
Abstract
Background:
We previously showed that cocoa, a rich source of polyphenols improved the age-associated health and extended the lifespan in C. elegans when supplemented starting from L1 stage.
Aim:
In this study, we aimed to find out the effects of timing of cocoa exposure on longevity improving effects and the mechanisms and pathways involved in lifespan extension in C. elegans.
Methods:
The standard E. coli OP50 diet of wild type C. elegans was supplemented with cocoa powder starting from different larval stages (L1, L2, L3, and L4) till the death, from L1 to adult day 1 and from adult day 1 till the death. For mechanistic studies, different mutant strains of C. elegans were supplemented with cocoa starting from L1 stage till the death. Survival curves were plotted, and mean lifespan was reported.
Results:
Cocoa exposure starting from L1 stage till the death and till adult day 1 significantly extended the lifespan of worms. However, cocoa supplementation at other larval stages as well as at adulthood could not extend the lifespan, instead the lifespan was significantly reduced. Cocoa could not extend the lifespan of daf-16, daf-2, sir-2.1, and clk-1 mutants.
Conclusion:
Early-start supplementation is essential for cocoa-mediated lifespan extension which is dependent on insulin/IGF-1 signaling pathway and mitochondrial respiration.
Introduction
As a result of combined effects of demographic and epidemiological transitions and “modernization” over the past few decades, well-recognized non-communicable diseases (NCDs) such as cardiovascular diseases, cancers, diabetes and neurodegenerative diseases have become the leading public health challenges globally. 1 Aging has been identified as the main risk factor for these prevalent NCDs. 2 A growing list of genetical, behavioural and pharmacological interventions that have shown to improve longevity also have proven effective in delaying the onset of age-related diseases and preserving healthspan. 3 The well-defined pathways that regulate longevity including insulin/insulin-like growth factor-1 signalling (IIS), mitochondrial respiration, calorie restriction (CR) are also found to be involved in the onset of aging-related diseases.4-8 Therefore, the therapies that modulate these pathways may be beneficial in delaying/preventing the onset of age-related diseases.
In recent years, dietary interventions have been proposed as alternative approaches to drugs for the prevention of aging-related diseases, particularly the consumption of foods rich in polyphenols. 9 Polyphenols have been shown to alleviate age-associated phenomena such as oxidative stress, chronic inflammation and toxin accumulation. 10 Resveratrol, curcumin, quercetin, catechin are some of the polyphenols well-studied for their antioxidant activity as potent compounds to mitigate the age-associated oxidative stress and damage induced by metabolic production of reactive oxygen species (ROS).11-14 Cocoa, one of the widely consumed foods is known to be rich in polyphenols comparatively in much higher levels of total phenolics and flavonoids than tea and wine. 15 Moreover, the cocoa-based product chocolate has been identified as a significant contributor to the total antioxidant capacity of European and American diets. 16
Previously, we reported the effects of long-term cocoa supplementation (starting from the first larval stage till die) at varying doses (1, 2, 3, 4, 5 mg/ml) on lifespan of Caenorhabditis elegans (C. elegans), with 3 mg/ml dose achieving the maximum mean lifespan extension. 17 In addition, the same dose extended the median lifespan as well, but not the maximum lifespan. Based on these findings, in this study, we wanted to investigate the influence of timing of cocoa exposure on the longevity improving effects with the dose which previously reported the maximum lifespan extension. Additionally, we determined the pathways and mechanisms involved in the lifespan extension effect of cocoa.
Materials and Methods
Strains and culture conditions
All C. elegans strains and Escherichia coli (E. coli) OP50 were acquired from Caenorhabditis Genetics Center (CGC) and C. elegans were maintained at appropriate temperature (15°C or 20°C) as per instructions. C. elegans strains used in this study were as follows: N2 (Bristol, wild-type), DA1116 eat-2 (ad1116) II., VC199 sir-2.1 (ok434) IV., CB4876 clk-1 (e2519) III., EU1 skn-1 (zu67) IV., CB1370 daf-2 (e1370) III., ZG31 hif-1 (ia4) V., TJ356 daf-16(zls356) IV., and GR1307 daf-16 (mgDf50) I. All strains were maintained on nematode growth medium (NGM) plates seeded with E. coli OP50.
Cocoa treatment
Specifications and the composition of cocoa powder used in this study as well as the preparation of E. coli OP50 food source, and the addition of cocoa to NGM plates were described previously. 17 The major bioactive phytochemicals in cocoa are catechins including mainly monomeric (-) epicatechin and (+) catechin as well as oligomeric and polymeric procyanidins. 18 The total phenolics in cocoa powder used for the study were 27.01 mg GAE/g which was about 56% compared to natural cocoa powder. Total flavonoids were 10.13 mg CE/g which was about 40% compared to the levels in natural cocoa powder. For all the experiments, 3 mg/ml cocoa dose was used. Influence of timing on lifespan extending effects was determined by exposing N2 worms to cocoa at different larval stages (L1, L2, L3, and L4) till they die, L1 to adult day 1 and from adult day 1 till worms die (Figure 1A and B). C. elegans comprises of 4 larval stages (L1, L2, L3, and L4) which are followed by adulthood. The larva emerges from the eggshell has the nervous system and musculature but lacks the reproduction ability. Over four larval stages which ends with a molt, gonad and reproductive system are formed finally making the sexually mature adult worm. The larva is known to grows roughly continuously in size throughout these 4 stages with little change in overall morphology. The larval stages are mainly discriminated by their size. The L1 larvae is about 250 μm in length while the L2, L3, L4, and adult are 360 to 380 μm, 490 to 510 μm, 620 to 650 μm, and 1110 to 1150 μm respectively. 19 We supplemented worm diet with cocoa at different larval stages as well as adulthood to represent the different stages of growth and development.
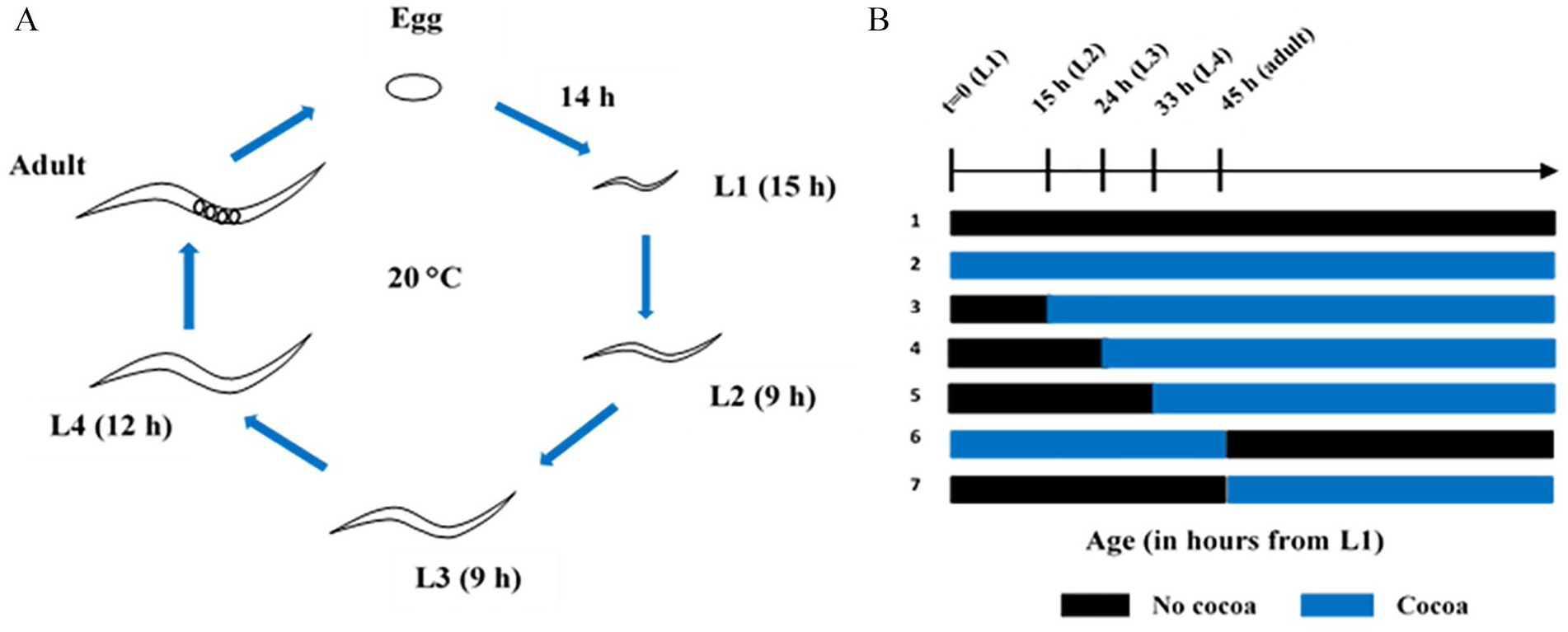
Cocoa-supplementation at different growth stages of C. elegans: (A) lifespan of wild-type C. elegans at 20°C and (B) schematic representation of cocoa-supplementation at different stages of life (treatment 1 = control; treatment 2 = cocoa from L1 stage; treatment 3 = cocoa from L2 stage; treatment 4 = cocoa from L3 stage; treatment 5 = cocoa from L4 stage; treatment 6 = cocoa from L1-adult day 1; treatment 7 = cocoa from adult day 1 till die).
To determine the pathways and mechanisms involved, mutant strains were supplemented with cocoa at a dose of 3 mg/ml starting from L1 stage till the death.
Obtaining synchronous cultures
Age-synchronized worms were obtained by bleaching gravid adults. Briefly, gravid hermaphrodites were washed off from an NGM plate with M9 buffer and made the final volume to 3.5 ml. Bleaching solution (bleach: 5N NaOH = 2:1) was added to the worm suspension at a volume of 1.5 ml. The suspension was mixed by vortexing the tube 5 seconds and keeping further 30 seconds on rest until all the worm bodies got dissolved. The suspension was centrifuged (2 min at 1300g) to pellet the released eggs and the supernatant was poured off. M9 buffer was added to a final volume of 5 ml, centrifuged again and the supernatant was removed. This step was repeated at least 3 times to completely get rid of the bleaching solution. The egg pellet was resuspended in 3 ml of M9 and kept for 48 hours on a shaker for hatching.
Lifespan assay
Lifespan assay was performed as described in Sutphin and Kaeberlein. 20 Floxuridine (FUdR) was not used to avoid progeny production and instead worms were transferred everyday onto fresh plates until they stop laying eggs (until day 9). Thereafter, worms were transferred every other day. Worms that were crawled off from the plates were excluded from the analysis. Survival curves were plotted. Mean and median lifespan were reported.
DAF-16::GFP localization assay
Control and cocoa-supplemented (3 mg/ml) TJ356 daf-16(zls356) IV. worms were scored at L4 stage for cytoplasmic, intermediate or nuclear Green fluorescent protein (GFP) localization as described by Wang et al. 21 About 100 control and cocoa-supplemented (from L1 stage) worms were collected at L4 stage, washed with M9 buffer twice and placed on a glass slide carrying 2 µl of 1 M NaN3. The sub-cellular DAF-16::GFP distribution was observed under a fluorescence microscope at 20-fold magnification. The number of worms in above mentioned three categories were counted and expressed as percentages. Three independent experiments were performed.
Statistical analysis
All statistical analyses were performed in IBM SPSS® statistics software (version 24). Data were expressed as mean ± standard error of mean (SEM). Survival function was estimated using Kaplan-Meier curves. Survival curves were compared using log rank (Mantel-Cox) test. The differences between groups for DAF-16 localization was determined using t-Test (two-sample assuming equal variances).
Results
Cocoa extends the lifespan of wild-type C. elegans when supplemented starting from L1 stage
We supplemented the worm diet with cocoa at different larval stages (L1, L2, L3, and L4) till the death, from L1 till adult day 1 and from adult day 1 till the death. Worms showed a significantly extended mean lifespan when supplemented with cocoa starting from L1 stage till they die (P < .05, 8.9% increase, Figure 2A, Table 1). Similarly, cocoa extended the lifespan of worms when supplemented at L1 till only day 1 of adulthood (P < .05, 6.7% increase, Figure 2A, Table 1). However, supplementation of cocoa at L2, L3, or L4 stages could not increase the mean lifespan of worms (Figure 2A, Table 1). Cocoa supplementation at adult day 1 till the death (treatment 7) also could not increase (P < .05) the mean lifespan of worms (Figure 2B, −3.7%, control = 16.2 ± 0.20, and cocoa = 15.6 ± 0.19 days).

Survival curves for control and cocoa-supplemented wild type (N2) C. elegans. Experiments were performed in triplicate. Differences between groups for mean lifespan were calculated using log rank (Mantel-Cox) test. (A) Survival curves for worms supplemented with cocoa at different larval stages (L1, L2, L3, and L4) till die and from L1 to adult day 1. Cocoa significantly increased (P < .05) the mean lifespan when worms were supplemented starting from L1 stage. Cocoa could not extend the lifespan of worms when supplemented at other larval stages (L2, L3, and L4). (B) Survival curves for cocoa-supplemented worms from adult day 1 till die [n = 335 for control and n = 336 for cocoa-supplemented worms (treatment 7)]. Cocoa could not extend the mean lifespan of worms when supplemented from adulthood.
Mean and median lifespans of C. elegans when supplemented with cocoa at different stages of life.

Statistically significant (P < .05) compared to the control group.
Cocoa extends the lifespan of C. elegans via DAF-16
DAF-16, the C. elegans homolog of the forkhead box transcription factors class O (FoxO) is a central regulator of aging, development, stress, metabolism, and immunity.
22
Therefore, we tested if DAF-16 can play a role in lifespan extension by cocoa in C. elegans. We found that cocoa could not extend the lifespan (P > .05) of daf

Cocoa-induced nuclear DAF-16 localization in C. elegans: (A) representative images of TJ356 C. elegans for cytosolic, intermediate and nuclear DAF-16::GFP localization (20-fold magnification, scale bar = 50 μm) and (B) percentage of control and cocoa-treated worms grouped for cytosolic, intermediate and nuclear DAF-16::GFP localization. About 100 worms (at L4 stage) per treatment were used for each replicate. Bar chart represents mean ± SE of 3 independent experiments. Statistical significance was determined using t-Test (two-sample assuming equal variances). Cocoa-treated worms showed a higher nuclear DAF-16 localization compared to control worms (P < .05).
Mean and median lifespan of control and cocoa-supplemented C. elegans.

Statistically significant extension compared to the control group (P < .05).
Cocoa may regulate insulin/insulin-like growth factor-1 signalling (IIS) pathway
In C. elegans, IIS pathway is known to regulate longevity via DAF-16. 23 Therefore, we explored whether cocoa could interact with molecules in IIS pathway to regulate the longevity. Single mutations of DAF-2 inhibit IIS pathway and increases longevity in C. elegans. 24 Therefore, we used daf-2 (e1370) III. mutants to study the involvement of IIS pathway in cocoa-mediated lifespan extension. Cocoa-supplementation could not extend the mean lifespan of long-lived insulin-like receptor mutant daf-2 (Figure 4B, Table 2).

Different pathways and mechanisms involved in cocoa-mediated lifespan extension. Experiments were performed in triplicate. Differences between groups for mean and median lifespans were calculated using log rank (Mantel-Cox) test. (A) Survival curves for control and cocoa-supplemented GR1307 daf-16 (mgDf50) I. worms. Cocoa could not extend the lifespan of daf-16 mutants (P > .05). (B) Survival curves for control and cocoa-supplemented CB1370 daf-2 (e1370) III. Worms. Cocoa could not extend the lifespan of daf-2 mutants. (C) Survival curves for control and cocoa-supplemented VC199 sir-2.1 (ok434) IV. worms. Cocoa could not extend the lifespan of sir-2.1 mutants. (D) Survival curves for control and cocoa-supplemented DA1116 eat-2 (ad1116) II. worms. Cocoa could not extend the lifespan of eat-2 mutants. (E) Survival curves for control and cocoa-supplemented EU1 skn-1 (zu67) IV. worms. Cocoa significantly extended the lifespan of skn-1 mutants (P < .05). (F) Survival curves for control and cocoa-supplemented ZG31 hif-1 (ia4) V. worms. Cocoa significantly extended the lifespan of hif-1 mutants (P < .05). (G) Survival curves for control and cocoa-supplemented CB4876 clk-1 (e2519) III. worms. Cocoa could not extend the lifespan of clk-1 mutants.
Cocoa could not extend the lifespan of sir-2.1 and eat-2 mutants
The silent information regulator 2 (SIR2) can bind to DAF-16 and extend the lifespan of C. elegans. 25 We investigated if cocoa could act on SIR-2.1 to extend the lifespan of C. elegans. Cocoa could not extend the mean lifespan of sir-2.1 mutant, indicating that SIR-2.1 is required for cocoa-mediated lifespan extension (Figure 4C, Table 2). In addition, as SIR-2.1 is considered as a key mediator of lifespan extension by CR, 26 we used the pharyngeal pumping defective mutant eat-2 (ad1116) II. to see if cocoa-induced lifespan extension is dependent on a CR mechanism. According to our results, cocoa could not extend the mean lifespan of eat-2 mutants (Figure 4D, Table 2).
Lifespan extension by cocoa was not dependent on stress response factors
In C. elegans skn-1 promotes resistance to oxidative stress and extends lifespan.
27
In this study, cocoa-supplementation significantly increased the mean lifespan of skn-1 mutants (P < .05, Figure 4E, Table 2). Additionally, we determined the effects of cocoa
Cocoa restores the mitochondrial function to extend the lifespan of C. elegans
The role of mitochondrial function in aging is well-described.29-31 Therefore, we used electron transportation chain (ETC) mutant clk-1 which shows an extended lifespan compared to wild type C. elegans 32 to see the involvement of mitochondria in cocoa-induced lifespan extension. Cocoa supplementation could not extend the mean lifespan of clk-1 mutants (Figure 4G, Table 2).
Discussion
In humans, early life nutrition has been reported to significantly influence the risk of developing NCDs in late life, 33 emphasizing the importance of early nutrition on healthspan. Earlier initiation of nutritional interventions in humans has been shown to confer greater benefits in long-term cardiometabolic outputs. 34 Moreover, studies suggest that early life nutrition can affect intestinal maturation and gut health in later life. 35 Enhancing the nutrition at early life is known to regulate the gut microbiota composition and improve the infant immunity development, shaping the life-long health. 36 Maternal intake of resveratrol, a polyphenol in Red wine has been shown to fight against the adverse effects of the high-fat diet or low-protein diet on offspring, such as glucose intolerance, obesity, cholesterol metabolic disorders, non-alcoholic fatty liver disease, or even hypothalamic leptin signaling dysregulation in mice. 37
We previously reported that long-term cocoa supplementation extended the lifespan of C. elegans when supplemented starting from L1 stage. 17 Based on aforementioned findings on early-life nutrition on health of individuals in later life, we wanted to investigate whether early exposure to cocoa is critical for this longevity improving effect of cocoa.
Our results supported the idea that cocoa supplementation starting from first larval stage (L1) is critical and essential for the longevity extending effects of cocoa. In addition, cocoa intervention starting from L1 stage which continued till day 1 of adulthood increased the lifespan of worms to a similar extent as life-long exposure (L1 to the death). Therefore, these results suggest that the longevity extending effects of cocoa require early-start exposure, but do not need to be continued in a long-term manner. Previous studies support our findings where supplementation of a cranberry extract rich in polyphenols has been prominently promoted the longevity in C. elegans when the supplementation was started at early developmental stage compared to the late-start. 38 Moreover, Zuckerman and Geist 39 reported that the antioxidant α-tocopherol supplementation at early in the pre-reproductive stage significantly extended both mean and maximum lifespan in C. elegans. These effects arose from early supplementation were prominent compared to the supplementation at day 4, indicating α-tocopherol was most effective if present during larval stages.
However, cocoa supplementation at late larval stages (L2, L3, L4) and adulthood did not seem to play a role in lifespan extension. Additionally, supplementation of cocoa at these stages significantly reduced the lifespan. Some of the previous studies have been reported the adverse effects of polyphenols and antioxidant supplementation in animal models. Resveratrol, one of the most researched polyphenols has been reported to act as a pro-oxidant at high concentrations, promoting DNA damage while increasing oxidative stress in vitro and in animal models. 40 Supplementation of the growth medium with alpha-tocopherol increased oxidative stress and decreased cell lifespan in Saccharomyces cerevisiae, highlighting the pro-oxidant action of antioxidants. 41 Another study reported that dietary supplementation of both vitamin E and C dramatically shortened the lifespan in voles with no exact explanation for the effect on lifespan but may be due to pro-oxidant effects. 42 In humans, supplementation of antioxidants (vitamin C and E) has been reported to suppress the health-promoting effects (insulin sensitivity, reactive oxygen species defense) of physical exercise. 43 In addition, the consumption of the green tea derived polyphenol (-)-epigallocatechin gallate or its metabolites has been reported to associate with hepatotoxicity in humans. 44 However, in this study, we were not able to establish the exact mechanism for lifespan reduction effects of cocoa when supplemented at late larval stages and adulthood.
Molecular mechanisms underlying aging have recently gained much attention as aging is the most significant risk factor for many chronic disease conditions.
45
Most of these mechanisms influencing aging are conserved across organisms.
46
Among different genetic factors that regulate aging, IIS, mitochondrial metabolism and CR pathways have been extensively studied.
47
To find out which pathways are involved in cocoa-associated lifespan extension in C. elegans, we used several mutant strains related to these well-known pathways. IIS pathway, the first pathway implicated in the aging of animals has a well-established role in aging where the reduced IIS leads to lifespan extension in C. elegans.
48
IIS pathway is a signal transduction cascades that consists of insulin-like peptides (ILPs), an insulin/IGF-1 receptor (DAF-2), a phosphoinositide 3-kinase (AGE-1/AAP-1/PI3K), serine/threonine kinases (PDK
Mitochondrial respiration is another significant contributor to the aging process.
52
Impairments in ETC is known to extend the lifespan of C. elegans.
53
We tested the effects of cocoa supplementation on long-lived clk-1 mutants. This gene encodes a hydroxylase in the ETC that is required for ubiquinone biosynthesis.
54
We found that clk
Conclusion
In summary, we demonstrated that early-start supplementation is essential for cocoa-mediated lifespan extension in C. elegans. In addition, longevity improving effects of cocoa were mediated through IIS pathway and mitochondrial respiration.
Footnotes
Acknowledgements
We would like to thank Dr. Ebony Monson and A/Prof. Karla Helbig from Helbig lab, Department of Physiology, Anatomy and Microbiology, School of Life Sciences, La Trobe University, Bundoora, Australia for the help with the fluorescence microscope for GFP images.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by La Trobe University Postgraduate Research Scholarship (LTUPRS) and La Trobe University Full Fee Research Scholarship (LTUFFRS).
Declaration of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Study conception and design: MM and MJ; conducting experiments and data collection: MM; data analysis and drafting the manuscript: MM; reviewing of the manuscript; MJ, AA, JT and DH.