
Abstract
Caenorhabditis elegans is a powerful model to study the neural and biochemical basis of behavior. It combines a small, completely mapped nervous system, powerful genetic tools, and a transparent cuticle, allowing Ca++ imaging without the need for dissection. However, these approaches remain one step removed from direct pharmacological and physiological characterization of individual neurons. Much can still be learned by “getting under the hood” or breaching the cuticle and directly studying the neurons. For example, we recently combined electrophysiology, Ca++ imaging, and pharmacological analysis on partially dissected ASH nociceptors showing that serotonin (5-HT) potentiates depolarization by inhibiting Ca++ influx. This study challenges the tacit assumption that Ca++ transient amplitudes and depolarization strength are positively correlated and has validated a new paradigm for interpreting Ca++ signals. Bypassing the cuticle was critical for the success of these experiments, not only for performing electrical recordings but also for the acute and reversible application of drugs. By contrast, drug soaking or mutating genes can produce long-term effects and compensatory changes, potentially confounding interpretations significantly. Therefore, direct studies of the physiological response of individual neurons should remain a critical objective, to provide key molecular insights complementing global Ca++ imaging neural network studies.
The nematode Caenorhabditis elegans is a key model organism in neuroscience owing to its tiny and relatively simple nervous system (302 neurons), which is stereotyped in development and structure and has been fully reconstructed by serial section electron microscopy. 1 Additional advantages of C elegans include powerful genetic tools to manipulate gene expression globally, or in specific neurons, and remarkable conservation with mammals of most key molecular pathways regulating nervous system function.2,3 Importantly, C elegans is also transparent, allowing optical imaging in intact behaving animals. Ca++ imaging has been the predominant approach. Depolarization activates voltage-gated Ca++ channels, leading to an influx of Ca++ ions into the neuronal cytoplasm, which bind and activate fluorescent Ca++ reporter proteins. Ca++ reporters, such as the GCaMPs, have been highly optimized for sensitivity and speed and are likely to remain the tools of choice for the foreseeable future. Ca++ imaging has been used effectively to characterize sensory neuron function, 4 to elucidate the roles of novel ion channels in sensory neurons 5 and to identify circuit motifs coupling sensory input to locomotory output. 6 More recently, global patterns of neuronal activity have been identified through pan-neuronal expression of GCaMP markers and whole-animal imaging of freely behaving worms to elucidate circuit activity patterns underlying behavioral states and transitions. 7
Caenorhabditis elegans is also a powerful system to understand neuronal plasticity, brought about by neuromodulatory transmitters such as monoamines and neuropeptides. These transmitters influence neuronal excitability and synaptic vesicle release through GPCR (G protein–coupled receptor) signaling cascades, shifting circuit activity patterns, and ultimately modifying behavior in the face of changing conditions. 8 Neuromodulation is critical for human health, with disrupted neuromodulatory signaling causing myriad neurological disorders including major depression, schizophrenia, and drug addiction. 8 Ca++ imaging studies performed in C elegans have begun to produce important insights into neuromodulation in simple circuits, correlating behavioral changes with altered neuronal activity patterns and identifying the underlying transmitters and receptors. These studies have demonstrated subtle changes in Ca++ transient patterns, including altered intensities,9,10 oscillations, 11 and differential regulation of local and global Ca++ pools, 12 and have revealed interesting circuit motifs such as stimulus-driven cross-inhibition of sensory neurons 9 and neuropeptide-mediated negative feedback loops. 11
We have examined the modulation of aversive responses by serotonin (5-HT).13,14 Caenorhabditis elegans is repelled by 1-octanol, which is sensed by the 2 ASH polymodal nociceptors. Off food, worms respond to 1-octanol after a 10-second lag, but on food or 5-HT, this lag time shortens to 5 seconds, dependent on the SER-5 5-HT receptor functioning in the ASH neurons. 15 We therefore expected 5-HT to augment ASH Ca++ responses, downstream of SER-5 signaling, but surprisingly, 5-HT dramatically reduced ASH Ca++ signals. 13 To explain this paradox, we characterized the Ca++ signal in detail, demonstrating that it was primarily due to activation of the EGL-19 L-type Ca++ channel, but also comprised a significant contribution from IP3-dependent and ryanodine receptor–dependent release of Ca++ from intracellular stores. Most importantly, we used patch-clamp recordings to show that 5-HT potentiated depolarization and reduced the Ca++ signal under exactly the same conditions, demonstrating that changes in neuronal Ca++ do not necessarily predict parallel changes in their depolarization in C elegans. 13
To address the mechanistic basis of this unexpected result, we hypothesized that 5-HT might be acting downstream of depolarization to inhibit Ca++ entry. We exposed ASH to the bath through partial dissection, and depolarized the neuron by exposure to high [K+] external buffer. 14 This approach allowed us to step up the membrane potential independent of 1-octanol sensation to focus on events subsequent to generation of the initial sensory potential. Under these conditions, 5-HT still inhibited ASH Ca++ entry, dependent on the SER-5 receptor and Gαq signaling, as we previously observed for 5-HT modulation of 1-octanol responses, 14 suggesting 5-HT was blocking Ca++ entry downstream of depolarization. Previous work in cardiomyocytes and mammalian neurons identified a pathway for regulation of L-type voltage-gated calcium channels (VGCCs) known as Ca++-dependent inhibition (CDI). Ca++-dependent inhibition inactivates the channels by dephosphorylating a highly conserved serine residue (also present in EGL-19) and is dependent on release of intracellular Ca++ from IP3-mediated stores and the subsequent activation of calcineurin (CaN).16,17 Two observations suggest that a CDI-like pathway operates downstream of 5-HT/SER-5 in the ASHs. First, acute application of 5-HT leads to a spike in ASH Ca++ from an internal source, dependent on SER-5. Second, CaN (TAX-6 in C elegans) is necessary for 5-HT inhibition of the Ca++ response, as the 5-HT inhibition is markedly reduced in tax-6 mutants, or in the presence of cyclosporin A to pharmacologically inhibit TAX-6. Together, these results point to 5-HT inhibiting the Ca++-dependent readout of the depolarization through a CDI-like pathway (summarized in Figure 1). 14
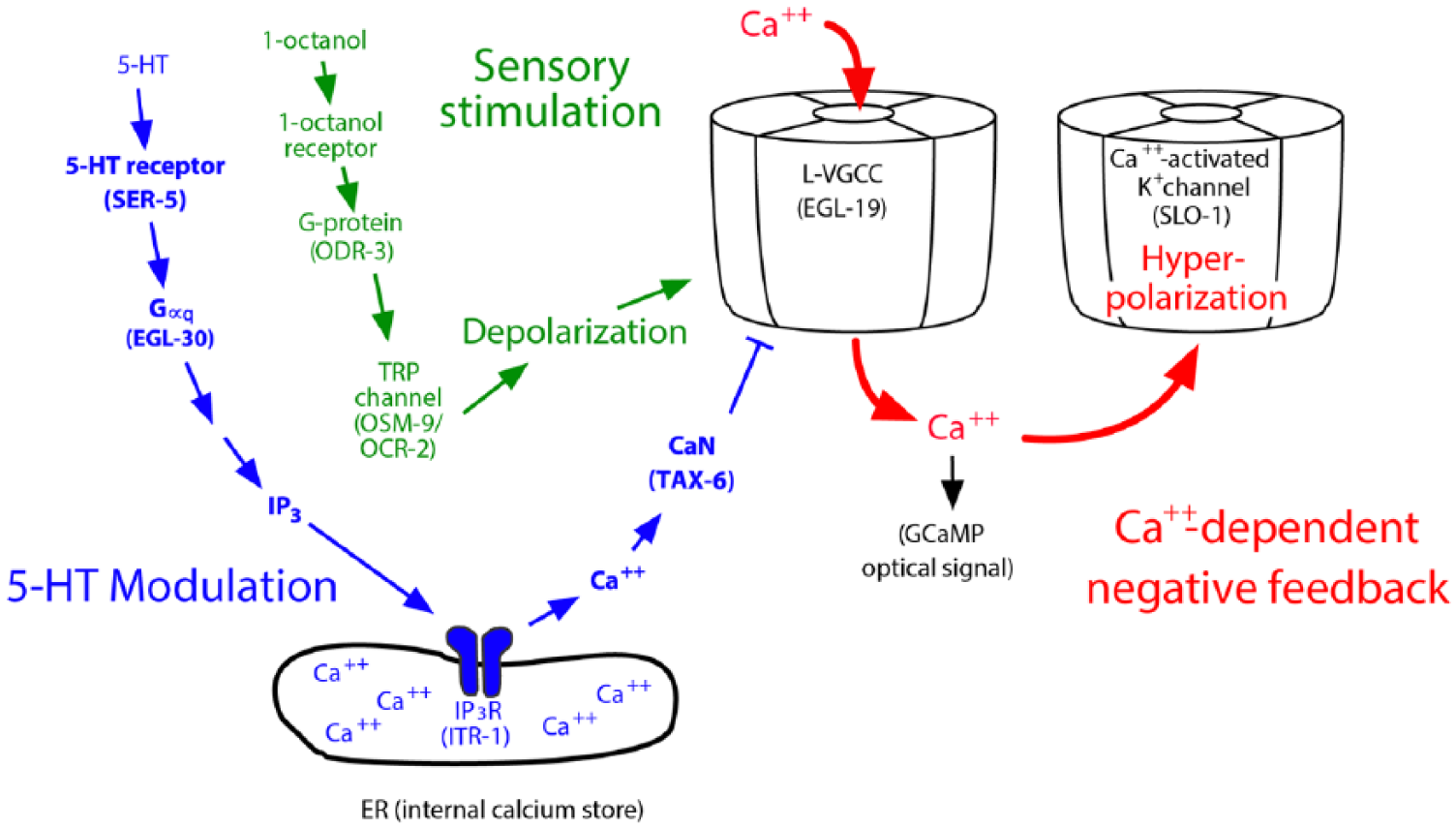
Serotonin (5-HT) disinhibits the ASH nociceptive neurons by modulating Ca++-dependent negative feedback. The aversive odorant 1-octanol activates an olfactory signal transduction pathway, which culminates in the activation of OSM-9/OCR-2 transient receptor potential (TRP) channels, and depolarizing inward currents (sensory stimulation, green). Depolarization activates EGL-19 L-type voltage-gated calcium channels (VGCCs), and the resulting Ca++ influx activates the SLO-1 Ca++-activated K+ channel, leading to hyperpolarizing outward currents (Ca++-dependent negative feedback, red). This Ca++ influx was also a major contributor to the 1-octanol–dependent Ca++ signals recorded in Ca++ imaging experiments.13,14 In parallel, 5-HT signaling, through the Gαq-coupled receptor SER-5, leads to the release of intracellular Ca++ (via the IP3 receptor), which activates the CaN orthologue TAX-6 to dephosphorylate and inhibit EGL-19 (5-HT modulation, blue), thereby decreasing 1-octanol–dependent Ca++ influx, reducing SLO-1–mediated negative feedback, and potentiating 1-octanol–evoked depolarization.13,14
This inverse correlation between Ca++ and depolarization amplitudes led us to hypothesize that Ca++ actually acts as a second messenger to inhibit depolarization. Using patch-clamp recording, we showed that the pharmacological inhibition of EGL-19 channels significantly increases the amplitude of depolarization stimulated by 1-octanol. 14 This observation explains the inverse correlation between Ca++ and depolarization amplitudes that we observed previously. Moreover, it precludes a role for voltage-gated Ca++ channels propagating the stimulus-dependent depolarization in the ASHs. Caenorhabditis elegans neurons do not have voltage-gated Na+ channels and rely on passive conductance instead of action potentials, which has led to speculation that VGCCs may amplify and propagate depolarization by gating cationic currents throughout the cell. However, we could not detect a compensatory increase in Ca++ influx in any part of ASH after inhibition of the EGL-19 L-type Ca++ channel that could play this role. 14 However, we did observe a strong persistent Ca++ signal in the cilia, which is likely due to activation of OSM-9/OCR-2 transient receptor potential channels as part of the sensory transduction cascade. 14 These channels conduct Na+ and Ca++ and can potentially provide sufficient driving force to depolarize the entire ASH neuron, given the high input resistances and large length constants of C elegans neurons, which allow local voltage changes to spread long distances passively, with minimal attenuation. 18
The Ca++-activated K+ channel SLO-1 is likely to be the downstream Ca++ effector in the 5-HT signaling pathway modulating 1-octanol responses in the ASHs. Loss-of-function mutation and ASH-specific RNAi knockdown of SLO-1 abrogate the 5-HT potentiation of 1-octanol response lag times. 14 L-type VGCCs and Ca++-activated K+ channels are well known to co-localize and function together to set and modulate membrane potential and electrical excitability in many other systems 19 and it appears they are playing this conserved function here as well. Interestingly, SLO-1 loss also disrupted the temporal regulation of ASH 1-octanol responses, leading to marked asynchrony between left and right ASH Ca++ transients and associated defects in aversive locomotory responses. 14 Together, these observations identify a point of convergence, at the L-VGCC, between a sensory response pathway and the 5-HT/SER-5–dependent modulatory pathway (see Figure 1), which work together to ensure highly reproducible sensory responses with physiologically appropriate amplitudes. It will be interesting to determine how other monoamine and neuropeptide signaling cascades within the ASHs interact with the 5-HT and Ca++-dependent pathways described above.
To gain these insights, it was essential to “get under the hood,” or in other words, bypass the barrier posed by the nematode cuticle and manipulate the neurons directly. Obviously, electrical recordings require direct physical contact of neuronal membranes with the patch pipette. However, direct solution contact with neurons was also critical because it allowed acute exposure of neurons to solutions of known concentration, over precisely defined intervals. By comparison, in traditional drug soaking approaches, solutes diffuse across the cuticle slowly (if at all), resulting in gradually ramping temporal concentration gradients, with specific internal concentrations unknowable at any given time point. The advantages of acute direct application uniquely allowed us to define the physiological concentration range of 5-HT, to demonstrate internal stores Ca++ release as part of the 5-HT/SER-5 signaling cascade, and to assay the immediate effects (within 60s) of EGL-19 L-VGCC blockade on depolarization amplitudes. Underscoring the importance of that short time frame, we observed that physiological adaptation occurred within 30 minutes of EGL-19 L-VGCC inhibition, which was severe enough to make experiments uninterpretable; similar effects may potentially occur over developmental time frames when key neuronal genes are mutated or knocked down using RNAi. Looking forward, acute application of ligands in partially dissected preparations can be a powerful approach to verify molecular relationships identified in genetic, behavioral, and Ca++ imaging studies. For example, in neuropeptide signaling, single peptides can be applied individually to detect specific effects on putative target neurons (ie, neuropeptide genes typically encode multiple distinct peptides with different targets and activities). 20 Monoamines can be applied at precisely defined concentrations, to address the relationships between multiple receptors for the same ligand within individual neurons. Ca++ imaging, electrical recording, and optical measurement of synaptic vesicle release, coupled with sensory stimulation and other modes of controlled depolarization may all be useful approaches to characterize the physiological changes brought about by these modulators and assign their roles in circuit modulation with the greatest possible accuracy.
Footnotes
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work is supported by National Institute of Allergy and Infectious Diseases.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
All authors contributed to the conception, writing, and editing of this article.