on atherosclerotic plaque development.

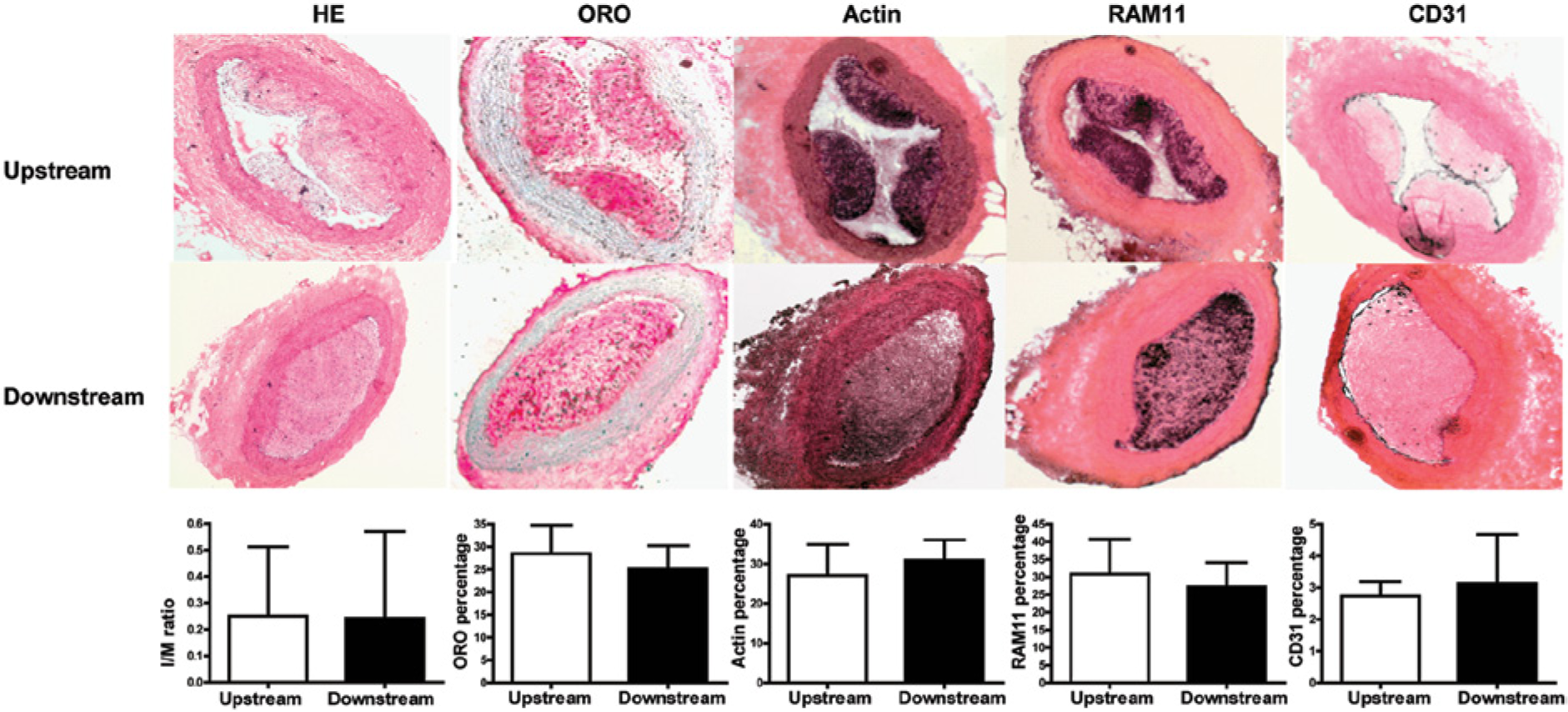
Analysis of plaque phenotype upstream and downstream of the cast in the cast only group. Representative pictures of hematoxylin/eosin staining (HE), lipid staining (oil-red-O; ORO), vascular smooth muscle cell (actin), macrophages (RAM11) and endothelial cells (CD31) upstream and downstream of the cast. Quantification of the different stainings revealed no significant difference between upstream and downstream of the cast. Graphs show mean per group ± SD, n=8 per group.
Shear stress alteration promotes atherogenic growth beyond the fatty streak phenotype
After the initial assessment of lesion size, we performed immunohistochemical analysis of the plaques for lipid, VSMC, macrophage and endothelial cell content in order to analyze plaque phenotype. No clear difference in lesion phenotype was observed between the plaques found in the low or the oscillatory shear stress region. As shown in Figure 2, both upstream and downstream atherosclerotic plaques contained abundantly foam cells as we observed a high percentage of lipids (23.0 ± 10.0% vs 25.2 ± 5.1%) and high macrophage content (30.8 ± 9.9% vs 27.3 ± 11.2%). This was accompanied by the accumulation of VSMCs (27.1 ± 7.7% vs 30.9 ± 5.1%) and CD31-positive endothelial cells (2.2 ± 0.8% vs 3.1 ± 1.0%) in the plaque, suggesting the presence of a fibrous cap and intimal neovascularization. However, no necrotic core could be detected and intraplaque hemorrhaging was absent in the lesions.
The combination of low shear stress and endothelial damage significantly increases the size of atherosclerotic plaque in the upstream region
Although shear stress alteration alone was capable to induce atherosclerotic lesions beyond the fatty streak state, as indicated by the presence of intimal VSMCs in the fibrous cap, these lesions still lacked a distinct necrotic core and showed a relative thick fibrous cap. We aimed to further increase lesion vulnerability by combining shear stress alteration with mechanical endothelial damage. Combined treatment increased the number of rabbits that developed atherosclerotic plaque both upstream and downstream of the cast to 75% (6/8). In the remaining two rabbits we found atherosclerotic plaque either upstream or downstream of the cast (the data are summarized in Table 2). Quantification of lesion size by intima/media measurement showed that there was a significant increase in the low shear stress, upstream region of the cast, as compared to the group that only received cast placement without endothelial damage (from 0.25 ± 0.09 to 0.66 ± 0.12, p < 0.05; Figure 4). Endothelial damage did not significantly affect the intima/media ratio in the downstream region (0.24 ± 0.11 vs 0.45 ±0.10, combined versus single treatment, respectively, p>0.05; Figure 4). Within the group that received combined endothelial damage and cast placement there was a trend towards a difference in lesion size in the upstream and downstream vascular segment (0.66 ± 0.12 vs 0.45 ± 0.10, p=0.07; Figure 4). These findings were confirmed when we measured lesion size by percentage of luminal stenosis (Figure 4).
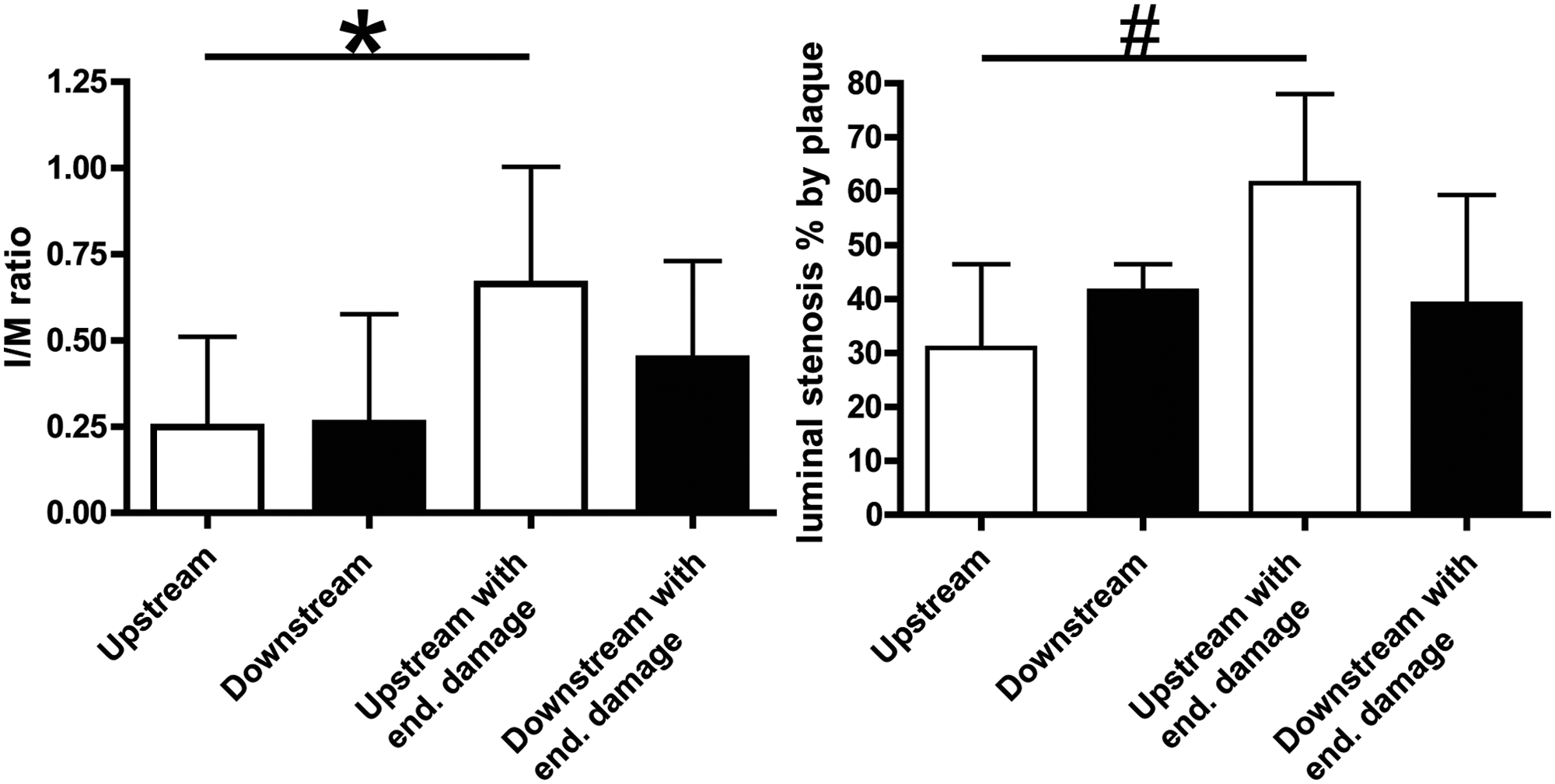
Analysis of plaque size by intima/media (I/M) ratio and percentage of luminal stenosis. Combination of cast placement and endothelial damage significantly increases plaque size upstream of the cast, as measured by I/M ratio and percentage of luminal stenosis. Graphs show mean per group ± SD, n=8 per group. *p < 0.05, 0.66 ± 0.34 vs 0.25 ± 0.26; #p < 0.05, 61.5 ± 16.6 vs 30.9 ± 15.6.
Combined treatment of shear stress alteration and endothelial damage increases plaque vulnerability
We compared lipid, macrophage, endothelial cell and VSMC content between the cast placement with endothelial damage and the cast placement without endothelial damage groups to see whether endothelial damage increased plaque vulnerability (Figure 5). There was a significant decrease in VSMCs upstream of the cast as demonstrated by VSMC-actin staining (from 27.1 ± 7.7% to 15.6 ± 4.8%, cast versus cast with endothelial damage, respectively, p < 0.05). Downstream of the cast there was also a decrease in VSMC content, but this reached no statistical significance (from 30.9 ± 5.1% to 21.1 ± 7.0 %, cast versus cast with endothelial damage, respectively, p>0.05).
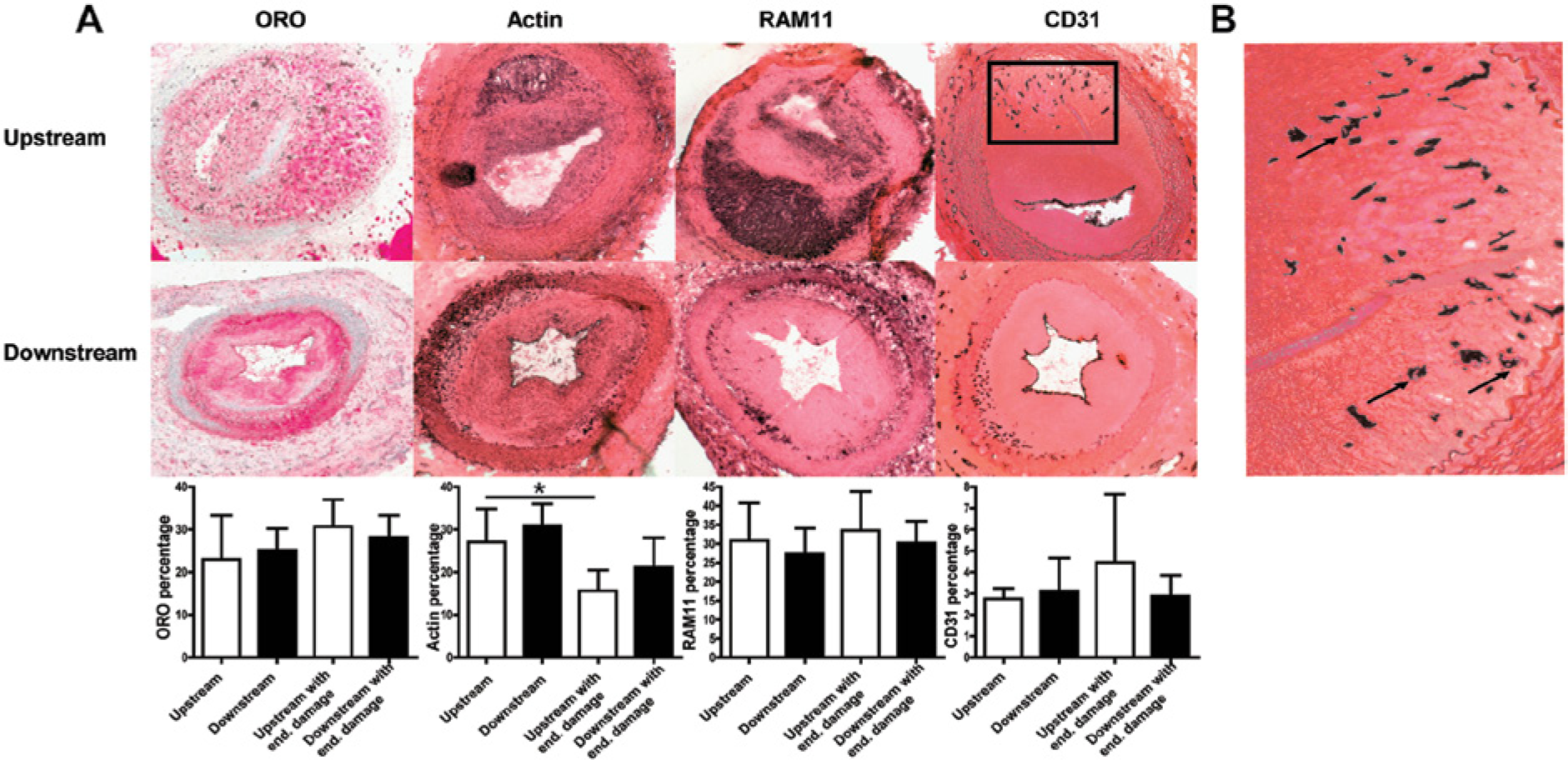
Analysis of plaque phenotype upstream and downstream of the cast in the combined cast and endothelial damage group. (A) Representative pictures of lipid staining (oil-red-O; ORO), vascular smooth muscle cell (actin), macrophages (RAM11) and endothelial cells (CD31) upstream and downstream of the cast of the combined cast and endothelial damage group. There was a significant decrease in vascular smooth muscle cell content upstream of the cast, compared with upstream of the cast only group. Graphs show mean per group ± SD, n=8 per group. *p < 0.05, 15.6 ± 4.8% vs 27.1 ± 7.7%. (B) Magnification of CD31 staining showing newly formed vessels with lumen formation (arrows) in the plaque.
These data indicate that endothelial damage aggravated plaque vulnerability by a decrease in plaque stabilizing VSMCs. In addition, we demonstrated the presence of CD31-positive endothelial cells in the intimal area, pointing towards neovascularization in the plaque (Figure 5).
Discussion
Many animal models used to study vulnerable plaque are long-term experiments and the plaques often do not fulfill the criteria for advanced human atherosclerotic plaque. The rabbit is widely used as it is easy to keep and handle, suitable for cardiovascular imaging, both non-invasively as well as intravascular, and is not as costly as the pig. Different models to develop atherosclerosis in rabbits exist and most models are based on a high cholesterol diet, with the percentage of cholesterol usually ranging from 0.5% to 4%. A high cholesterol diet in rabbits typically leads to early atherosclerotic lesions with foam cells.14 –16 This high cholesterol diet has been combined with a number of other methods causing endothelial dysfunction or injury, such as balloon denudation, hollow cuff implantation, hypertension, and the induction of diabetes or infection.17 –21 However, none of these methods have specifically created a human-like vulnerable plaque model in rabbits. Low shear stress is a known atherogenic stimulus and we have previously shown that induction of low shear stress with a flow-tapering cast induces vulnerable plaque formation in the carotid artery in mice. 5 Although Marano and co-workers showed that increased shear stress protected rabbits from collar-induced neo-intima hyperplasia, 22 the induction of low shear stress to induce atherosclerosis has never been described in rabbits. Therefore, the aim of the current study was to develop a low shear stress-based vulnerable plaque model in the rabbit that would allow advanced lesions to develop in a large animal model with resemblance to human vulnerable plaque. We hypothesized that we could induce vulnerable plaques by flow alteration in New Zealand white rabbits within 12 weeks after cast placement using a scaled-up model of the flow-altering murine cast device. Furthermore, we investigated whether endothelial damage could accelerate this process.
In the murine model, the tapered cast is known to induce low shear stress upstream and oscillatory shear stress downstream of the device. To validate that we could also induce shear stress alterations in the carotid artery of the rabbit, we measured Doppler blood flow velocities in the naive carotid artery and upstream and downstream of the cast after cast placement. In the mouse model, a reversal of flow downstream of the cast and a reduction of approximately 33% of flow (15 N/m2 to 10 N/m2) in the upstream region, is induced by cast placement. 8 Comparable with the murine model, we measured a reduction in flow of 33% upstream of the cast (30 ml/min to 20 ml/min), while downstream of the cast oscillatory shear stress was observed, indicating that we were able to recreate the same hemodynamic conditions as in our murine model.
In our rabbit model, we were able to create atherosclerotic plaques with a high amount of lipids, high macrophage and VSMC content, and intimal neovascularization as indicated by CD31-positive endothelial cells in the plaque. There were no signs of intraplaque hemorrhaging or plaque rupture. The combination of cast placement with endothelial damage increased lesion size and decreased VSMC content, but did not affect other vulnerable plaque (VP) parameters. VSMCs are crucial in determining the thickness and integrity of the fibrous cap as they constitute a major part of this fibrous cap and also produce the extracellular matrix which further stabilizes and thickens the cap. On the other hand, macrophages produce matrix metalloproteinases which break down extracellular matrix, thus weakening the fibrous cap. In our combined group, we also found a high amount of macrophages but fewer VSMCs, indeed indicating thinning of the fibrous cap. In addition, it is known that intimal neovascularization promotes plaque progression towards a vulnerable plaque, as those newly formed vessels are often leaky and form the basis of intraplaque hemorrhaging. 23 Recently, Hellings and co-workers also showed that intimal neovascularization not only promotes intraplaque hemorrhaging but also predicts clinical outcome, as patients with increased vessel density in a single excised carotid plaque were at higher risk for future cardiovascular events. 24 In both our cast and combined groups, neovascularization was also observed, indicating progression towards a more advanced lesion. Moreover, our model could be used to develop and validate new tools to study intraplaque neovascularization. As mentioned earlier, different rabbit models exist to create atherosclerotic lesions in rabbits. None of them have fulfilled all the criteria for human-like vulnerable plaques. One may postulate whether our model could be further improved by combining it with a different existing rabbit-atherosclerosis method. For example, it has become clear that the immune system plays an important role in atherosclerotic plaque formation and it has been postulated that activation of inflammatory cells could trigger atherosclerosis. Indeed, Lehr and co-workers showed that repeated intravenous endotoxin injection in hypercholesterolemic rabbits accelerated atherosclerosis formation. 20 Combination of our rabbit model with injection of endotoxin might enhance plaque vulnerability by stimulation of the immune system, thus stimulating necrotic core formation. Constantinides and co-workers showed that the rabbit can also be used to study plaque disruption and rupture, although the rupture described in their study was not spontaneous as it was induced with Russell’s viper venom and histamine. 25 Their original protocol was a long-term experiment (more than 1 year), so it would be interesting to see whether we could induce the same amount of plaque rupture in our shorter protocol.
Although we created the same conditions as in our murine vulnerable plaque model, hypercholesterolemia in combination with altered shear stress, the plaques in the rabbit were not as vulnerable as those observed in the murine model. We did find abundant intimal neovascularization and high macrophage and lipid content, but these advanced lesions lacked a prominent necrotic core or intraplaque hemorrhage. This lack of a vulnerable phenotype in the altered shear stress-induced lesions in the New Zealand white rabbit model could be explained by a number of arguments. Wild-type mice are normally protected against atherosclerosis, as their lipoprotein profile is enriched in HDL versus LDL, whereas in humans this profile is reversed. LDL oxidation and deposition in the vessel wall promotes atherogenesis. In the atherosclerosis-prone ApoE–/– mouse, a functional ApoE gene is lacking, causing a shift in the lipoprotein profile, mainly towards VLDL and in a lesser extent to LDL. Furthermore, the absence of ApoE causes a pro-inflammatory state due to T-lymphocyte activation and proliferation and promotes VSMC proliferation that is present from birth and favors atherosclerotic plaque development. 26 Similar to mice, the lipoprotein profile of wild-type rabbits is larger in the HDL fraction, thus rendering the species protected against atherosclerosis. We were able to induce a state of hypercholesterolemia via a high cholesterol diet. However, the main mass of plasma cholesterol in diet-induced hypercholesterolemia in rabbits is found in LDL, in contrast to VLDL in the ApoE–/– mice. The use of transgenic rabbits, such as the Wattanabe Heritable Hyperlipidemic rabbit (WHHL), which, like the ApoE–/– mice, are born with lipoprotein profiles with high LDL/HDL rations due to a defect in the LDL receptor, could provide a more suitable experimentation model for the induction of vulnerable plaque development by shear stress alteration. We are currently conducting a study in WHHL rabbits to test this hypothesis.
Our study has several limitations. First, the animals were already sacrificed at 12 weeks after cast placement. It is possible that we would be able to create a more human-like vulnerable plaque when the duration of the protocol was prolonged. The 12-week duration for the experiments was chosen because we wanted to create a rapid model for vulnerable plaque development. Moreover, the duration that New Zealand white rabbits can be fed a cholesterol-rich diet is limited as they develop hepatic failure when it is fed to them for a longer period. 27 Second, we did not perform the Doppler wave measurement in the same group of rabbits in which we induced the immunohistological analysis of the atherosclerotic plaques. As a result we cannot correlate the degree of shear stress change with morphological changes in the carotid artery (i.e. plaque formation and phenotype) per animal. Third, we decided to injure the endothelium by external stretch, which is an easy and quick procedure. However, this is not as controlled as balloon denudation and we do not know if any additional damage was done to the vessel. For example, we might have injured the vasa vasorum surrounding the carotid artery, leading to impaired oxygen supply to the vessel.
In summary, we present a novel method to induce atherosclerosis in New Zealand white rabbits using a flow-altering device. Both low and oscillatory shear stresses are capable of inducing atherosclerosis in a large animal model. The phenotype of the lesion induced by low shear stress is beyond that of the fatty streak, with infiltration of VSMCs into a thick cap and macrophages, abundant lipid deposition and neovascularization, but a prominent necrotic core is still lacking, rendering the overall phenotype of the lesions less vulnerable than those induced with the similar device in ApoE–/– mice.
Footnotes
Declaration of conflicting interest
The authors declare no conflicts of interest.
Funding
This work was funded by grants from the Dutch Organization for Scientific Research (grant no. 91776325 [HJD] and grant no. 91696061 [CC]).