
Abstract
Colorectal cancer (CRC) is the third most common cause of cancer mortality globally. Development of CRC is closely associated with lifestyle, and diet may modulate risk. A Western-style diet is characterised by a high intake of red meat but low consumption of fruit, vegetables, and whole cereals. Such a diet is associated with CRC risks. It has been demonstrated that butyrate, produced by the fermentation of dietary plant fibre, can alter both genetic and epigenetic expressions. MicroRNAs (miRNAs) are small non-coding RNAs that are commonly present in both normal and tumour cells. Aberrant miRNA expression is associated with CRC initiation, progression, and metastasis. In addition, butyrate can modulate cell proliferation, differentiation, apoptosis, and miRNA expression in CRC. In this review, the effects of butyrate on modulating miRNA expression in CRC will be discussed. Furthermore, evidence on the effect of butyrate on CRC risk through reducing oncogenic miRNA expression will be presented.
Introduction
Cancer is the leading cause of death in both developing and developed countries. 1 This may be related to population ageing and lifestyle risk factors such as smoking, low levels of physical activity, energy imbalance, and unhealthy diets. 2 Colorectal cancer (CRC) is the third most common cause of cancer mortality and constitutes about 10% of all cancers globally. 2 The highest incidence rates of CRC are in developed countries, such as the United States and the United Kingdom, and lifestyle, rather than genotype, is regarded as the most important risk factor.3,4
Colorectal cancer risk is closely related to the consumption of dietary types.5,6 There is evidence to support the notion that people who consume Western-style diets have a greater CRC risk than those who consume a Mediterranean- or Asian-style diet.7–10 Nowadays, Western-style diets contain high levels of energy-dense foods, red and processed meat, and a low consumption of fresh fruits, vegetables, and whole grains. In contrast, traditional Mediterranean- and Asian-style diets include a variety of plant-based foods and unrefined cereals and a lower consumption of red and processed meat.7,10
Plant-based foods are usually high in dietary fibre, which has been shown to decrease the risk of CRC in both human and animal studies.11–14 Dietary fibre in the colon, following fermentation, is the source of short-chain fatty acids (SCFAs) which are beneficial for gut health. 15 The SCFAs can help protect against DNA damage and mutations, 16 both of which are associated with increased cancer risk. Acetate, propionate, and butyrate are the most common SCFAs, with butyrate being associated with protection against aberrant crypt foci (in rats). 17 In addition, the increase in butyrate production may reduce CRC risk by regulating gene expression. 18
Gene expression in both healthy and malignant tissues can be modulated by epigenetic mechanisms, such as DNA methylation, histone acetylation, and by the presence of micro- RNA (miRNAs). Epigenetics is defined as the regulatory processes which modulate the transcription of information encoded in the DNA sequence into RNA before their translation into proteins. 19 Epigenetics offers an explanation of the phenome that identical DNA sequences can lead to diverse phenotypes and varying disease susceptibilities.10,20 In this review, the effects of butyrate on inhibiting carcinogenesis in the colon, through modulating miRNA-related gene expression, are presented and discussed.
Butyrate and Risk of CRC
The risk of developing CRC is closely correlated with dietary intake as dietary factors can influence carcinogenesis in the gut.9,21 Red and processed meat, particularly the cooking of red meat at high temperatures, can cause high levels of N-nitroso compounds (NOC) and haem iron in the colon. These compounds are common carcinogens that may lead to increased levels of DNA damage. 22 Butyrate can help ameliorate the harmful effects of red meat consumption. This notion is supported by studies conducted by Le Leu et al23,24 who conducted an animal study and a human trial to test the effect of a butyrylated high-amylose maize starch (HAMSB) diet on O(6)-methyl-2-deoxyguanosine adducts in rectal tissue. The authors demonstrated that butyrate can decrease the toxic influence of a cooked red meat diet on colon cells. 24 Also, high consumption of red and processed meat may increase the proportion of pathogenic bacteria and decrease the proportion of beneficial bacteria in the gut. This imbalance may cause inflammation in the colon, 5 leading to a more carcinogenic environment. The high intake of fresh fruit and vegetables, foods high in antioxidants, can reduce DNA damage to colon cells resulting from reactive oxygen species (ROS), 22 and in this way, the negative impact of NOC, haem iron, and ROS can be modulated.
Beneficial effects of fibre, and the inverse relationship between fibre intake and premature death from diseases, have led the World Health Organization to establish a recommended daily intake (20 g of fibre per 1000 kcal consumed) which, unfortunately, is generally not achieved in Western countries. 15 Dietary fibre consists of polysaccharides, such as indigestible starch, pectin, cellulose, and gums. Partially digested or undigested polysaccharides are transited into the large intestine where they are fermented by non-pathogenic microbiota to produce SCFAs. 25 The most common SCFAs produced in the gut are acetate (C2), propionate (C3), and butyrate (C4). 15 Acetate and propionate reach the liver via the portal vein, and butyrate is predominantly utilised by colonocytes. 15 Resistant starch (RS) granules are fermentable polysaccharides that are naturally rich in amylose, and following fermentation, RS gives rise to high levels of butyrate production.
It is not practical to collect colon biopsy samples in free-living individuals. For this reason, butyrate levels are usually determined from faecal samples (dry weight) as this sample type has been shown to represent colonic values. 26 Butyrate levels were found to vary as much as 10-fold in a healthy population depending on body mass index, microbiota, and other factors.26,27 Faecal butyrate concentrations increased significantly following the consumption of RS.26–28
There are 2 major groups of butyrate-producing bacteria, namely, Faecalibacterium prausnitzii and Eubacterium rectale/Roseburia spp and the type and ratio of microbiota in the gut can influence the amount of butyrate produced.25,29 In addition to the microbiota, dietary intake can influence the production of butyrate. For example, cellulose, lignin, and some insoluble fibre have low ferment ability, so the production of butyrate from these sources is low. 30 Besides dietary fibre, other butyrate substrates, such as the oligosaccharides acarbose and tributyrin, can increase butyrate production in the colon. 31 Butyrate regulates inflammation, immunity, and oxidative balance and can function as histone deacetylase inhibitors (HDACi) to promote human health. 15 In addition, butyrate is an important energy source (providing 5%-15% of caloric requirements 30 ) and controls cell proliferation, differentiation, and apoptosis in colon cells (Figure 1).15,32,33
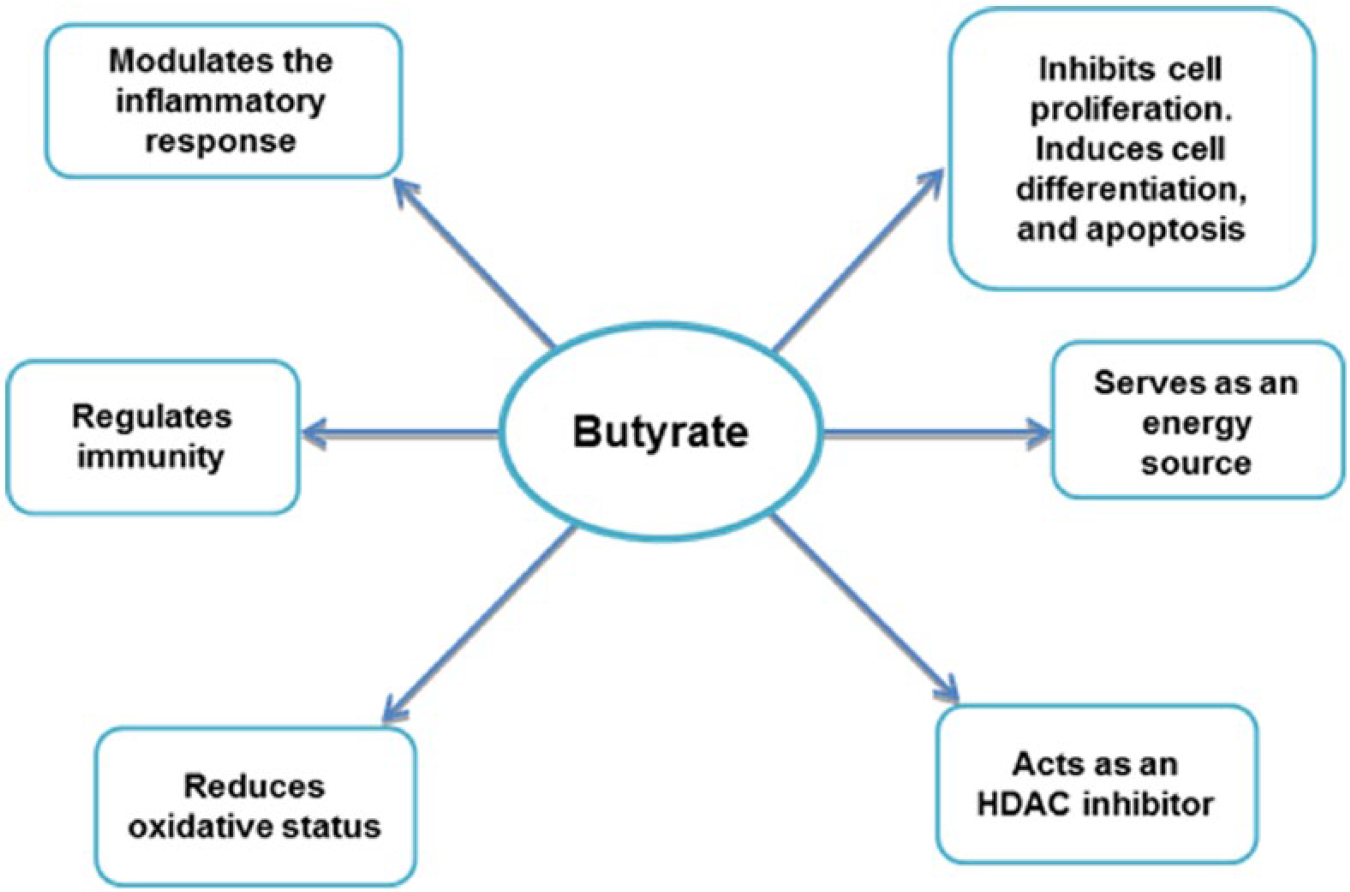
The effects of butyrate on colon cells. HDAC indicates histone deacetylase.
In both in vivo and in vitro studies, it has been reported that butyrate has bioactivity to reduce colon cancer risk. 22 Butyrate can modulate immune response in the colon and regulate the balance of gut bacteria to maintain colon homeostasis and reduce colon carcinogens (Figure 1). 34 In this way, butyrate can ameliorate the CRC risk induced by high consumption of red and processed meat, although there is some conflicting data in this regard.22,30,34,35
HDACI are used as chromatin-modifying drugs to treat cancers and other diseases. 36 Butyrate has the ability to inhibit HDAC activity and thus inhibit DNA damage and the activation of oncogenes.36–38 Table 1 shows a summary of effects of butyrate on colon and other cancer types.
Anti-cancer properties of butyrate through regulating miRNA and gene expression.

Abbreviations: ANXA1, lipocortin 1; DNMT 1, DNA (cytosine-5)-methyltransferase 1; HDACi, histone deacetylase inhibitors; MUC 2, mucin 2; NaB, sodium butyrate; NF-κB, nuclear factor κB; PCR, polymerase chain reaction; qRT-PCR, reverse-transcription quantitative PCR; RT-PCR, real-time PCR; TSA, trichostain A (histone hyperacetylating agent).
Butyrate can help maintain homeostasis and oxidative status in the gut and it can modulate immune response through gut microbiota and colonic epithelial cells. 21 One of the problems associated with experimentation with butyrate is that it is difficult to quantify butyrate in the human body because the absorption of butyrate occurs very soon after production, making it difficult to measure. 30
Evidence from animal trials and in vitro studies supports the anti-tumorigenic effects of butyrate.24,29,46 In an animal model study conducted by Shen et al, 47 HDACi upregulated selective miRNAs to modulate B-cell differentiation and the immune response of the host. In vivo and in vitro studies found that butyrate can also modulate the inflammatory response by the suppression of nuclear factor κB activation. 48 Many studies claimed that butyrate can reduce cancer risk via regulating cell proliferation, differentiation, and apoptosis.34,42,43 However, a combination of SCFAs as well as individual tested SCFAs (acetate, propionate, and butyrate) may damage the colonic mucosa of infant rats, in an age-dependent fashion. 49 This is thought to be a function of an immature immune system.
In in vitro studies, sodium butyrate (NaB) is often used but is generally referred to as butyrate. 36 The predominant effect of butyrate is the ability to induce apoptosis of cancer cells and inhibit DNA damage. Both animal and human trials demonstrated that butyrate is associated with inflammatory bowel disease (IBD) such that lower concentrations of butyrate are found in the faecal matter from patients with IBD relative to that found in people with a healthy gut. 30 According to animal and human trials, butyrate can help to attenuate the inflammatory profile of intestinal mucosa in patients with IBD, in particular, in patients with ulcerative colitis. 50 Patients with IBD are more susceptible to CRC, although the connection between IBD and CRC remains inconsistent. 51 Furthermore, butyrate can help suppress diet-induced obesity which is a risk factor for CRC. 52
Besides the ability of butyrate to inhibit HDAC activity, another anti-cancer mechanism of butyrate is the modulation of miRNAs in cancer cells. In several studies, it has been claimed that butyrate regulates both oncogenic miRNAs and cancer suppressor miRNA expression in various cancers, such as lung, breast, and colon cancers.45,53–56
MiRNA and Risk or Progression of CRC
MicroRNAs are small non-coding RNAs of approximately 22 nucleotides that are found in eukaryotic cells. 57 MicroRNAs can epigenetically regulate gene expression by post-transcriptionally regulating messenger RNA (mRNA) function which can inhibit or promote protein production. 58 MicroRNAs are commonly found in both normal and malignant cells and they can regulate at least 30% of human genes. 59 More than 940 miRNAs have been identified since they were first discovered in 1993. MicroRNAs play an important role in cellular proliferation, differentiation, and apoptosis 60 and therefore are clearly important in cancer inhibition and progression.
MicroRNA expression is deregulated in cancer processes including cancer initiation, progression, and metastasis.58,61 Therefore, it is possible that miRNA profiles could be used as biomarkers for cancer diagnosis and for the assessment of response to treatment. MicroRNAs may be more appropriate targets for treating cancers than DNA or mRNA because a single miRNA can regulate hundreds of mRNAs in specific biological pathways. 57 Also, it may be more practical to modify the expression of the approximately 200 miRNAs that have been found to be associated with various cancers than it is to modulate the activity of more than 16 000 mRNAs. 57
MicroRNAs associated with cancers can be crudely classified into 2 groups: tumour suppressor miRNAs and oncomirs or oncomiRNAs (tumour promoters) (Table 2). For example, miR-25 promotes cell migration and invasion in oesophageal squamous cell carcinoma, 62 whereas miR-143 can inhibit cell invasion and migration in CRC. 63 The overexpression of oncogenic miRNAs and the decreased expression of tumour suppressor miRNAs are often responsible for carcinogenesis. 59 Therefore, the inhibition of oncogenic miRNAs and the promotion of tumour suppressor miRNAs can provide the mechanism to prevent and treat cancers. Although there are some limitations associated with the notion of modulating miRNAs as an anti-cancer therapeutic strategy, some novel drugs and techniques have yielded positive results. 59
MiRNAs associated with the inhibition or promotion of genes associated with colorectal cancer.
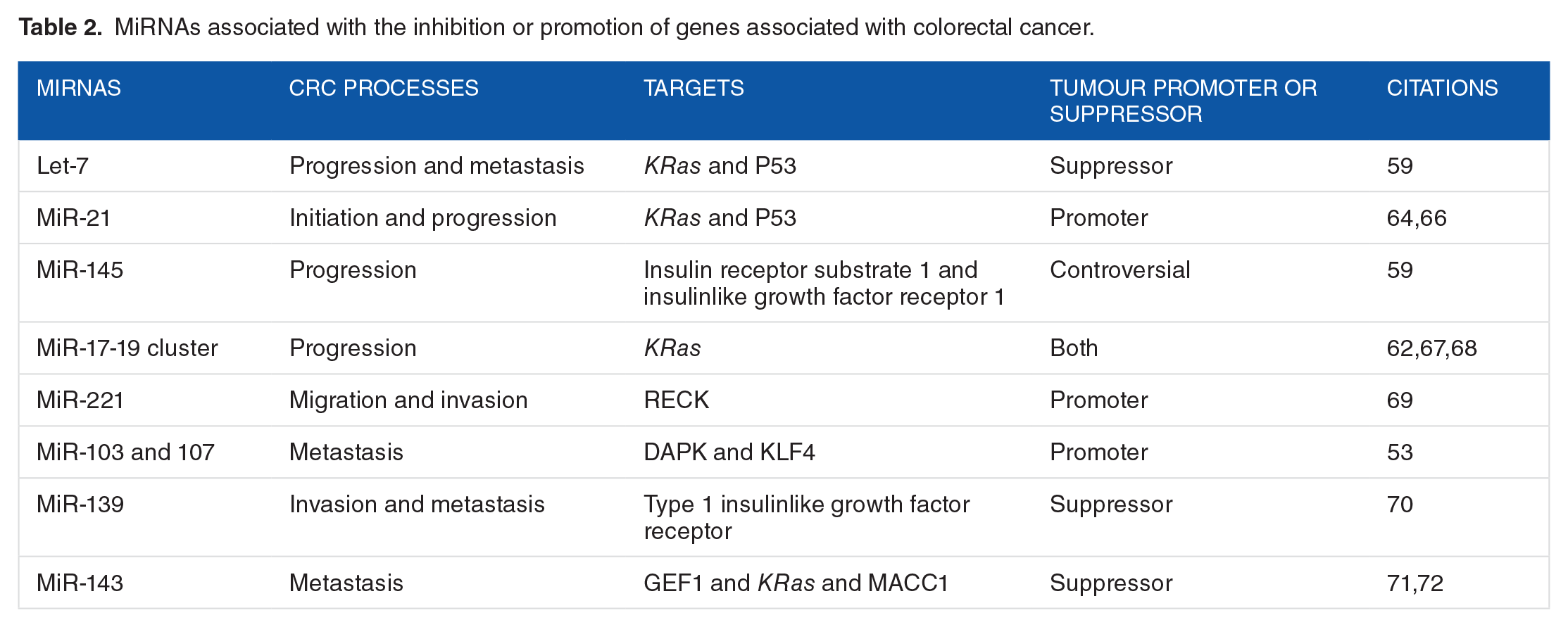
In CRC, miRNAs are involved in initiation, progression, and metastasis. There are several miRNAs commonly associated with colon cancers, such as Let-7, miR-21, miR-145, miR-17-19 cluster, miR-221, and miR-143, among others21,59,64 (Table 2). Moreover, Chiang et al 65 claimed that miR-192, miR-194, and miR-215 are associated with increased tumour size in colon cancer. It is essential to understand the role of miRNA expression in colon cancer to create effective preventive and therapeutic methods.
MicroRNA expression can be modulated by dietary nutrients.58,60 Generally, vitamins, minerals, and bioactive components in foods are considered to maintain people’s health and protect against some diseases. The effect of nutrients on regulating miRNA expression may help explain how these nutrients affect health. Hu et al 73 reported that butyrate can modulate miRNA expression in colon cancer such that the expressions of Let-7, miR-17-92a, miR-18-106a, and miR-25-106b clusters are decreased.
Effects of Butyrate on miRNA Expression in CRC Cells
In CRC cells, miRNA expression is largely associated with cancer initiation, progression, and metastasis. MicroRNA expression can be modulated by dietary factors including butyrate, and thus butyrate, as well as other dietary bioactives, could be used to create a less carcinogenic environment.
APC-β-catenin-TCF4 is an important signalling pathway in CRC initiation which regulates both cell proliferation and colonic cell differentiation. This pathway can be regulated by SCFAs. 74 In addition, Lazarova et al 75 investigated the effect of butyrate on the Wnt signalling pathway which is associated with the induction of apoptosis. They found that Wnt signalling–specific gene expression was modified by butyrate in the CRC HCT-116 cell line. 75 In turn, miRNAs can regulate the expression of the genes involved. 75 Therefore, there may be some association between butyrate and miRNA expression in CRC initiation.
In a study conducted by Hu et al, butyrate was also found to regulate miRNA expression in HCT-116 cells. 44 The miR-17-92a, miR-18b-106a, and miR-25-106b clusters were found to reduce the proliferation of HCT-116 cells relative to non-cancerous cells. 73 In addition, in a study conducted by Schlörmann et al, 76 it was reported that butyrate can upregulate the tumour suppressor gene P21, which is associated with the development of colon tumours. This occurs through impacting certain miRNAs. The effect of butyrate on miRNA expression in CRC cells is summarised in Table 3. According to the findings of studies listed in this table, the beneficial effect is to promote the expression of tumour suppressor and inhibit oncogenic miRNAs associated with CRC.
Evidence supporting the alteration of miRNA expression in CRC in response to butyrate.

Abbreviations: AOM, azoxymethane; HAMSB, butyrylated high-amylose maize starch; HRM, high red meat; LAMS, low-amylose maize starch.
In a human trial conducted by Humphreys et al, 28 a dietary intervention was used to determine the association between diet and miR-21. The dietary interventions included a high red meat (HRM) (300 g daily) intake versus a high intake (40 g daily) of HAMSB together with a HRM diet. This study was conducted in healthy human volunteers for 2 to 4 weeks, and the outcome showed that HRM may increase miR-21 in human rectal mucosa. 28 In this study, it was reported that HRM may increase CRC risk and the positive effects of butyrate may help suppress the harm induced by an HRM diet. Levels of miR-17-92 were restored when HRM was consumed with HAMSB, but the levels of miR-21 were not restored to baseline. 28
Outside of a clinical trial, the equivalent intake of 20 to 40 g of HAMSB/RS is feasible in the form of maize/corn, oats, barley, fruits, and vegetables but not through the consumption of wheat products.
Conclusions and Prospects
In conclusion, from the evidence presented, it is clear that butyrate is an HDACi that can modify histones which in turn can modulate the expression of miRNAs in colon and other cancers. This may provide novel ideas to prevent and treat CRC. Various miRNAs are sensitive to different concentrations of butyrate and to different treatment durations, and it is important that future work focuses on determining the physiological relevance. Butyrate may reduce CRC risk through decreasing the effect of exposure to carcinogens from foods. However, the ability of butyrate to negate the effect of regular and frequent red meat consumption remains unclear.
In addition, a number of unanswered questions are raised, particularly around exactly how butyrate may interact with a receptor or ligand to modify histones and regulate the expression of miRNAs. With an increase in understanding will come progress in the clinical application of butyrate perhaps as an anti-cancer prodrug and or in combination therapy with anti-inflammatory therapies.
Footnotes
Peer review:
Three peer Reviewers submitted review reports. The reports consisted of 864 words, excluding comments to the Editor.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: K.S.B. was supported by Auckland Cancer Society Research Centre and H.X. by University of Auckland Medical Foundation.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
KSB conceived and wrote the first draft of the manuscript. HX performed the review of the literature and prepared the figure. KSB, HX, and GM contributed to the writing of the manuscript and made critical revisions and approved final version. KSB and GM jointly developed the structure and arguments for the manuscript. All authors reviewed and approved the final manuscript.
Disclosures and Ethics
As a requirement of publication, author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including but not limited to the following: authorship and contributorship, conflicts of interest, privacy and confidentiality, and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest.