
Abstract
Multiple sclerosis (MS) is a complex neurological disorder characterized by an aberrant immune system that affects patients’ quality of life. Several environmental factors have previously been proposed to associate with MS pathophysiology, including vitamin D deficiency, Epstein-Barr virus (EBV) infection, and cigarette smoking. These factors may influence cellular molecularity, interfering with cellular proliferation, differentiation, and apoptosis. This review argues that small noncoding RNA named microRNA (miRNA) influences these factors’ mode of action. Dysregulation in the miRNAs network may deeply impact cellular hemostasis, thereby possibly resulting in MS pathogenicity. This article represents a literature review and an author’s theory of how environmental factors may induce dysregulations in the miRNAs network, which could ultimately affect MS pathogenicity.
Keywords
Background
Multiple sclerosis (MS) is a multifactorial disease that is influenced by both genetic predisposing factors and the environment. Environmental factors, such as vitamin D deficiency, Epstein-Barr virus (EBV) infection, and cigarette smoking have been confirmed to be associated with MS; however, the mechanisms by which such variants influence disease pathophysiology are still unclear.1-21 Several theories have been previously proposed; herein, the author proposes a new theory linked to microRNA (miRNA) biogenesis. In this review, the author explores the influence of environmental variants on the miRNA network that could affect MS pathogenesis.
miRNAs, small noncoding RNA molecules with a size of approximately 22 nucleotides, are involved in posttranscriptional regulation of genes.22-25 miRNAs are thought to regulate 30% to 60% of genes, resulting in controlled cell proliferation, differentiation, and apoptosis.22-30 The functions of miRNAs are mediated through their binding to other macromolecules, leading to the formation of RNA-induced silencing complexes (RISCs). RISCs target complementary messenger RNA (mRNA) strands for the destruction and silencing of gene expression.22,23
miRNA genes are either located independently or within a coding region of other genes. Transcription of miRNA molecules is mediated by two RNA polymerases (II and III), forming long primary miRNA (pri-miRNA). The pri-miRNA, an approximately 100-nucleotide-long hairpin structure, is then acted upon by the Drosha enzyme and is transformed into one or more precursor miRNAs, called pre-miRNAs. The pre-miRNA is a 70-nucleotide precursor that is transported into the cytoplasm by exportin-5. Within the cytoplasm, the pre-miRNA is further processed into mature miRNA comprising 20 to 24 nucleotides by the enzyme, Dicer, which cleaves the double-stranded pre-miRNA into two strands, one of which is called a guide strand. The guide strand binds to an Argonaute protein, forming the RISC complex, while the other complementary strand is degraded (Figure 1).23,31 A mature miRNA can bind mRNA via a sequence specific to a target, thereby inhibiting its translation or even destroying it. Interestingly, a single miRNA can target several hundreds of mRNAs and have a profound effect on cellular functions.26,31

Process of miRNA transcription, transportation, and processing. Transcription of miRNA is mediated by two RNA polymerases (II and III), forming pri-miRNA. The pri-miRNA is then converted by Drosha into one or more pre-miRNA. The pre-miRNA is transported into the cytoplasm by exportin-5. Within the cytoplasm, pre-miRNA is further processed into mature miRNA by Dicer. Dicer then cleaves double-stranded pre-miRNA into two strands, the guide and the complementary strands. The guide strand binds Argonaute, forming the RNA-induced silencing complex, while the complementary strand is degraded. miRNA indicates microRNA.
Within the immune system, the miRNAs may affect cell activation, cytokine secretion, as well as immune tolerance. Dysregulation of miRNA expression has been observed in different immunological diseases, such as MS, type 1 diabetes (T1D), spontaneous systemic autoimmunity, systemic lupus erythematosus (SLE), and Alzheimer disease, and Parkinson disease.24,25,32 Intriguingly, in one report, significant differences in miRNAs expression were reported to be present in 364 peripheral blood mononuclear cell (PBMC) samples collected from MS patients undergoing remission and relapse compared with those from healthy controls. 33
Consequently, it would be intriguing to explain the reasons for such dysregulation of miRNA
The Theory
As described by Belbasis et al, vitamin D deficiency, EBV virus, and cigarette smoking are proven risk factors for MS. Their article is based on an extensive literature review of 44 meta-analyses studies conducted on 416 primary studies related to different MS risk factors. The results indicated a high risk of EBV and smoking on MS disease, while vitamin D deficiency posed a lower risk. 3
The risk posed by the environmental factors on MS pathogenicity needs to be addressed to explain the complexity of the disease. Many researchers have tried to address this issue, but a few missing pieces are still remaining to complete the whole picture. For example, vitamin D deficiency was proposed to increase the risk of MS, MS severity, and Expanded Disability Status Scale (EDSS) scores.34-41 By contrast, vitamin D supplementation could attenuate demyelination, inflammation, and promote remyelination of the central nervous system (CNS).42,43 Several studies have pointed out to the effect of vitamin D supplementation on the regulation of immune cells. 44 Vitamin D can modulate CD8+ T cell cytokine expression profile in the early part of the disease. 45 It also has a potent effect on CD4+ T cells46,47 and can improve regulatory T-cell (Treg) function and decrease accumulation of macrophages (MØ) in the CNS.48,49 Moreover, it can inhibit chemokine and cytokine expression, monocyte accumulation, and inducible nitric oxide synthaseaccumulation in the CNS.50,51 It can also suppress the expression and activity of Matrix metalloproteinases–2 and–9 (MMP-2 and MMP-9), 52 which are associated with a risk of MS. 53
Although many investigators have demonstrated the effect of vitamin D supplementation on MS or experimental autoimmune encephalomyelitis (EAE) models, the exact modulation pathways remain elusive. Several theories have been proposed to explain the modulation effects of vitamin D on MS pathophysiology: (1) Vitamin D supplementation could increase the vitamin D receptor (VDR)-binding sites across the genome, 25 which were found to be enriched in genes previously associated with MS or its variants.25,54,55 (2) Several single nucleotides polymorphisms (SNPs) in the genes CYP27B1, CYP24A1, and VDR were found to be associated with MS pathology; CYP27B1 and CYP24A1 are enzymes responsible for the catalysis of the 1,25(OH)2D.56-62 Some researchers even documented differential expression of the CYP27B1 gene in MS patients.63,64 (3) Vitamin D, additionally, ameliorates Th17 autoimmunity in EAE via recruitment of histone deacetylase 2 to the IL-17A promoter region (epigenetic modulation). 65
Nevertheless, the abovementioned modulation theories are not shown to be reproducible in all the studies. In a recent study, Pytel et al carried out whole-exome sequencing on the vitamin D signalling pathways in 15 families comprising at least two MS members. The study aimed to evaluate gene variants in the vitamin D pathway that may explain the presence of MS in these patients. Unfortunately, the researcher did not find any significant genetic variants that could explain the presence of the disease, suggesting that vitamin D may affect MS by some other means. 66 Agnello et al 67 also did not find a significant association between vitamin D-binding protein (VDBP) and CYP27B1 genetic variants and MS. In addition, Barizzone et al 68 did not find any genetic association between CYP27B1 and 2608 Italian and Belgian MS patients. Therefore, the risk of vitamin D deficiency needs to be further deliberated upon to explain its effects on MS.
EBV infection in MS is documented in several articles.69-77 Hassani et al have shown the presence of EBV in 90% of the tested MS cases (n = 101) compared with that in only 24% of the non-MS control cases (n = 24). EBV infection is associated with B cells, CD8+ T cells, astrocytes, and microglia.69,74-76
The increased incident of EBV infection in MS patients suggests that such patients may possess a genetic predisposition to be easily infected with EBV. Tørring et al transformed both MS and control lymphoblastoid cell lines with EBV virus and demonstrated that lymphocytes of the MS cell line had a significantly higher incidence of B-cell transforming events. This suggests that MS patients may be genetically more prone to EBV infection than controls. 78 In another study, the SNP (rs2516049) located in the human leukocyte antigen (HLA) region was associated with higher anti-EBV nuclear antigen-1 (anti-EBNA-1) titres in MS. Moreover, the authors found an association of other non-HLA genes with anti-EBNA-1 IgG titres in MS patients; the anti-EBNA-1 titres were positively correlated with the development of MS. 79
A ‘two-hit model’ may explain the infiltration of the immune cells into the CNS. The model proposes that a primary EBV infection occurring during early adulthood may trigger blood-brain barrier (BBB) permeability, allowing a few anti-EBV IgG producing cells to enter the CNS and are found in the intrathecal space. This may be triggered by a second event to attack. 80 The second event may be the presence of alpha-B crystallin, an amino acid protein in the CNS, which belongs to the family of small stress proteins found in the earliest stages of MS lesions. The infection of B cells with EBV results in the expression of the alpha-B crystallin that could be presented to cytotoxic T cells. 81 The second event could alternately be the presence of an antigen having a homology to EBV viral proteins. Such an antigen could be the myelin basic protein (MBP) peptide which is derived from the myelin sheaths surrounding an axon. Kumar et al have demonstrated molecular mimicry of MBP to viral EBNA-1 that could trigger autoreactivity of T cells to myelin sheaths. Hence, EBNA-1 is one of the most relevant non-self-antigens that is thought to induce MS. 82 Another possible antigen with amino acid homology to the EBNA-1 would be the chloride channel protein Anoctamin 2 (ANO2). ANO2 is a Ca2+-activated chloride channel that is important in neuronal excitability. ANO2 is expressed in neurons and glial cells from normal hippocampal and cortical regions. 83 Increased autoreactivity against ANO2 was documented in MS cases. Furthermore, the expression of ANO2 as small cellular aggregates near and within MS lesions was documented. 84 Besides, an increased antibody reactivity to the ANO2 was documented in MS (P = 3.5 × 10−36). The antibodies against EBNA-1 strongly competed for the ANO2 and vice versa, confirming the auto-reactivity against the ANO2 auto-antigen. 83 The second event could also be a human herpesvirus 6 variant A (HHV-6A) infection that could trigger EBV reactivation within the infiltrated B cells in the CNS. According to Flamand et al 85 and Cuomo et al, 86 the HHV-6A acts as an activator of EBV replication in the infected cell lines. HHV-6A was documented to increase MS neuroinflammation and relapses.87-90
When B cells were infected with the EBV virus, it upregulated antigen-presenting cell (APC)-related markers and activated the cross-presentation machinery. 91 EBV-infected memory B cells, more so, escaped the surveillance of T cells and maintained long-term latency by expressing lower levels of self-reactive and polyreactive antibodies. 92
First, it is important to understand the mechanism by which EBV affects cells and makes them more aggressive against CNS in MS patients. Therefore, the genetic makeup of MS patients must be considered. In a study by Zhou et al, 79 several HLA and non-HLA genetic markers were associated with higher anti-EBNA-1 titres in MS, which were positively correlated with the development of MS.
Second, the EBV encoded genes must be studied. Harley et al have revealed the function of the EBV-encoded EBNA2 protein in 44 genetic loci associated with MS in B cells. The EBV may exhibit its effect through DNA binding and downstream regulation of gene expression. 93 However, the mechanism by which EBV provoke immune cells to attack the CNS needs to be further investigated.
The risk proposed by cigarette smoking on MS severity has been documented in a number of articles.94-102 In MS, the risk induced by smoking may be influenced by the genes of an individual smoker. In a study by Hedström et al,14,17 smokers carrying HLA-DRB1*15 and lacking HLA-A*02 had a 13-fold increased risk for developing MS compared with nonsmokers without these genetic factors. Furthermore, carrying an A allele of the NAT1 SNP rs7388368A was associated with a risk to tobacco smoke in MS patients (odds ratio [OR] for the A/A = 5.17). The NAT1 encodes an enzyme involved in the metabolism of the smoke products. 103
One of the proposed mechanisms by which cigarette smoking induces MS is the dysregulation of immune cells. Nicotine, one of the components of cigarette smoke, can modulate bone marrow dendritic cells (DCs). DCs subjected to nicotine have reduced capacity for antigen uptake and produce substantially less Th1-promoting cytokine, IL-12. Nicotine is also responsible for proliferation and differentiation of ovalbumin (OVA)-specific naive T cells into Th2 cells. 104 Tobacco glycoprotein (TGP), a glycoprotein isolated from cured tobacco leaves, can induce B cell differentiation to produce immunoglobulin (Ig) without the stimulation of B cells. 105 Cigarette smoking can further increase the levels of MMP-9, facilitating infiltration of immune cells through the BBB. 106 Furthermore, smoking induces genome-wide dysregulation of DNA methylation. The DNA methylation dysregulation was even larger in MS associated with major risk factors, such as being a female and having an HLA-risk haplotype. Moreover, the DNA methylation correlated with the smoking load. 107 Although many researchers have tried to explain the risk of cigarette smoking on MS, the risk pertaining to a pathological load of cigarette smoke seems to be stronger than previous thought.
Considering that the theories on modulation of environmental factors could not explain the whole story behind MS-associated risk, a new theory must be investigated. In this context, miRNA is considered as an intriguing molecule to be explored. The miRNA dysregulation has been linked to EBV infection and cigarette smoking in several studies, while vitamin D supplementation was found to regulate the miRNA populations.108-122 MS has been associated with many dysregulated miRNAs with disease modulating effects.111,117,123-140 Finding the link between these two groups, environmental factors and miRNAs, may explain the risk of environmental factors on MS pathogenicity. However, only a few researchers have explored the environmental-miRNAs-MS axis.120,141-142 In one study, vitamin D supplementation ameliorated EAE by regulating the miRNA populations related to CD4+ T cell proliferation and differentiation. 120 In another study, significant expression levels of ebv-miR-BHRF1-2-5p and ebv-miR-BHRF1-3 in the circulation of relapsing-remitting multiple sclerosis (RRMS) patients were correlated with MS risk and increased EDSS scores. 141 Furthermore, the genetic makeup of miR-146a (rs2910164) correlated with baseline anti-EBNA-1 IgG titres, which helped in predicting the risk of MS. 142
In this review, the association between environmental factors, miRNA, and MS is further analysed. This could explain the missing piece in the environmental factor-MS picture. Towards this objective, several database searches were conducted to extract the dysregulated miRNA populations related to either environmental factors or MS/EAE. Next, the dysregulated miRNAs were cross-linked between the two groups to identify possible risk modulators. The possible miRNA modulators were then investigated for their function, targets, and possible risk related to MS. Therapeutic approaches towards MS were investigated for their roles in regulating the miRNA populations. In addition, the association of vitamin D supplementation, EBV, and cigarette smoke drugs with miRNAs regulation is deliberated.
Methods
A comprehensive literature search was conducted to extract data regarding environmental factor–induced dysregulations in miRNA networks and MS-associated miRNAs. The protocol in this article has not been registered previously. A flow diagram of the study selection methodology is presented in Figure 2.

Flow diagram of the study selection methodology.
Eligibility criteria
The following inclusion criteria were developed and refined as needed during the search process. For the inclusion criteria, all the studies were required to be (1) published journal articles or electronically published ahead of print articles, (2) peer-reviewed articles, and (3) written in English. No restrictions were placed on the dates of publication.
Search strategy and sources
A comprehensive list of search terms was developed by the author. Keyword searches included (1) multiple sclerosis or its variants (eg multiple sclerosis, MS; relapsing-remitting multiple sclerosis, RRMS; secondary progressive multiple sclerosis, SPMS; primary progressive multiple sclerosis, PPMS; experimental autoimmune encephalomyelitis, EAE) combined with microRNA or its variant terms (eg, multiple sclerosis miRNA, multiple sclerosis microRNA), (2) any term that reflected one of the three environmental factors included in this review (eg, vitamin D; EBV; cigarette smoking) combined with microRNA or its variant terms (eg, vitamin D microRNA), (3) any term that defines multiple sclerosis or its variants combined with one of the three environmental factors and microRNA or its variant terms, (eg, vitamin D microRNA multiple sclerosis), (4) any term that reflected multiple sclerosis or its variants combined with the word therapy or its variant terms (eg, therapy, drug, medicine) and microRNA or its variant terms (eg, multiple sclerosis drug microRNA), and (5) any term that reflected one of the three environmental factors combined with the word therapy or its variants and microRNA or its variant terms (eg, EBV drug microRNA).
Searches were conducted in the following databases: NCBI PubMed, Google Scholar, The Cochrane Library, and WHO Global Index Medicus. The PubMed search was filtered to include only studies with (1) English language and (2) full-text articles. As Google Scholar search yields all kinds of articles (peer-reviewed vs non-peer-reviewed articles), the search was first modified to yield only peer-reviewed articles. Later, the articles were searched for their availability through the unmodified Google Scholar engine. The Google Scholar search was modified through the Setting, Library Links, to include only studies found in Lane Community College, University of Oregano, University of Otago, University of Oulu, and University of Oxford libraries. Furthermore, the search was filtered to include only articles with (1) English language (2) that contain the exact search term in the title of the article (Advanced Search). The Cochrane Library search was limited to search terms found in title/abstract keywords. The WHO Global Index Medicus search was filtered to include only studies with (1) English language, (2) full-text article, that contain the (3) search terms in the title, abstract, and subject. Additional references were identified via the reference list from the searched articles. All searches were performed before October 2019.
Study selection
Unique titles and abstracts were independently screened for eligibility. Ineligible studies were excluded while the remaining studies underwent full-text review. The studies were screened for miRNAs that were dysregulated in multiple sclerosis or its variants (RRMS, SPMS, PPMS, paediatric MS, EAE) and were previously associated with one or more of the environmental factors (vitamin D, EBV, cigarette smoking). Studies that showed miRNA cross-link between two groups were further analysed to understand the significance of the association. Studies that did not match the purpose of this article were excluded. Study selection was carried out between August 2018 and October 2019.
Data extraction and summary measures
Data extracted included the following criteria: (1) miRNAs associated with vitamin D, EBV infection, or cigarette smoking, their P values and fold-changes; (2) whether the same miRNAs were associated with MS, MS subtype, or EAE, their P values and fold-changes; (3) the function of these associated miRNAs; (4) the targets of the associated miRNAs and the association of these targets with MS or its variants; (5) the miRNAs validated across different studies that are dysregulated in response to one or more environmental factors, MS, or its variants; (6) the possible outcomes of current therapeutic approaches to MS on these dysregulated miRNAs (supported by significant P values and fold-changes); (7) The possible outcomes of therapeutic approaches to environmental factors on miRNA populations and possibly MS or its variants (supported by significant P values and fold-changes).
Result and Discussion
Study selection
Database searches retrieved a total of 613 481 articles (Supplementary Table 1). The term ‘Multiple Sclerosis’ yielded 237 955 articles. The search was narrowed down to miRNA associated with MS; as such, the phrase ‘Multiple Sclerosis microRNA’ was used in the search engines, which yielded 454 articles. Keywords such as ‘RRMS miRNA’, ‘SPMS miRNA’, ‘PPMS miRNA’, ‘Paediatric Multiple Sclerosis miRNA’, and ‘EAE miRNA’ resulted in 454, 20, 16, 20, and 105 articles, respectively. The term ‘MS’ was excluded from the search terms due to the nonspecific search yields (Figure 2).
The terms ‘Vitamin D’, ‘EBV’, and ‘Cigarette smoking’ yielded 197 525, 66 720, and 38 098 articles, respectively. The search was further narrowed down to miRNA associated with vitamin D, EBV, and cigarette smoking which returned 203, 507, and 129 articles, respectively (Figure 2).
Furthermore, the phrases ‘vitamin D miRNA multiple sclerosis’, ‘EBV miRNA multiple sclerosis’, and ‘cigarette smoking miRNA multiple sclerosis’ were used, and the search returned 8, 9, and 0 articles, respectively. From the yield of ‘vitamin D miRNA multiple sclerosis’ search, 6 articles were excluded—2 articles were related to other none autoimmune nor neurological diseases, 3 non-miRNA informative articles, and 1 review article. Of the ‘EBV miRNA multiple sclerosis’ search yield, 5 articles were excluded; 1 article was related to other none autoimmune nor neurological diseases, 1 non-miRNA informative article, and 3 review articles. Different terms and phrases were used to extract as much information as possible (see Supplementary Table 1). Moreover, 37 articles were included using the reference lists of some of the articles; 8 articles were related to other autoimmune diseases, and 29 articles related to MS-associated genes (Figure 2). The purpose of this search was to define the dysregulated miRNAs related to MS or its variants and dysregulated miRNAs related to one or more of the environmental factors. The articles were further cross-linked to define the miRNAs which seemed to be dysregulated in both the groups.
Going through articles titles, abstracts, and contents, several miRNAs were cross-linked between MS or its variants and one or more of the environmental factors. Three articles could be linked between the search yields of the ‘vitamin D miRNA multiple sclerosis’ and ‘multiple sclerosis miRNA’ phrases. Also, 2 articles could be linked between the ‘vitamin D miRNA multiple sclerosis’ phrase and each of the ‘EAE miRNA’, ‘PPMS miRNA’, and ‘SPMS miRNA’ phrases. Only 1 was linked between the ‘vitamin D miRNA multiple sclerosis’ and the ‘RRMS miRNA’ phrases. The author also linked articles between the ‘vitamin D miRNA’ phrase and ‘multiple sclerosis miRNA’, ‘RRMS miRNA’, ‘EAE miRNA’, and ‘SPMS miRNA’ search phrases yielding 4, 4, 3, and 2 articles, respectively. The search yields with the phrase ‘EBV miRNA’ returned 15, 13, and 11 articles when cross-linked with the search yields of the phrases ‘multiple sclerosis miRNA’, ‘RRMS miRNA’, and ‘EAE miRNA’, respectively. The search result for the phrase ‘cigarette smoking miRNA’ returned 25, 11, 5, 3, and 2 articles when cross-linked with the search results of the phrases, ‘multiple sclerosis miRNA’, ‘EAE miRNA’, ‘RRMS miRNA’, ‘SPMS miRNA’, and ‘Paediatric MS’, respectively (Figure 2). The remaining articles did not match the purpose of this search.
For the search conducted on therapeutics, search terms including MS or its variants combined with therapy or its variant terms and miRNA or its variant terms were used (e, g., multiple sclerosis drug microRNA). The purpose of this section is to illustrate the effect of the ‘available therapeutics to MS approach’ on the dysregulated miRNA populations. The search yielded 141 articles, of which 64 articles were included in this review. The remaining articles did not match the purpose of the search. Also, phrases representing the environmental factors combined with therapy or its variant terms and miRNA or its variant terms were used (eg, EBV drug microRNA). The purpose of this section is to illustrate the effect of vitamin D supplementation, and EBV, or cigarette smoking drugs on the dysregulated miRNA populations. This was then cross-linked with the dysregulated miRNA populations found in MS or its variants. This search yielded 131, 87, and 45 articles for ‘vitamin D supplementation miRNA’, ‘EBV drug miRNA’, and ‘cigarette smoking drug miRNA’, respectively, of which 16, 12, and 2 articles were cross-linked with previous articles on ‘multiple sclerosis miRNA’ (Figure 2). The remaining articles did not match the purpose of the search.
In the end, the reference list included 272 articles that were eligible for this review. Of these articles, 25 articles were related to miRNA regulation in animals, while 90 articles studied miRNA regulation in humans. The selected studies were published between 1965 and 2019, with the majority being published between 2010 and 2019.
Data extraction
The data extracted were thoroughly reviewed under ‘vitamin D and miRNA’, ‘EBV and miRNA’, ‘cigarette smoking and miRNA’, ‘MS therapy and miRNA’, and ‘environmental risk factor modifying drugs and miRNA’ sections. The P values and the fold-changes for the associated miRNAs with MS or its variants or environmental factors are listed in Tables 1 and 2. The targets of the associated miRNAs are listed in Table 3. Please note that only the MS-associated genes are listed as targets in Table 3, as the list of the overall miRNA targets is quite long. Influence of environmental factors on miRNAs and the possible link to MS or EAE pathogenicity according to the fold-change of expression is further highlighted in Table 4. miRNAs that respond to MS or environmental factors therapies are mentioned in Table 5. The validated miRNAs across different studies found to be dysregulated in either MS or its variants or one or more environmental factors are noted in Table 6. Only data with significant P values (<.05) are listed in this review.
Dysregulation of miRNAs in response to vitamin D supplementation, EBV infection, and cigarette smoking.

Abbreviations: miRNA, microRNA; Case N, number of patient / EAE experimental mice included in the study; Control N, number of healthy controls included in the study; FC, fold-change; EBV, Epstein-Barr virus; NA, not available or not applicable; NPC, nasopharyngeal carcinoma; SAE, small air-way epithelium; mDCs, lung myeloid dendritic cells.
miRNAs previously associated with MS, MS subtypes, Paediatric MS, or EAE with respect to the P value, fold-change, sample type, and number of cases.
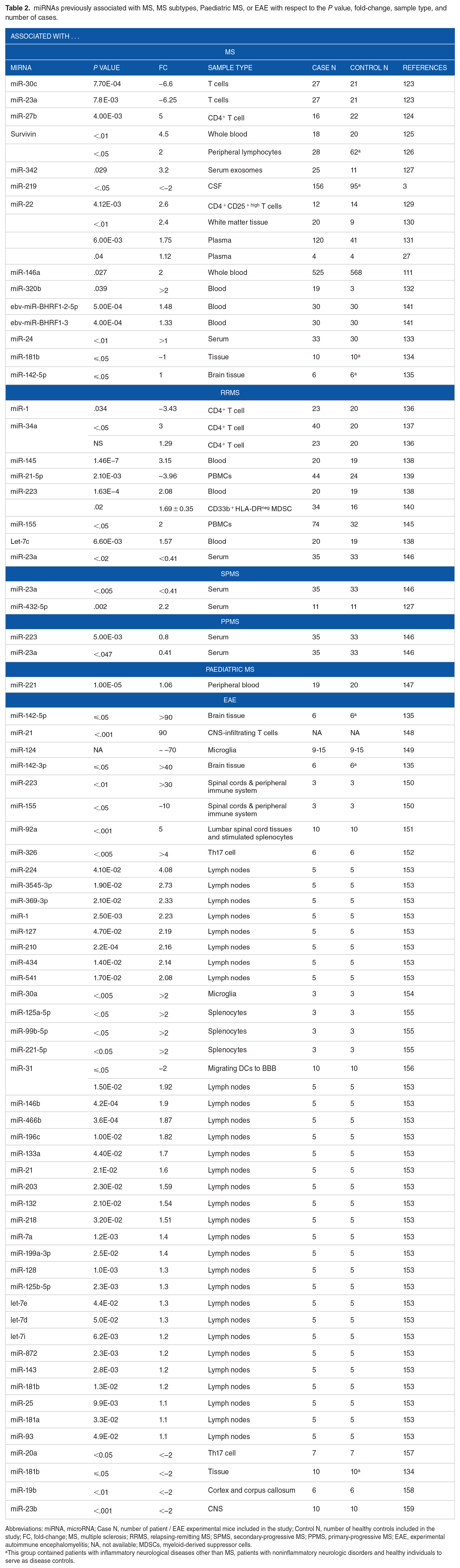
Abbreviations: miRNA, microRNA; Case N, number of patient / EAE experimental mice included in the study; Control N, number of healthy controls included in the study; FC, fold-change; MS, multiple sclerosis; RRMS, relapsing-remitting MS; SPMS, secondary-progressive MS; PPMS, primary-progressive MS; EAE, experimental autoimmune encephalomyelitis; NA, not available; MDSCs, myeloid-derived suppressor cells.
This group contained patients with inflammatory neurological diseases other than MS, patients with noninflammatory neurologic disorders and healthy individuals to serve as disease controls.

These genes have previously been associated with MS. The function of these genes is also illustrated in this table. The miRBase was used to identify the different miRNA targets and NCBI gene was used to define the function of the target genes.
Abbreviations: MS, multiple sclerosis; miRNA = microRNA.
miRNA populations which associated with MS or EAE and one or more of the environmental factors.
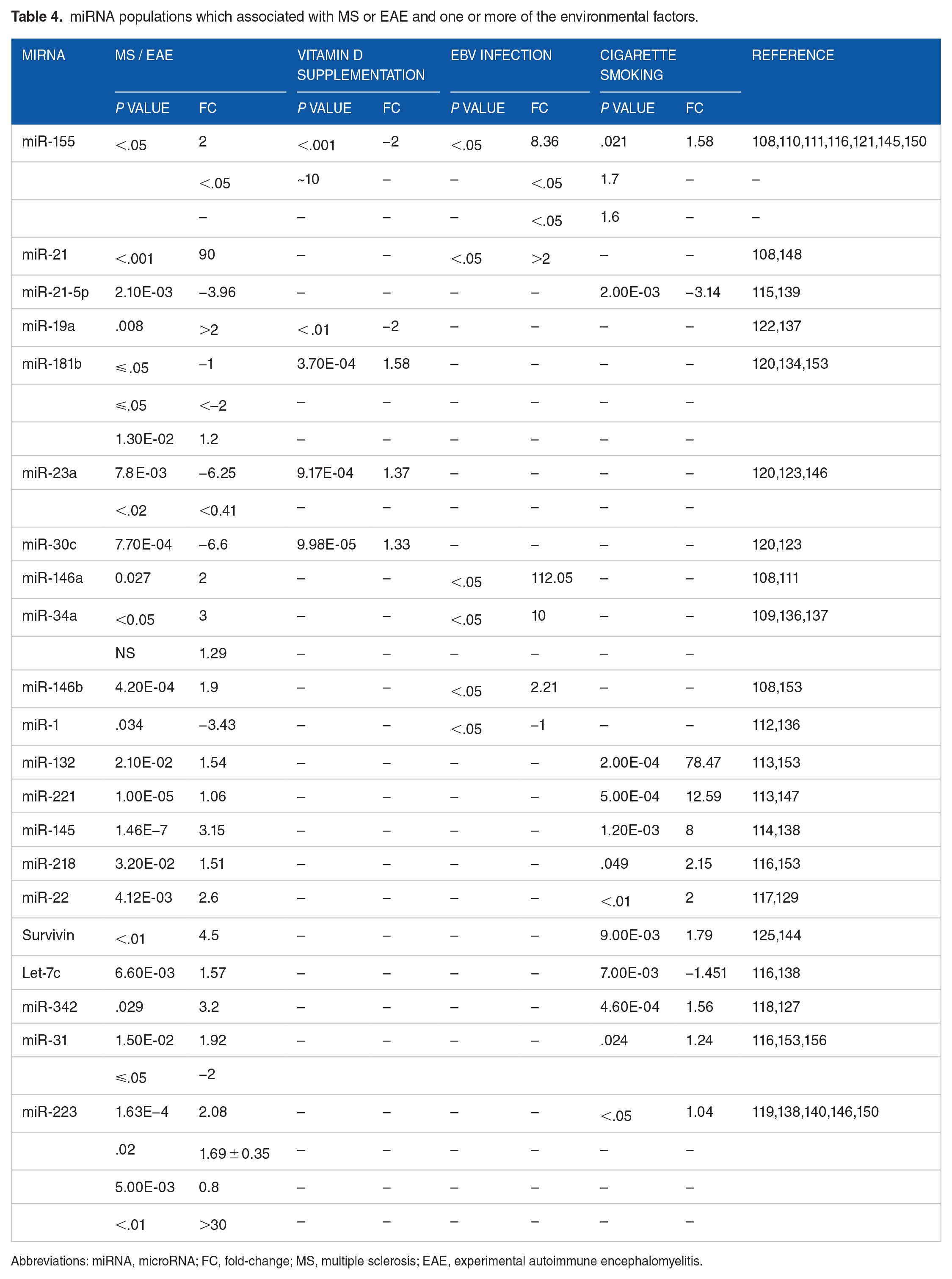
Abbreviations: miRNA, microRNA; FC, fold-change; MS, multiple sclerosis; EAE, experimental autoimmune encephalomyelitis.
Effect of MS therapy on the dysregulated miRNAs, and the effect of Vitamin D diet, EBV treatment, and smoking drugs on the dysregulated miRNAs.

Abbreviations: miRNA, microRNA; Case N, number of patient / EAE experimental mice included in the study; Control N, number of healthy controls included in the study; FC, fold-change; IFN-β, interferon-beta; DMF, dimethyl fumarate; AHSCT, autologous hematopoietic stem cell transplantation; NA, not applicable; PBMCs, peripheral blood mononuclear cells; DCs, dendritic cells; ACV, acyclovir; HNG, human neuronal-glial cells; PrEcells, primary prostatic epithelial cells.
miRNAs validated across different studies to be deregulated in response to either environmental stimuli (vitamin D, EBV, and cigarette smoking), or MS, MS subtypes, and EAE.

Abbreviations: MS, multiple sclerosis; EAE, experimental autoimmune encephalomyelitis.
miRNAs listed in this review represent those that are cross-linked between MS or its variants and one or more environmental factors. Those that were not cross-linked between the groups were excluded. Articles written in languages other than English, abstracts and non-peer-reviewed articles, and duplicate articles were also excluded from this study.
Vitamin D and miRNA
Vitamin D deficiency is an established risk factor for MS pathogenesis. 23 The relation between vitamin D and miRNAs regulation may shed light on the effect of vitamin D on MS patients. In fact, Zeitelhofer et al identified the effect of vitamin D supplementation on expression levels of 92 miRNAs in CD4+ T cells in EAE models. Of the 92 miRNAs, the most highly expressed 30 miRNAs were further analysed according to the pathways they regulate, including pathways associated with mitochondrial functions, protein synthesis, telomere extension, T-cell activation and differentiation. Interestingly, pathways important to cell survival were not affected by miRNA upregulation. The affected pathways were those pertinent to T-cell receptor (TCR) and IL-2 signalling. Upregulated miRNAs in CD4+ T cells included miR-30c, miR-134, miR-181b, miR-483, miR-9-3p, miR-449c, miR-23a, and miR-377. These are known to target several critical genes in T-cell activation and signalling (Table 1). 120
Indeed, two previous studies identified a significant downregulation of miR-30c and miR-23a in T cells of MS patients (Table 2).123,194 miR-30 overexpression was previously documented to reduce angiopoietin-2 (Ang2), which resulted in decreased levels of inflammatory cell-cell adhesion molecules, E-selectin, ICAM-1, and VCAM-1. 195 ICAM-1 and VCAM-1, known for their association with increased risk of MS, can mediate transfer of immune cells through the BBB. 196 Moreover, miR-30c is predicted to target TRAF3, one of several genes associated with an increased risk of MS (Table 3).160,162 This gene mediates signal transduction through members of the TNF receptor (TNFR) superfamily. The TRAF3 gene was found to be associated with MS risk in one of the largest studies, comprising 14 498 patients and 24 091 healthy controls, which was conducted through the International Multiple Sclerosis Genetics Consortium (IMSGC) (Table 3).161,162 The IMSGC was formed in 2003 to define MS susceptibility genes from different genome-wide scan (GWS) analysis studies. 197
The miR-23a was previously documented to be significantly decreased in the serum of relapsing-remitting MS (RRMS) and primary progressive MS (PPMS) patients and was observed to correlate with the EDSS score (Table 2).25,146 miR-23a has a suppressive effect on cytotoxic CD8+ T cells, and its overexpression is linked to the inhibition of LAMP1/CD107a surface expression on activated CD8+ T cells. 198 LAMP1/CD107a was shown to mediate the cytotoxic activity of CD8+ T cell in an antigen-specific manner. 199 Cytotoxic CD8+ T cells have been previously correlated with MS plaques. 200 miR-23a has also been predicted to target TNFSF14, another gene associated with an increased risk of MS, as reported by the IMSGC (Table 3).160,163 The product of this gene functions as a costimulatory factor for the activation of lymphoid cells and as a deterrent to infection by herpesvirus (Table 3). 161
Evidence of downregulated miR-181b was previously found in the lesions of white matter samples taken from autopsies of MS patients and in the lumbar cord tissue samples taken from EAE mice models. miR-181b was significantly reduced in T cells following immune stimulation. miR-181b acts as a negative regulator of MØ activation and differentiation into a proinflammatory phenotype. It also suppresses Th1 polarization. Overexpression of miR-181b was observed to be inversely correlated with Smad7 transcripts of EAE models (Table 2).134,153 Smad7 is a negative regulator of TGF-β, 24 and TGF-β is a promoter of Treg differentiation. 24 miR-181b additionally regulates the expression of GALC and MAPK1, both of which were reported by IMSGC to be significantly associated with MS (Table 3).160,163
In addition, miR-9-3p was previously linked to CNS development. Sim et al presented a very comprehensive article on the effect of miR-9-3p expression in the brain-hippocampus. In their article, they detail a series of experiments carried out using miR-9-3p sponge that targets the expression of miR-9-3p. According to their observations, miR-9-3p is critical for hippocampal long-term potentiation (LTP) and long-term memory. Likewise, miR-9-3p was linked to the regulation of LTP-related genes, such as Dmd and SAP97. 201 Hence, the upregulation of this miRNA by vitamin D could be beneficial to neuronal recovery.
Although miR-449c has not yet been linked to MS, it can target several genes, including CXCL-10, CXCL-11, NFKBIA, SERPINE1, IL2RB, CCR1, MYC, SNAI1, and BCL6, which are involved in the functioning of the immune system. The downregulation of this miRNA was associated with an increase in the expression of the aforementioned targets and resulted in immune cell trafficking into the brain, production of proinflammatory mediators, and neuronal apoptosis. 202 C-X-C motif chemokine ligand (CXCL)-10 was associated with the accumulation of T cells in the CNS of MS patients. It was also reported to be highly expressed in CNS lesions, cerebrospinal fluid (CSF), and serum of MS patients in several studies.203-207 Although not many articles were uncovered on CXCL-11, in 1 article, CXCL-11 was not observed to be associated with MS. 207 NFKBIA has previously been associated with primary progressive MS (P value < .01). 208 A genotype in the promoter of the SERPINE1 was associated with MS in women (genotype: 4G4G; OR = 0.376; P = .032). 209 Furthermore, IL2RB was found to be associated with MS in 116 French sib-pairs after stratification of the analysis based on sharing (or not) of the HLA-DRB1*1501 allele (P = 3 × 10−9; maximum likelihood score [MLS] = 2.32) 210 ; however, IL2RB was not associated with another larger MS cohort from Madrid (Spain) (P = .30, OR = 0.9). 211 A study by Rottman et al 212 indicated that CCR1-/- mice were partially protected against EAE. Mice with CCR-/- were less prone to develop EAE and had less severe disease (55% incidence, P < .001; severity 1.2 ± 0.2, P < .001). The Eltayeb group has indicated the presence of a large number of CCR1 +/CCR5 + cells in the inflammatory brain lesions of EAE rats and has studied the role of CCR1 selective antagonist in a therapeutic plan. 213 The Misu group found increased levels of CCR1 T cells within the CSF of MS patients (P < .01). 214 The Trebst group have pointed out that CCR1 +/CCR5 + monocytes represent approximately 70% of the CSF monocytes regardless of the presence of CNS pathology. Moreover, Trebst et al 215 also found that monocytes infiltrating CNS lesions downregulate CCR1 expression. Nevertheless, the role of CCR1 in the infiltration of immune cells of the CNS has been revealed in several other articles.216-218
Vitamin D can also attune MØ-induced inflammation via targeting miR-155. miR-155 targets the suppressor of cytokine signalling -1 (SOSC1), a negative regulator of cytokine signalling (Table 1). 121 It can also induce T-cell differentiation in the Th17 and Th1 cell lines. 219 miR-155 targets transcription factor Ets1, a negative modulator of Th17 differentiation. 24 In addition, it targets CD47, a molecule involved in self-recognition and protection of cells from phagocytosis. 26 miR-155 also suppresses src homology 2 domain-containing inositol-5-phosphatase 1 (SHIP1), a known negative inhibitor of proinflammatory pathways of MØ and DCs. 24 Besides, miR-155 has been shown to suppress essential neurosteroids in the white matter of MS cases.26,220 In addition, miR-155 was upregulated in RRMS cases and more importantly in the white matter active lesions (Table 2).145,221 Notably, this miRNA targets some of the genes associated with MS risk (Table 3).160-163,166
Vitamin D has also been linked to miR-19a suppression and escalation of the anti-inflammatory IL-10 cytokine in B cells, along with ameliorating allergic rhinitis (AR) symptoms (Table 1). 122 miR-19a is a member of the miR-17-92 cluster, which includes six miRNAs: miR-17, miR-18a, miR-19a, miR-20a, miR-19b-1, and miR-92a. Augmentation of this cluster has been associated with several immunological diseases, such as lymphoproliferative disease, autoimmunity, cancer, allergic asthma, and premature death, as well as EAE pathogenesis (Table 2).24,122 miR-17-92 can suppress tumour suppressor phosphatase and tensin homology (PTEN), proapoptotic protein Bim and Ikaros Family Zinc Finger 4 (IKZF4), and promote Th17 differentiation in EAE. Specifically, miR-19 can dampen PTEN, SOCS1, deubiquitinase A20, and enhance Th2 responses.24,222 Evidence of miR-19a upregulation during the relapse phase of RRMS has been documented before, further supporting the role of this miRNA in immune dysregulation. 137
Although few studies seem to support the hypothesis related to the effect of vitamin D on miRNAs expression levels associated with MS pathogenicity (Figure 3A), the data presented in this review may encourage researchers to seek better understanding through methodological studies.
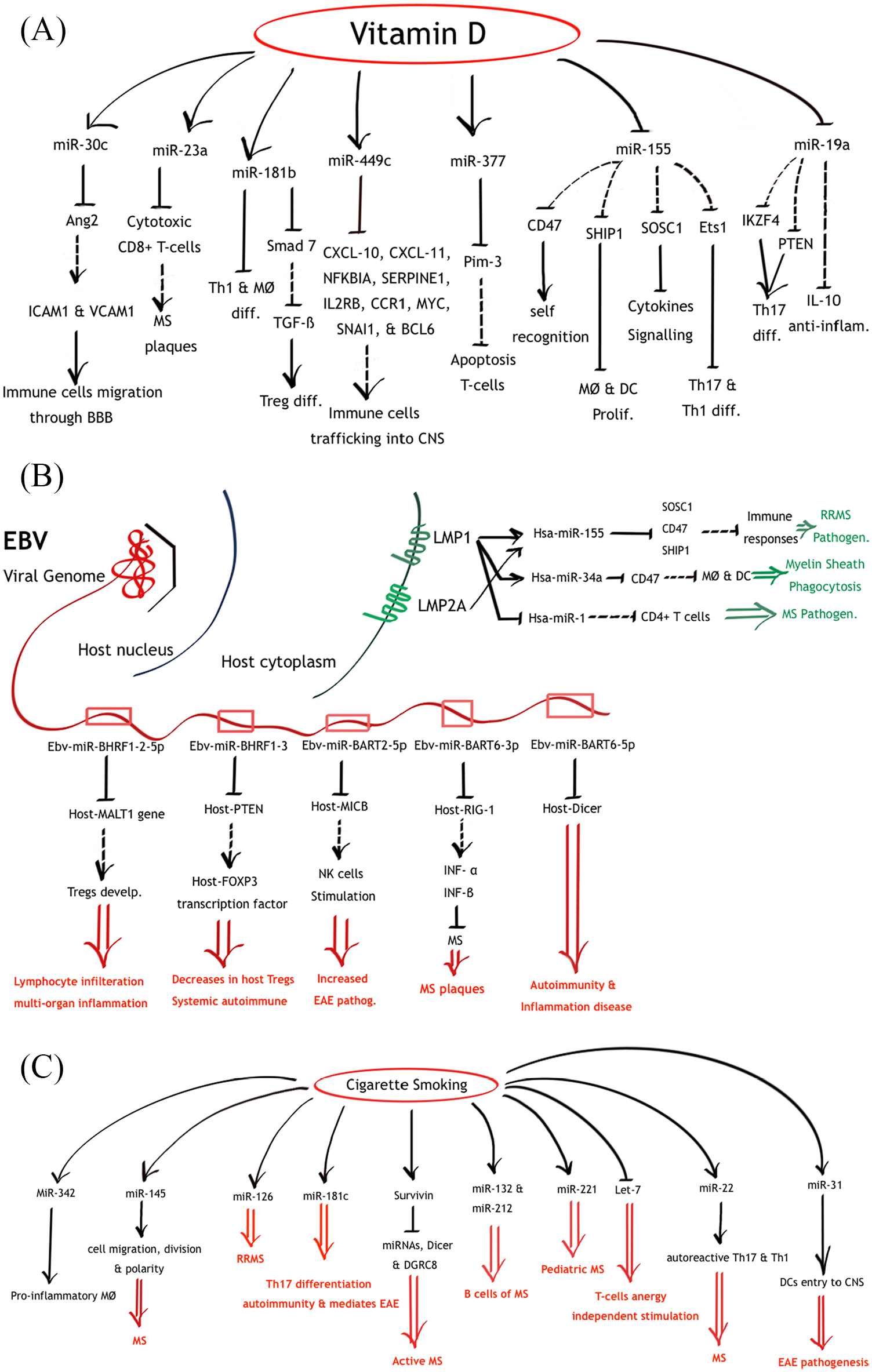
Illustration of miRNAs network dysregulations as a consequence of vitamin D supplementation (A), EBV (B), and cigarette smoking (C). Lines ending with arrows illustrate an induction process, while lines ending with hyphens illustrate an inhibition process. Dashed-lines illustrate pathways interrupted by environmental stimuli induced miRNAs dysregulations. Red double-arrows illustrate possible pathological outcomes according to previous studies.
Epstein-Barr virus (EBV) and miRNA
EBV infection is known to increase the risk of MS.3,223 Interestingly, the EBV virus encodes 44 conserved miRNAs located in two regions of the genome: the BART and the BHRF1 (BamHI fragment H rightward open reading frame 1). 224 These viral miRNAs have been previously linked to MS pathogenicity.
EBV-miR-BHRF1-2-5p and EBV-miR-BHRF1-3, for instance, were significantly increased in EBV-infected MS patients compared with those in healthy controls within a group of age- and sex-matched Chinese inhabitants. More so, the expression levels were associated with EDSS scores of MS patients (Table 2). 141
EBV-miR-BHRF1-2-5p can significantly suppress the MALT1 gene product. MALT1 has a vital role in the development of Tregs. Inactivation of MALT1 in mice was related to an absence of Tregs, Th1 and Th2 cell elevation, and increased interferon-gamma (IFN-γ) and IL-4 expression. Finally, it resulted in lymphocyte infiltration and multiorgan inflammation. 141 MALT1 has previously been linked to MS in one of the IMSGC studies, comprising 14 498 patients and 24 091 healthy controls. 163
EBV-miR-BHRF1-3 inhibits CXCL-11 and PTEN. CXCL-11 is an interferon (IFN)-inducible chemokine and PTEN is a key regulator of the phosphatidylinositol 3-kinase (PIK3)/AKT survival pathway. PTEN preserves the FOXP3 transcription factor and prevents Th1 and T follicular helper (TFH) cell responses. PTEN deficiency was related to decreased cell count of Tregs, increased IFN-γ, eventually, leading to systemic autoimmune disease.141,165
EBV virus may pose a higher risk of MS through other miRNAs. For example, miR-BART2-5p, encoded by EBV, is a negative regulator of the stress-induced molecule MICB. MICB binds the receptor NKG2D, which is a dominant activator of natural killer (NK) cells.165,225 Dysregulation of NK cell functionality has been associated with MS, and depletion of these cells can aggravate disease pathogenesis in animal models of MS. 226
The miR-BART6-3p, another EBV miRNA, targets the Retinoic acid-inducible protein 1 (RIG-1).227,228 RIG-1 activates type I IFN (IFN-α and IFN-β) cytokine production. These cytokines are proven to have a therapeutic effect on MS patients.165,229
EBV also encodes miR-BART6-5p, which was identified to target the human Dicer mRNA 3ʹ-UTR. 230 As previously mentioned, Dicer is important for maturation of miRNA within the cytoplasm before its incorporation into the RISC complex.23,26 The suppression of Dicer molecules may significantly affect the immune system. Deficiency in Dicer has been documented with aberrant B-cell receptor (BCR) signalling, induction of autoantibodies, and failure to discriminate activating and anergic stimuli. In addition, Dicer-deficient Tregs can provoke autoimmune and inflammatory diseases. 24
In addition, EBV can also induce dysregulation of host miRNAs, causing a profound host immune impairment. This virus significantly downregulated 99.5% of the host miRNAs population in infected B cells (median downregulation of 130.74-fold). However, 6 months post infection, 4 oncomirs (miR-17-5p, miR-20, miR-21, and miR-155) were significantly upregulated in EBV-positive cells compared with those in EBV-negative cells (Table 1). 108 Several studies have found an association of miR-21 and miR-155 elevation with EBV infection (Table 1).231-233 Interestingly, these miRNAs were previously documented to be elevated during the relapse stages of MS and EAE (Table 2).145,148,153,222
Furthermore, Godshalk et al have demonstrated that during the lytic cycle of EBV infection, several host miRNAs are upregulated, two of which are miR-146a and miR-146b. These are the key modulators of lymphocyte signalling; in particular, those targeting Toll-like receptor (TLR) and cytokine signalling pathways (Table 1). 108 miR-146a has been predicted to downregulate several MS-associated genes that play vital roles in the immune system, including CLEC16A, TRAF3, and IL7 (Table 3).160-163 Notably, miR-146a was upregulated in MS and EAE patients (Table 2).24,111,130,145,152,153,234
Interestingly, interaction between rs2910164, a polymorphism in miR-146a, and baseline anti-EBNA-1 IgG titres predicted a significant risk of conversion to MS (relative risk = 2.39, P = .00002). 142 This finding further highlights the association between EBV, miRNAs, and MS.
EBV also encodes the latent membrane proteins (LMP1 & LMP2A) that are known to induce the cellular miR-155 (Table 1).110,165,168,235,236 As discussed before, miR-155 targets SOSC1, CD47, and SHIP1 are negative regulators of immune responses. More so, miR-155 suppresses essential neurosteroids and can induce T-cell differentiation into the aggressive Th17 and Th1 cells.24,26,220,224 Also, miR-155 was significantly increased in RRMS cases (Table 2).2,130,145
Moreover, evidence associating the EBV viral protein, LMP1, to the suppression of host miR-1 is reported. LMP1 can increase the expression of phos-Stat-3 (the active form of Stat-3). Overexpression of Stat-3 was correlated with the inhibition of miR-1 promoter activity. A putative binding site for Stat-3 was uncovered using bioinformatics prediction tools in the upstream regions of the miR-1 gene (Table 1). 112 This miRNA was previously reported to be decreased in CD4+ T cells upon stimulation in RRMS (Table 2).25,109
LMP1 has also been reported to induce host miR-34a during primary B-cell infection (Table 1).109,168 The same miRNA has previously been associated with MS (Table 2).136,137 It targets CD47, a known inhibitor of MØ and DC cytokine synthesis. Upregulation of this miR-34a can release MØ from their inhibitory status and increase their phagocytic activity against myelin sheaths.2,25,136 miR-34a is also predicted to target several MS risk genes, including SLC2A4RG, MYB, FOXP1, TNFSF14, and OLIG3. These genes were identified as MS risk factors through the work of IMSGC, and some of them were further confirmed through GWS analysis. These genes encode transcription factors, regulators of haematopoiesis, costimulatory factors, and regulators of apoptosis (Table 3).160-163,166
EBV viral molecules not only can exert their effect on infected cells, but can also induce cellular deregulation in neighbouring uninfected cells via exosome-mediated transport of viral miRNAs and proteins. These neighbouring uninfected cells can either act as hosts or do not act as hosts to viral cells.165,228,237,238 Magliozzi et al 239 have detected immediate early (BZLF1) and early (BFRF1) EBV-proteins within cells of meninges adjacent to the cortical lesions infiltrated with immune cells in 2 of the 3 FFPE-blocks collected from post-mortem MS cases. Similarly, Serafini et al 240 reported a reactivity to EBV-encoded small RNA (EBER1 and EBER2) probes in sections adjacent to the immune cells–infiltrated white matter lesions.
EBV infection can constitute a potentially high source of MS risk in a host via miRNAs (Figure 3B); however, such a claim remains hypothetical until proven by more methodological studies.
Cigarette smoking and miRNA
Cigarette smoking has been identified as a risk factor for MS since the mid-1960s and has been linked to a 50% increased susceptibility to developing MS.3,94
Smoking can modulate miRNAs expressions in ways that affect vital body functions. In smokers, miR-145, miR-126, and miR-181c were differentially upregulated compared with those in healthy nonsmokers (fold-changes = 8.0, 5.0, 3.0 and P value = 1.2 × 10−3, 7.6 × 10−4, 1.3 × 10−3, respectively), along with other miRNAs. RNA was extracted from bronchoscopy samples of healthy nonsmokers, smokers, and smokers who quit smoking for 3 months. Interestingly, levels of miRNAs between smokers who quit smoking for 3 months and healthy nonsmokers were statistically insignificant (P value > .05), further confirming the effect of smoking on miRNA expression levels. The average miRNA index of the smoking cessation group was higher than that of healthy nonsmokers’ group by 2.4-fold, suggesting that disrupted miRNA levels may persist for a longer period (Table 1). 114 Interestingly, the miR-145, miR-126, and miR-181c were noted to be upregulated in MS cases (Figure 3C).
The miR-145 is one of the MS-associated miRNAs that can affect cell migration, division, and polarity (Table 2).138,234,241 miR-145 is predicted to target ELMO1, TET2, HHEX, and RPS6KB1, which are involved in cell growth, proliferation, and migration. Notably, these target genes have been associated with MS risk, as shown by the IMSGC group (Table 3).160-163 miR-126 was recently found to be significantly upregulated in RRMS cases.139,242 miR-181c could promote Th17 differentiation, induce autoimmunity, and mediate EAE and was upregulated in the peripheral blood and the CSF of MS patients.22,159,243
Furthermore, cigarette smoking could cause a global decrease in miRNAs profiles in alveolar MØ. 113 Smoking was further confirmed to cause a global reduction of miRNAs in alveolar MØ by the Gross’s group. According to their report, cigarette smoking altered the RNA endonuclease, Dicer, which causes defective miRNA processing. 244 In agreement with this study, smoking was found to induce high levels of the protein, survivin, in early rheumatoid arthritis (RA), which is associated with a restricted pattern and low production of miRNAs and decreased Dicer and DGRC8 production (Table 1).245,144 In fact, increased expression of survivin is a feature of active MS cases (Table 2).125,126,246
Smoking could also upregulate the expression of miR-132 and miR-221 and downregulate that of Let-7b in alveolar MØ (Table 1). 113 miR-132 was upregulated in B cells of MS patients and lymph nodes of EAE models (Table 2).153,247-249 miR-221 was found to be upregulated in paediatric MS patients (Table 2). 147 miR-221 targets ZMIZ1, a gene involved in the regulation of various transcription factors, previously shown to be associated with MS in the IMSGC study (Table 3).160,161,163 In contrast, Let-7 downregulation results in T-cell-evoked anergy-induced stimulation, a feature characteristic of MS. 24 Meanwhile, Let-7d and Let-7c, members of the Let-7 family, have been reported to be upregulated in MS and EAE.131,153,168,234,250,251
Cigarette smoking could induce autoreactive Th17 and Th1 cells in the lungs. This proinflammatory reaction is mediated by the induction of miR-22 expression of APCs (Table 1).117,143 Indeed, the upregulation of miR-22 was reported in Tregs, plasma, blood cells, PBMCs, and brain white matter tissues of MS patients (Table 2).27,129-131,252 It is also predicted that miR-22 can suppress the expression of VCAM1, CD86, IL22RA2, GPC5, CLEC16A, and CD40 genes; all of which were proven to be associated with MS in several GWS studies (Table 3).160,162-164 These genes play important functions in immune responses, cell adhesion, cell signalling, and stimulation (Table 3). 161
Smoking is also implicated in the upregulation of miR-342, which is known to distinguish RRMS and progressive MS from controls and RRMS from progressive MS (Tables 1 and 2).118,127 miR-342 has been shown to promote inflammatory activation of MØ. 118 Interestingly, this miRNA can target several genes, including TNFRSF6B, TRAF3, and KIF21B, which was proved to pose a risk towards MS in the IMSGC study (Table 3).160,162,163 TNFRSF6B encodes a protein involved in the suppression of FasL- and LIGHT-mediated cell death. TRAF3, as mentioned earlier, mediates signal transduction in members of the TNFR superfamily. KIF21B is involved in the intracellular transport of membranous organelles (Table 3). 161
In a different study, cigarette smoke-induced inflammation could result in the upregulation of miR-31, miR-155, and miR-218 and downregulation of Let-7c in chronic obstructive pulmonary disease (COPD) (Table 1). 116 miR-31 was enriched in DCs in the CNS of EAE and was proposed to mediate their entry into the CNS during EAE pathogenesis (Table 2). 156 miR-31 targets a number of MS pathogenic genes: HLA-A, TNFSF14, JAZF1, and ZFP36 L1 (Table 3).160-163,253 Furthermore, miR-218 was documented to be upregulated in EAE lymph nodes (Table 2). 153 Let-7c was dysregulated in the blood of MS patients and also targeted some of the high-risk MS genes, including KIF21B, TET2, and GALC, as indicated in the IMSGC study (Tables 2 and 3).160-163,168
Also, cigarette smoking could downregulate miR-21-5p in SPMS, more specifically in the relapse phases (Tables 1 and 2).115,139,193 This miRNA could target numerous MS risk genes, including SLC2A4RG, MALT1, STAT3, TAGAP, DDAH1, and IL12A, as indicated by the GWS and IMSGC studies (Table 3).160-163,166,167
Notably, maternal tobacco smoking could induce miR-223 upregulation in the blood, which subsequently affected Treg counts and functionality (Table 1). 119 miR-223 was recorded to be increased in MS and EAE cases, especially in the relapse phases of the disease (Table 2).146,150 This miRNA has previously been predicted to target HLA-DPB1, IL2RA, and WWOX, which are also MS risk genes (Table 3).160-162,254,255
Maternal cigarette smoking was also shown to induce downregulation of foetal miR-16, miR-21, and miR-146a. 256 miR-146a-deficient mice were observed to develop a more severe form of EAE (P value < .01; fold-change < −1). 257 Cigarette smoking could also deregulate many miRNAs involved in immune system haemostasis that could influence the risk of MS. 258
Explaining how cigarette smoking influences miRNAs has partially clarified some issues regarding this subject. However, until proven through methodological studies, the link among cigarette smoking, miRNAs, and MS pathogenicity remains hypothetical.
Environmental factors-miRNAs-MS pathogenicity
Analysing the effect of the environmental factors on the miRNA populations parallel to MS miRNA dysregulated populations can further highlight the possible associations among environmental factors, miRNAs, and MS (Table 4). miR-155 was found to be upregulated by 2-fold in MS and ~10-fold in EAE.145,150 Vitamin D supplementation can decrease this miRNA by 2-fold, while EBV infection and cigarette smoking can increase this miRNA by 1.6-8.36 and 1.58-folds, respectively.108,110,111,116,121 miR-21 was highly upregulated in EAE (90-fold) and slightly upregulated in EBV infection (2-fold).108,148 The EBV infection may not be completely responsible for the upregulation of this miRNA but could be one of the influencers. miR-21-5p was found to be downregulated in RRMS cases by 3.96-fold 139 and was also downregulated in cigarette smokers by 3.14-fold. 115 miR-19a was upregulated in relapse stages of MS by 2-fold. 137 Vitamin D supplementation was documented to decrease this miRNA by 2-fold. 122 Although Ghorbani et al 134 demonstrated a downregulation of miR-181b in both MS and EAE (less than 2-fold); Bergman et al 153 documented upregulation of this miRNA in EAE models (1.2-fold). Vitamin D supplementation was revealed to increase the expression of this miRNA by 1.58-fold. 120 miR-23a was found to be underexpressed in MS cases (0.41- to 6.25-fold).123,146 and was upregulated by vitamin D supplementation by 1.37-fold. 120 miR-30c under-expression in MS cases (6.6-fold) could not be corrected by vitamin D supplementation, which only elevated this miRNA by 1.33-fold.120,123 miR-146a was elevated in MS cases by 2-fold, 111 although the levels of miR-146a in MS did not reach to those predicted in EBV infection, which was up to 112-fold. 108 This may indicate the presence of other influencers that may affect this miRNA. miR-34a expression was raised in both RRMS- and EBV-infected patients, though not to the same extent (1.3- to 3-fold and 10-fold, respectively).109,136,137 miR-146b was upregulated in EAE to levels comparable to those found in EBV infection (1.9 and 2.2-folds; respectively).108,153 miR-1 was downregulated in EAE and EBV infection, but not to the same extent (3.4 and 1-folds; respectively).112,136 Cigarette smoking could increase miR-132 up to 78-fold; however, this could not explain the 1.54-fold increase in EAE lymph nodes.113,153 Also, cigarette smoking could not explain the minor elevation of miR-221 and miR-145 in MS patients (fold-changes in MS patients = 1.6 and 3.15-folds; in cigarette smokers = 12.59 and 8-folds, respectively).113,114,138,147 miR-218 and miR-22 were elevated in MS and EAE to levels similar to those found in cigarette smokers.116,117,129,153 Survivin expressions in MS (4.5-fold) could not be attributed to cigarette smoking only (1.79-fold).125,144 Levels of Let-7c in MS did not correlate with those found in cigarette smokers (-1.45-fold).116,138 miR-342 was upregulated by 3.2-fold in MS patients, half of it could be attributed to cigarette smoking (1.51-folds increase).118,127 Nonconclusive results of miR-31 in EAE models make it hard to correlate to the results of cigarette smoke.116,153,156 miR-223 was upregulated in blood and serum of MS patients by 0.8- to 2.08-fold and in spinal cords and peripheral immune system of EAE by up to 30-fold.138,140,146,150 However, cigarette smoking could upregulate this miRNA in Tregs by only 1.04-fold. 119 Taken together, environmental factors may influence some of the miRNA populations which could result in MS or EAE. However, miRNAs dysregulation could not be attributed to environmental factors only; other factors may also play a role.
MS therapy and miRNA
To date, it is clear that miRNA disruption may play a role in MS. The next step was to investigate whether such miRNAs are linked to MS treatments and whether such drugs can re-regulate the disrupted miRNA expressions.
Current FDA (Food and Drug Administration)-approved drugs have been shown to regulate some of the MS-associated miRNAs. An example is glatiramer acetate, which significantly decreases the expression of miR-146a and miR-142-3p in RRMS compared with that in normal (P = .028, P = .003, respectively) (Table 5).145,259 As discussed previously, miR-146a is one of the miRNAs associated with MS and RRMS and is associated with different environmental stimuli. Moreover, it affects several MS-associated genes (Tables 2 and 3).24,111,142,145,152,153,160-163,176,223,227,234
Another example is the interferon-beta (IFN-β), which Waschbisch et al have excluded to play a role on miRNAs of RRMS patients. In contrast, Hecker et al have extensively studied and documented a significant influence of IFN-β on several miRNAs in patients with clinically isolated syndrome (CIS) and RRMS (Table 5).145,169,259 IFN-β could affect the expression of 20 miRNAs 1 month after the first injection. Of these, miR-29c-3p and miR-532-5p were confirmed to be downregulated in MS patients using TaqMan miRNA arrays and Affymetrix miRNA arrays in 2 different data sets (P < .001, P = .048; respectively), confirming the reliability of the results (Table 5). 169
De Felice et al had also studied the effect of IFN-β treatment on 40 RRMS patients. The analysis showed a differential expression of several miRNAs, including miR-326, miR-155, miR-3676, miR-18b, miR-599, and miR-26a-5p. In particular, the expression of miR-29a-5p underwent the most significant alteration post IFN-β treatment, 3 and 6 months after therapy (Table 5). 170 Furthermore, as detailed previously, miR-155 was associated with MS and MS environmental stimuli (Table 2).108,110,116,121,130,145,235
Likewise, Ehtesham et al demonstrated that IFN-β can restore the expression levels of the previously described MS- or EAE-associated miRNAs: miR-145 and miR-20a (Tables 2 and 5).24,122,138,171,234,241,260
Similarly, Manna et al studied 16 differentially expressed miRNAs in IFN-β treated RRMS patients vs naive patients. Of these 16 miRNAs, 2 (miR-22-3p and miR-660-5p) were upregulated and 14 (miR-486-5p, miR-451a, miR-let-7b-5p, miR-320b, miR-122-5p, miR-215-5p, miR-320d, miR-19b-3p, miR-26a-5p, miR-142-3p, miR-146a-5p, miR-15b-3p, miR-23a-3p, and miR-223-3p) were downregulated (Table 5). 172 Some of these miRNAs, namely, miR-22, miR-320b, miR-146a, miR-23a, miR-142-3p, miR-19b and miR-223, have previously been documented to be associated with MS or EAE pathogenicity (Table 5).24,27,111,122,123,129-132,135,138,145,146,150,152,153,158,234
Natalizumab, another FDA-approved drug for MS treatment, was similarly studied for its effect on miRNAs. Meira and her colleagues found miR-126 and miR-17 to be downregulated in CD4+ T cells of RRMS patients upon treatment with natalizumab (Table 5).173,174,259 In another study by Sievers et al, ten differentially expressed miRNAs related to natalizumab treatment were found in RRMS patients, with two clusters standing out: miR-106b-25 and miR-17-92. More so, they found two deregulated EBV miRNAs (miR-BART3-5p and EBV-miR-BART11-5p) in the untreated RRMS patients compared with those in treated patients and healthy controls (Table 5). 175
In agreement with Meira et al, Ingwersen et al 176 found four miRNAs to be upregulated in MS patients treated with natalizumab: miR-18a, miR-20b, miR-29a, and miR-103. In contrast, one miRNA (miR-326) was downregulated (Table 5). Furthermore, Munoz-Culla and colleagues found three miRNAs (let-7c, miR-125a-5p, and miR-642) to be differentially expressed 6 months post-natalizumab treatment in 19 RRMS patients (Table 5). 132 Augmentation of the miR-17-92 cluster, which includes miR-18a, is associated with EAE pathogenicity (Table 2).24,122 As previously mentioned, Let-7c is associated with RRMS (Table 2). 138
Besides, Mameli et al reported that natalizumab could downregulate miR-155 and miR-26a, miRNAs that had previously proven to increase MS pathogenicity (Tables 2 and 5).130,145,177
Dimethyl fumarate (DMF), another FDA-approved MS treatment drug, has similarly shown efficacy in reducing the expression of the pathogenic miR-155 (Table 5). 178
Fingolimod (FGM), also FDA-approved MS drug, proved to significantly reverse the aberrant miR-15b, miR-23a, and miR-223 expression after 6 months of the administration. Promisingly, the levels of expression remained stable throughout the remaining year of treatment (Table 5). 179 In another study, several miRNAs were significantly differentially expressed only 5 hours post administration (P < .05). Of these, 46 were overexpressed (fold-change > 2) and 33 were underexpressed (fold-change < −2). Analysing their target genes revealed several pathways related to MS pathogenicity. Among these pathways, tumour necrosis factor-related apoptosis-inducing ligand (TRAIL) signalling, beta 1 integrin family cell surface interactions, epidermal growth factor receptor (EGFR)-dependent endothelin signalling, Arf6 signalling, and S1P1 were the most prominently discerned. 157
Nanocurcumin, a drug still in phase 2 clinical trials, has been postulated to regulate a number of MS-associated miRNAs, including, miR-16, miR-17-92, miR-27, miR-29b, miR-126, miR-128, miR-132, miR-155, miR-326, miR-550, miR-15a, miR-19b, miR-106b, miR-320a, miR-363, miR-31, miR-181c, miR-150, miR-340, and miR-599 (Tables 2 and 5).22,24,122,124,130,139,145,148,153,156,159,180,188-192,242,243,247-249
Interestingly, autologous hematopoietic stem cell transplantation (AHSCT) was also able to restore normal levels of miR-16, miR-155, and miR-142-3p 6 months post therapy in 24 MS patients. 181
Current therapeutic approaches can potentially restore some disrupted miRNAs to their normal levels. However, the restoration is not complete. Therefore, only some symptoms in patients, but not all, are ameliorated. Consequently, such medications are termed ‘disease-modifying drugs’.
Environmental risk factor modifying drugs and miRNA
The next step in this review is to identify the effects of environmental risk factors modifying drugs or diet on the distrusted miRNA populations.
Vitamin D supplementation and miRNA
Karkeni et al demonstrated the effect of TNFα (a signal protein involved in systematic inflammation) on the expression of miR-146a, miR-150, and miR-155 in the human adipocytes,182,261 which are some of the MS dysregulated miRNAs, as discussed earlier in this article. In a report by Karkeni et al, incubation of 1,25(OH)2D with the human adipocytes dramatically decreased the expression of the aforementioned miRNAs. Moreover, the levels of these three miRNAs were significantly reduced in mice fed with a high-fat diet supplemented with vitamin D, while their levels were significantly increased in those fed with only a high-fat diet (Table 5). 182 These results indicate the effect of vitamin D supplementation on modulating the expression of some MS dysregulated miRNAs.
Zeitelhofer et al demonstrated the effectiveness of vitamin D supplementation on the regulation of the impaired miRNAs involved in the EAE development. In their report, 92 miRNAs expression levels in CD4+ T cells were re-regulated by vitamin D supplementation. These miRNAs were pertinent to TCR and IL-2 signalling (Table 5). 120
Pedersen and his colleagues have shown the effect of vitamin D3 incubation on DC. The DCs treated with vitamin D3 resembled immature DCs, except for high CD14 and reduced CD1a on their cell surface. These cells also have presented long-lasting inefficient T-cell stimulation and induced T-cell hyporesponsiveness with a regulatory potential. Interestingly, the DCs treated with vitamin D3 had low levels of IL-23 secretion and low expression of miR-155 when exposed to maturation stimuli. The treated DCs also showed overexpression of miR-378 (Table 5). 183 The maturation of DCs is vital in the process of MS development. Such cells present myelin antigen in vivo to stimulate myelin-reactive T cells and mediate CNS inflammation. 262 miR-155, as stated, was upregulated in RRMS cases and more importantly in the white matter active lesions (Table 2).145,221 miR-155 targets genes involved in inhibiting cytokines signalling and T-cell differentiation, and genes involved in self-recognition and protection against phagocytosis.24,26,121,162
A study by Giangreco et al indicated the effect of vitamin D supplementation on the miR-100 and miR-125b expression levels in primary prostatic epithelial cells. Primary prostatic epithelial cells were collected from 45 patients enrolled for a vitamin D randomized trial. In the patients, primary prostatic epithelial cells had lower expression levels of miR-100 and miR-125b than that found in benign prostate cells. Incubation of vitamin D with these cells resulted in an augmentation of the expression levels of miR-100 and miR-125b (Table 5). Furthermore, vitamin D incubation suppressed the expression levels of E2 F3 and Plk1, targets of miR-100 and miR-125b. 184 Augmentation of miR-125b seems to suppress the prostate cancer pathophysiology; on the contrary, the same miRNA seems to promote EAE pathophysiology (Table 2). 153
A study by Nunez Lopez et al 185 demonstrated the effect of vitamin D supplementation on circulating levels of miR-7, miR-152, and miR-192 (Table 5). miR-7 and miR-192 levels were reduced by 6- and 1.7-fold after 4 months of daily supplementation with 2000 units of cholecalciferol, while miR-152 level showed a significant increase after daily supplementation with the 2000 units of cholecalciferol (P = .03; fold change = 1.5). Notably, miR-7 was observed to be increased in EAE lymph nodes (P = 1.2E-03, fold change = 1.4) (Table 2). 153
In an article by Mansouri et al, 186 daily supplementation of 2 μg paricalcitol in 12 patients with chronic kidney disease (CKD) significantly reduced the levels of miR-432-5p, miR-495-3p, and miR-576-5p compared with those in the placebo group (n = 12). miR-432-5p was reported to be a significantly dysregulated miRNA that could distinguish between healthy controls and progressive MS and between progressive MS and RRMS (P = .002, 0.051, fold-change = 2.2, 0.5; respectively). 127
EBV drug and miRNA
Lukiw et al 187 have documented the effect of Acyclovir (ACV) drug on the miR-146a levels; however, their work was performed on HSV-1 virus (Table 5). ACV is a nucleoside analogue that selectively inhibited viral but not cellular replication. 263 Lukiw et al 187 reported that ACV could significantly quench the upregulated miR-146a associated with HSV-1 (P < .01) (Table 5). miR-146a has been predicted to downregulate several MS-associated genes and is noted to be upregulated in MS patients (Tables 2 and 3).24,111,145,152,153,160-163,234
Cigarette smoking drug and miRNA
Varenicline (Chantix) is an FDA-approved drug for smoking cessation. This drug acts as a partial nicotinic acetylcholine receptor agonist. 264 This drug was effective in reducing smoking-dependent miRNA index in 10 smokers after quitting for 3 months (from 3.8-fold to 1.6-fold change; P value < .05). However, the smoking-dependent miRNA index was upregulated in smokers compared with that in healthy nonsmokers (3.8-fold; P value < .05) (Table 5). 114
Conclusion
MS is a complex autoimmune disease that poses a challenge to the medical field. To understand such a disease, the molecular mechanisms in cells must be understood in details. To our current knowledge, MS is a disease with predisposing genetic variants and environmental influencers. Environmental factors may influence disease pathology through different mechanisms, one of which is speculated to be miRNAs.
This review has attempted to illustrate the influence of different environmental stimuli on the miRNA networks pertinent to MS pathology. Nevertheless, it must be emphasized that the data reproducibility has been a major limiting factor. Such a limitation could be due to the following reasons: (1) Although miRNAs were first discovered in 1993 by Lee and colleagues, it was not until 2008 that it was first studied in MS or EAE.33,265 (2) Vitamin D deficiency, EBV virus, and cigarette smoking have been recently proven to be risk factors for MS. Belbasis et al have extensively reviewed the literature and conducted 44 meta-analyses with 416 primary studies of different MS risk factors. In their article, they concluded that EBV and smoking were among the highest MS risk factors, while vitamin D deficiency had lower effects. 3 (3) The thriving of new ideas as a result of advanced scientific technology has made data reproducibility much harder. Fewer researchers are interested in exploring the influence of environmental factors on miRNAs concerning MS pathogenicity,120,141,142,175 this subject matter seems to be lost over the years because of newer technological and biological discoveries. (4) Scientific knowledge can be expensive and time-consuming, and if it does not lead to interesting finding, it could be forgotten. Thus, there is a need for scientific tools to help predict possible outcomes of a study and to direct researchers to the best scientific paths. Such tools could be literature reviews, meta-analysis studies, or bioinformatics databases.
Environmental – miRNA – MS axis could be explored through methodological analysis consolidated with literature reviews, meta-analysis studies, and bioinformatics databases. Bioinformatics tools help in the prediction of the transcription, regulation, and functionality of genes, epigenetic markers, and proteins. The use of such tools has been implicated in many analytical studies to guide investigators in exploring the properties, targets, or functionalities of the analysed components. These tools can help in understanding the complex nature of MS disease, especially when dealing with complex components such as the miRNAs. For example, in an article by Zeitelhofer et al, the authors have investigated the effect of vitamin D in regulating the distributed miRNA populations in an EAE-induced model, which resulted in EAE amelioration. The genes targeted by such miRNA populations were then explored using bioinformatics tools. Employing a literature review, the targeted genes were found to be involved in the activation and differentiation of T cells. Thus, the team investigated the targeted gene expression levels in these models, which were observed to be affected by miRNAs regulation. To further support the vitamin D – transcriptome – MS axis hypothesis, the investigators conducted adoptive transfer experiments in DA rats using MBP63–88-specific T-cell lines treated with vitamin D. These rats demonstrated lower proliferation and differentiation of CD4+ T cells, and the epigenetics markers and predicted associated genes were downregulated. 120 Another notable example of the use of literature reviews, meta-analysis and bioinformatics as tools to enrich scientific work is the article by Teymoori-Rad et al. In this article, the authors have first explored different search databases to find suitable microarrays data sets (literature review). Next, the data sets were used to identify differentially expressed miRNAs (DEMs) through meta-analysis. With the help of different bioinformatics tools, the targets of the DEMs, protein-protein interaction networks, and signalling pathways from RRMS patients were proposed to be associated with vitamin D deficiency and EBV infection. Thus, literature reviews have connected the dots between the presented data. Although the interaction among vitamin D /EBV, miRNAs, and MS was constructed, based on a meta-analysis, bioinformatics proposed data, and literature reviews without the support of clear methodological data, it still helped to clarify the complex nature of MS pathogenicity. 234 Nonetheless, methodological studies must be conducted to confirm the proposed pathogenic paths in the MS disease.
It should be noted that few investigators have studied the environmental-miRNAs-MS axis.120,141,142,234 Zeitelhofer et al, in their article, have pointed to the effect of vitamin D supplementation in regulating the disrupted miRNAs population in CD4+ T cells, which were vital to CD4+ T cell proliferation and differentiation. The regulation of such miRNAs resulted in lower proliferation and differentiation of CD4+ T cells, leading to amelioration of EAE. 120 Teymoori-Rad et al reviewed literature for transcriptome data sets that were differentially expressed between RRMS and healthy controls. A meta-analysis of the DEMs revealed 9 miRNAs as the best candidates for significant dysregulation. The 9 miRNAs were then analysed through different bioinformatics databases to define the underlying protein–protein interaction networks and signalling pathways. The analysis indicated the role of these miRNAs in many vital pathways and emphasized the possible interaction between vitamin D/EBV with MS disease. 234 Wang et al conducted a study on 30 RRMS patients and 30 age- and sex-matched healthy controls from the Chinese population. In their article, they reported significant expression levels of ebv-miR-BHRF1-2-5p and ebv-miR-BHRF1-3 in the circulation of RRMS patients. Furthermore, they found a positive correlation between the ebv-miRNAs and EDSS scores of MS patients. The ebv-miR-BHRF1-2-5p was confirmed to target MALT1 gene, which is a key regulator of immune homeostasis. 141 Furthermore, Zhou et al 142 demonstrated a significant interaction between a genetic polymorphism in the miR-146a (rs2910164) and baseline anti-EBNA-1 IgG titres in predicting the risk of conversion to MS (relative excess risk due to interaction [RERI] 2.39, P = .00002) and relapse (RERI 1.20, P = .006).
Although the data available on miRNAs seem encouraging, unfortunately, they are collected from only a few studies showing only nominal P values. Most of such associations seem to be false positive due to the insufficient power of the studies and the compounding effects of other factors, such as disease course, disease activity states, treatment, cell types analysed, use of animal models, etc. However, the available data points to the need to explore such hypotheses through methodological studies, with an aim to further understand the complexity of the MS disease.
Nevertheless, through literature review some miRNAs have stood out as particularly associated with MS risk, since they have been validated across different studies to be linked with MS, MS subtypes, or EAE. Such miRNAs include miR-155, miR-23a, miR-34a, miR-22, miR-21-5p, miR-181b, miR-221, miR-223, miR-1, and survivin (Table 6). Interestingly, the validated miRNA to be deregulated in response to different environmental stimuli was limited to miR-155 (Tables 4 and 6). miR-155 was a major factor that linked vitamin D, EBV, and/or cigarette smoking to MS. Since this miRNA plays a vital role within the immune system, it is of particular concern. As mentioned, miR-155 targets SOSC1, a negative regulator of cytokines signalling, induces T-cell differentiation in the Th17 and Th1 cell lines, and targets transcription factor Ets1 (a negative modulator of Th17 differentiation).24,121,219 It also targets CD47, a molecule involved in self-recognition and protection of cells from phagocytosis, and suppresses SHIP1, a known negative inhibitor of proinflammatory pathways related to MØ and DCs.24,26 More so, miR-155 can suppress essential neurosteroids in the white matter of MS patients.26,220 Importantly, miR-155 targets several genes associated with MS pathogenicity, such as IL22RA2, TRAF3, IRF8, DDAH1, MYB, GALC, and RPS6KB1 (Table 3).160-163,166
Current therapeutic approaches exert a notable impact on some of the miRNAs associated with MS. FDA-approved drugs have shown their efficiency in regulating some of the known MS-associated miRNAs, namely, miR-146a, miR-155, miR-145, miR-20a, miR-22, miR-320b, miR-146a, miR-23a, miR-223, miR-BART3-5p, and EBV-miR-BART11-5p.132,145,170-172,175,177,178,259 However, in the future, novel approaches need to focus on designing drugs that modulate the miRNA networks as a target. This approach may need more time, expense, and research, but could provide a better life for these patients. At present, the prescribed therapeutic interventions are suggested through experimental trials of known available medications in the market rather than targeting MS aetiological factors. Consequently, this kind of therapy is described as ‘disease-modifying drugs’.
Vitamin D supplementation has been proven to modulate some of the miRNAs posing a risk towards MS, such as miR-146a, miR-150, miR-155, miR-30c, miR-23a, miR-181b, miR-449c, miR-125b, miR-7, and miR-432-5p.120,182-186 It was also linked to EAE amelioration. 120 The EBV drug, ACV, was related to a significant downregulation of MS-associated miR-146a. 187 Varenicline (Chantix) prescribed for smoking cessation was effective in reducing the smoking-dependent miRNA index in 10 smokers who quit smoking for 3 months. 114 Hence, implementing vitamin D supplementation, treating EBV, as well as advising cessation of cigarette smoking in the MS therapeutic plan must be considered for susceptible individuals.
This review has focused on the role of miRNAs in mediating the influence of environmental factors on MS pathogenicity; however, environmental factors have been documented to influence MS by other epigenetic means. For instance, in an article by Zeitelhofer et al, not only did vitamin D supplementation regulate the miRNAs population, but it also affected the DNA methylation in CD4+ T cells (P < 2 × 10−24, <.001; respectively). The DNA methylation seemed to affect the miRNA genes; miRNA expression levels regulated the protein levels and thus ameliorated the EAE. 120 In another study, Moore et al 266 demonstrated the effectiveness of vitamin D in regulating the DNA methylation process, which, in turn, directed the transition of CD4+ T cells to Treg cells. Smoking was documented to affect DNA methylation in MS patients and the effect was proportional to the smoking load. DNA methylation could affect gene expression levels, of which the AHRR gene was confirmed to be highly expressed after smoking in MS patients (P = 2.10 × 10−3). 107 Although EBV-induced DNA methylation is not documented in MS patients, it has been recognized in several other diseases, such as nasopharyngeal carcinoma, Hodgkin lymphoma, Burkitt lymphoma, and gastric carcinoma.267-272
miRNAs signatures in MS patients may be defined by the environment and genetics. However, data from diverse groups, ethnicities, and countries need to be consolidated and reproduced to be accepted as robust. In conclusion, until methodological studies can prove the relationships demonstrated among environment, miRNAs, and MS, this review remains a mere hypothesis.
Over the last few decades, the biology of MS has become much clearer. Nevertheless, much work has to be done in the future to elucidate the aetiology of MS, to ultimately lead to better therapeutic interventions and improve the quality of life of MS patients.
Supplemental Material
Supplementary_table_1 – Supplemental material for Environmental Influencers, MicroRNA, and Multiple Sclerosis
Supplemental material, Supplementary_table_1 for Environmental Influencers, MicroRNA, and Multiple Sclerosis by Eiman MA Mohammed in Journal of Central Nervous System Disease
Footnotes
Acknowledgements
The author dedicates this work to her family, who always support, encourage, and fight for her. The author is deeply thankful and blessed for the special, unique, and amazing people in her life, the Molecular Genetics Laboratory staff; Dr Shakir Bahzad, Mona AlAteeqi, Hiba Jama, Noor AlRumaidhi, and Jumana Rashid, who always support, uplift, comfort, and bring joy to her soul. The author thanks Editage (![]() ) for English language editing.
) for English language editing.
Funding:
The author received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests:
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
EMAM alone is responsible for the content and writing of this article. The author has read and approved the final manuscript.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.