
Abstract
Recent decades have seen much progress in the identification and characterization of cannabinoid receptors and the elucidation of the mechanisms by which derivatives of the Cannabis sativa plant bind to receptors and produce their physiological and psychological effects. The information generated in this process has enabled better understanding of the fundamental physiological and psychological processes controlled by the central and peripheral nervous systems and has fostered the development of natural and synthetic cannabinoids as therapeutic agents. A negative aspect of this decades-long effort is the proliferation of clandestinely synthesized analogs as recreational street drugs with dangerous effects. Currently, the interactions of cannabinoids with their biochemical substrates are extensively but inadequately understood, and the clinical application of derived and synthetic receptor ligands remains quite limited. The wide anatomical distribution and functional complexity of the cannabinoid system continue to indicate potential for both therapeutic and side effects, which offers challenges and opportunities for medicinal chemists involved in drug discovery and development.
Keywords
Introduction
Cannabis sativa is one of society’s oldest cultivated plants, with records of its propagation and use dating back thousands of years. Due to widespread consumption for medicinal and recreational purposes, the manner in which the chemical constituents in cannabis produce their pharmacological effects has been the subject of considerable interest and investigation. Initially, the production of crude but concentrated forms, such as hashish and tinctures, showed that the pharmacologically active chemical constituents could be isolated and studied. As a result, the molecular structures of the unique compounds in cannabis (the phytocannabinoids) and their associated pharmacological activities were further defined. These efforts culminated in the structural elucidation of the principal psychoactive constituent, Δ9-THC (Figure 1), and facilitated the discovery of extremely potent synthetic cannabimimetic compounds, identification of their specific receptors and mechanisms of action, and further elucidation of the underlying biochemical systems through which cannabinoids exert their pharmacological actions. This article is an overview of the current understanding of the biomolecular basis for the pharmacological effects of cannabis and Δ9-THC–like cannabimimetics in humans. Emphasis is on (1) the types of cannabinoid receptors and their anatomical distribution; (2) the endogenous lipid signaling molecules that interact with these receptors (the endocannabinoids); (3) the enzymes and cellular processes involved in the synthesis, liberation, and degradation of the primary endocannabinoids N-arachidonoylethanolamine (anandamide) and 2-arachidonoylglycerol; and (4) the role of the endocannabinoid system in the modulation of cellular signaling processes and neuronal excitability in areas of the brain effecting central nervous system function and behavior.

Δ9-THC, the principal psychoactive constituent in Cannabis sativa.
Cannabinoid Receptors
Although interest in the pharmacology of cannabis dates back several millennia, unequivocal evidence supporting the existence of specific cannabinoid receptors in the central nervous system (CNS) of animals began to accumulate only recently. In the 1980s, extremely potent synthetic compounds (Figure 2) with cannabimimetic activity in laboratory animals were shown to have rigorous structure-activity relationships, including stereoselectivity.1–4 Further indication of the interaction of cannabinoids with specific receptor proteins was revealed through their ability to dose-dependently inhibit adenylate cyclase in a pertussis toxin–sensitive fashion that correlated with their pharmacological potency in laboratory animals.5–7 Radiolabeling of the potent synthetic cannabinoid CP-55,940 provided proof of a saturable, high-affinity stereoselective biomolecular site in rat brain whose binding affinity for cannabinoids correlated to both the inhibition of adenylate cyclase in vitro and analgesic activity in vivo. 8 Autoradiography studies subsequently revealed the anatomical distribution of these high-affinity binding sites in the CNS9–13 and peripheral tissues. 14 The pattern and density of distribution of the binding sites for cannabinoids in the brains of rats, dogs, monkeys, and humans were exceptional, with extremely high concentrations in the basal ganglia, hippocampus, cerebral cortex, and cerebellum (Figure 3). The widespread distribution of binding sites in the CNS correlates with the diverse effects of cannabinoids observed in laboratory animals and humans, including alterations in movement (eg, locomotor activity, coordination, and catalepsy), feeding and satiety (“the munchies”), learning, memory, and affective states. The unique pattern of distribution facilitated the cloning, sequencing, and identification of a 7-transmembrane-spanning G protein–coupled cannabinoid receptor (GPCR) in the brain (CB1) that was shown to be responsible for many of the diverse pharmacological actions of cannabinoids.15–17 A second receptor (CB2) was subsequently cloned and sequenced and found to be expressed in macrophages in the marginal zone of spleen and in other cells with immunological function. 18 While other receptors are substrates for certain cannabinoids, CB1 and CB2 are most closely associated with the pharmacological actions of Δ9-THC and other psychoactive cannabinoids in humans.
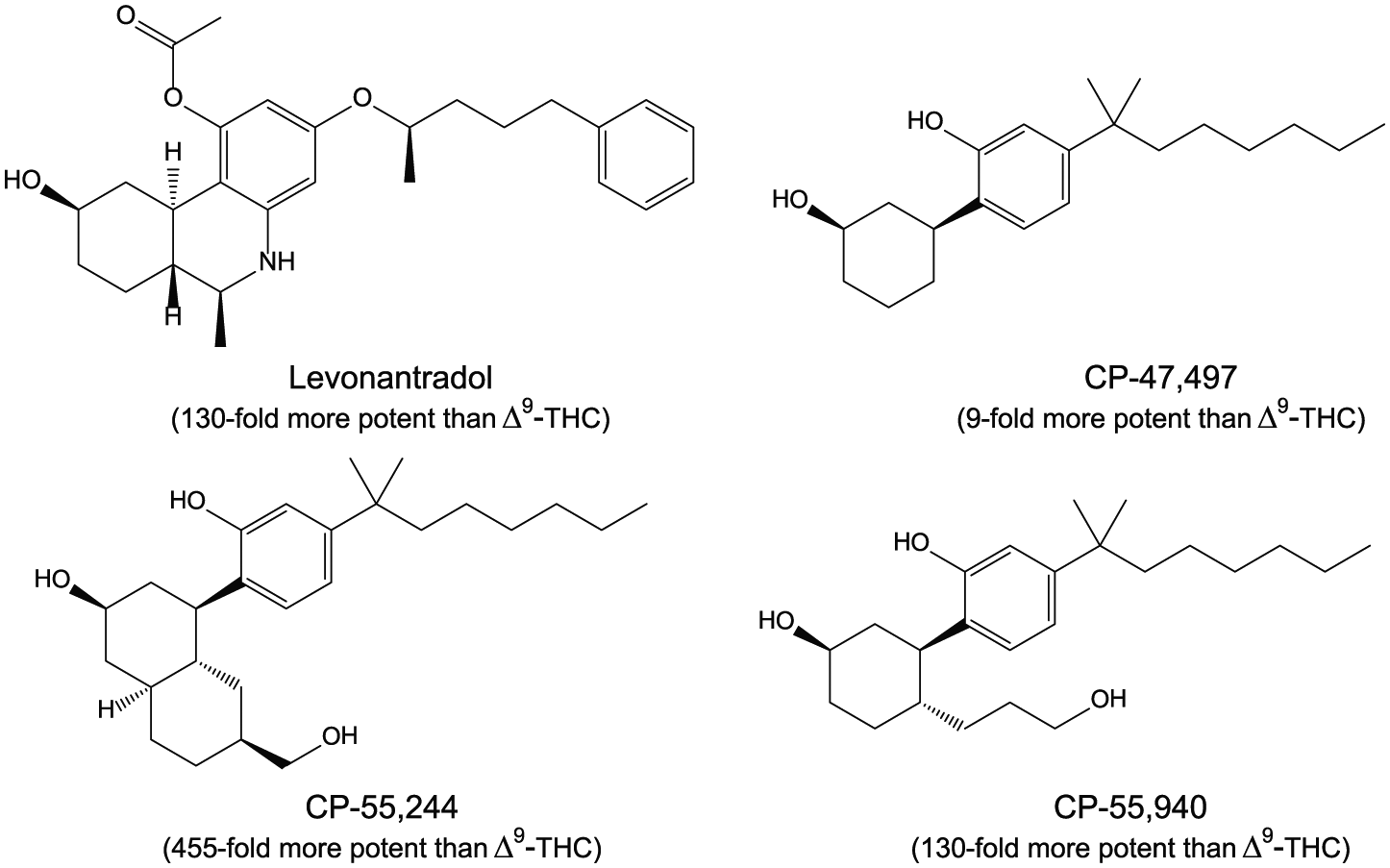
Four potent synthetic cannabinoids.

Radioligand autoradiograph of specific binding of [3H]CP-55,940 to a sagittal section of rat brain, showing high CB1 receptor densities in the hippocampus, basal ganglia, and cerebellum.
The CB1 and CB2 subtypes couple preferentially to G proteins of the Gi and Go classes.7,19–21 The GPCR signal transduction pathway serves to amplify intracellular processes in response to the formation of the ligand-bound receptor complex. On binding of a cannabinoid ligand, the conformational equilibrium of the GPCR is altered. A cannabinoid agonist alters the receptor conformations into those that activate the trimeric G protein, causing Gα to bind a molecule of guanine triphosphate (GTP) and dissociate from the other two G protein subunits. The activated G protein subunits disengage from the receptor and modulate signal amplification pathways involving several downstream effector proteins, including inhibition of specific adenylate cyclases and voltage-dependent calcium channels, and activation of inwardly rectifying potassium channels and several mitogen-activated protein kinases (with some variation depending on cell type, levels of receptor expression, and endogenous tone or constitutive activity; see Figure 4). Cannabinoid ligands can vary in their intrinsic activity or efficacy in producing conformational changes that result in a subsequent receptor-mediated response, with full agonists producing a maximal cellular response, partial agonists an intermediate response, antagonists no response (but reducing or blocking the effect of another ligand), and inverse agonists decreasing the constitutive activity of the receptor signaling complex, that is, driving the basal level of responsiveness negative, which is opposite to the effect of agonists. For example, Δ9-THC is a partial agonist with respect to its ability to stimulate GTP binding through the CB1 receptor compared with CP-55,940, which is a full agonist.22–24 This may help explain why smoking “Spice” and other synthetic cannabinoid–containing herbal formulations causes more severe panic attacks and other adverse effects than smoking cannabis.

Cannabinoid receptor signaling and trafficking. Modified and reproduced with permission from Al-Hasani and Bruchas. 25
The discovery of the CB1-selective receptor inverse agonist SR141716A (rimonabant) (Figure 5)26,27 permitted the effects of Δ9-THC and opposing endogenous cannabinoid function to be further illuminated. For example, this compound was shown to produce physical withdrawal signs in rats made tolerant to the effects of Δ9-THC. 28 Many of the central effects produced by Δ9-THC, such as increased food consumption and impaired learning and memory, were attenuated by SR141716A,29,30 providing further evidence of mediation through CB1. Furthermore, treatment with SR141716A alone enhanced spatial memory, 31 decreased food consumption, 32 and produced other pharmacological effects indicative of its ability to act as an inverse agonist or reverse existing endocannabinoid tone at the CB1 receptor. SR141716A also blocked the discriminative stimulus effects of Δ9-THC in rodents and reduced the reinforcing effects and reinstatement of palatable food33–35 and other drugs of abuse in animal self-administration models, including heroin, 36 cocaine, 37 nicotine,34,38,39 and ethanol.33,40 The CB2-selective inverse agonist SR144528 (Figure 5) was identified by the same research group at Sanofi 41 and shown to be a useful probe for cannabinoid-mediated effects in immune function. 42
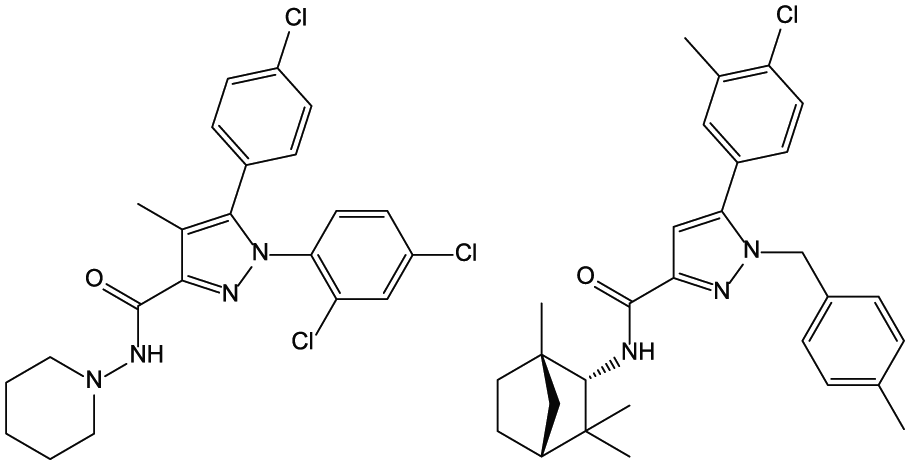
The CB1- and the CB2-selective inverse agonists SR141716A (left) and SR144528 (right).
Cannabinoid receptors also contain allosteric sites that bind small molecules that alter receptor structure and function—that is, allosteric modulators. Allosteric sites are topographically distinct from those bound by orthosteric agonists (eg, Δ9-THC and CP-55,940) and antagonists (eg, SR141716A and SR144528). When allosteric modulators bind to the allosteric site, they alter the conformational equilibrium of the receptor, which affects the ability of the receptor to bind compounds at the orthosteric site and couple to signal transduction molecules. Thus, allosteric modulators may either positively or negatively modulate the affinity or efficacy of orthosteric agonists and antagonists. Both positive and negative allosteric modulators (Figure 6) of cannabinoid receptors produce pharmacological effects distinct from those produced by orthosteric ligands.43–51 Theoretically, allosteric modulators possess no basal activity in the absence of orthosteric ligands. Because of this, allosteric modulators might provide more selective spatial and temporal signaling, 52 exerting their effects only in the presence of orthosteric ligands such as endocannabinoids, which are transiently released on demand and removed from their sites of action by enzymatic hydrolysis or cellular uptake. Thus, this fine-tuning of endogenous ligand signaling through orthosteric binding sites, instead of directly activating or blocking the receptor, may provide a promising alternative strategy for treating cannabinoid receptor–mediated disorders. However, as some compounds might bind to the allosteric site and act alone to alter the receptor’s inherent coupling to signaling molecules (constitutive activity and signaling bias), it is also possible for compounds to act as allosteric agonists and allosteric inverse agonists, exerting their effects in the absence of an orthosteric ligand.53,54 Interestingly, there are phytocannabinoids in cannabis, 47 as well as endogenous chemical constituents found in humans,49,55,56 that have been reported to produce allosteric modulation of cannabinoid receptor function.

CB1 allosteric modulators. Org 27569 (left) and PSNCBAM-1 (right) display positive binding cooperativity with the orthosteric agonist CP-55,940 for CB1 receptor binding as would positive allosteric modulators, but reduce the efficacy of orthosteric agonists in several in vitro functional assays.
Endogenous Cannabinoid Signaling Molecules (Endocannabinoids)
The sequencing and cloning of the cannabinoid receptors and the availability of high-affinity radiolabeled ligands facilitated the screening and identification of endocannabinoids that act as cannabinoid receptor modulators. The first endocannabinoid, anandamide (Figure 7), was isolated and identified in the porcine brain.57,58 It was shown to have an affinity (Ki) of approximately 50 nM for the CB1 receptor 57 and a much lower affinity of 1600 nM for CB2. 19 Another endocannabinoid lipid, 2-arachidonoylglycerol (2-AG), was isolated and identified in canine gut and shown to bind to both CB1 and CB2 with Ki values of approximately 500 and 1400 nM, respectively. 59 Like Δ9-THC, both compounds inhibit adenylate cyclase in isolated mouse spleen cells and electrically evoked contractions of the mouse vas deferens and produce the typical tetrad of pharmacological effects after intravenous administration in mice: antinociception, immobility, reduction in spontaneous activity, and reduction in rectal temperature.57,59–61 2-Arachidonoylglycerol is generally considered to be an orthosteric full agonist and anandamide an orthosteric partial agonist at both cannabinoid receptor subtypes in most tissues and signal transduction assays.62,63 Many other endogenous and synthetic lipids have since been shown to interact with cannabinoid receptors.

The endocannabinoids arachidonoylethanolamine (anandamide, left) and 2-arachidonoylglycerol (2-AG, right).
Anandamide is derived from the precursor N-arachidonoyl phosphatidylethanolamine, which is synthesized by the relatively nonselective enzyme N-acyltransferase.64,65 In the brain, anandamide and other N-acylethanolamines are released “on demand” in an activity-dependent manner by enzymatic cleavage of lipid precursors by phospholipase D 66 and other distinct biochemical pathways.67–69 When released from the membrane, these lipids are typically bound reversibly to carrier or transport proteins called fatty acid–binding proteins. 70 N-acylethanolamines distributed in this way can act as diverse cellular signaling molecules in neural and nonneural tissues. Anandamide and other fatty acid amides are primarily degraded through the actions of fatty acid amide hydrolases (FAAHs).71–73 Inhibitors of FAAH activity lead to increased levels of anandamide and other fatty acid amides and prostamides in tissues and biological fluids. Anandamide is also a substrate for metabolism through cyclooxygenase 2 (COX-2), resulting in the production of bioactive prostamides. Inhibition of FAAH and/or COX-2 increases endocannabinoid tone and is recognized as having therapeutic potential distinct from that available with exogenous receptor agonists of cannabinoid receptors. 74 FAAH (−/−) knockout mice have elevated levels of anandamide in the brain and demonstrate analgesia that is reversed by the CB1 antagonist SR141716A. 75 Moreover, several FAAH inhibitors have analgesic and anti-inflammatory activity in animal models76,77 and have been tested in clinical trials for a variety of therapeutic end points.78,79 However, there is heightened awareness of the risks for significant adverse events with this relatively indirect approach to cannabinoid therapeutics after a fatality, and severe neurologic disorders were encountered in clinical trials of the FAAH inhibitor BIA 10-2474. 80 Although this compound is suspected to have a prolonged and broad spectrum of activity at hydrolases compared with more selective FAAH inhibitors (such as PF-3845 by Pfizer and JNJ-42165279 by Janssen), 81 the underlying biochemical mechanisms involved in the toxic cerebral syndrome remain largely unknown. 82
2-Arachidonoylglycerol is also an integral signaling component of lipid membranes but is produced, liberated, and degraded in a manner different from that of anandamide. In neuronal tissues, the synthesis of 2-AG begins with the precursor phosphatidylinositol 4,5-bisphosphate and proceeds via a diacylglycerol (DAG) intermediate that is cleaved to 2-AG by one of 2 diacylglycerol lipases, DAGLα or DAGLβ.83,84 It appears likely that the DAG intermediate of 2-AG is synthesized on demand following
Depolarization-induced activation of voltage-gated Ca2+ channels, wherein the increased intracellular Ca2+ concentration facilitates cleavage of phosphotidylcholine (PC) by phospholipase D to phosphatidic acid (PA), and subsequent removal of the phosphate group via PA phosphatase, to yield DAG;
Activation of Gq/11-coupled receptors, such as metabotropic glutamate receptors and muscarinic acetylcholine receptors, coupled to phospholipase C (PLCβ)-mediated generation of DAG;
Increase in DAG through activation of PLCγ-coupled tyrosine kinase–linked growth factor receptors;
Most of the DAG in the brain is enzymatically degraded by monoacylglycerol lipase (MAGL), and even modest systematic inhibition of MAGL can profoundly enhance 2-AG concentrations and effects. Acute inhibition of MAGL activity potentiates CB1 receptor signaling 87 and produces agonist-like effects in laboratory animals. 88 However, both chronic pharmacological inhibition 89 and genetic deletion 90 of MAGL produce functional reductions in CB1 receptor signaling (ie, β-arrestin 2–mediated desensitization) rather than activation.86,91 The alpha/beta domain–containing hydrolase 6, alpha/beta domain–containing hydrolase 12, COX-2, and FAAH enzymes can also participate in the degradation of 2-AG.92,93
The Multifaceted Functioning of the Endocannabinoid System
The neuronal endocannabinoid system involves the concerted actions of endocannabinoids, cannabinoid receptors, and the enzymes responsible for the synthesis and degradation of endocannabinoids. In the CNS, most CB1 receptors function to modulate neurotransmitter release and are present in particularly high concentrations in preterminal axon segments and axon terminals while being sparsely distributed in the active zone of the synapse 94 (Figure 8). The enzymes involved in endocannabinoid biosynthesis and degradation are also heterogeneously distributed throughout the brain in a manner consistent with their function. 95 In the hippocampus, for example, the DAGLα isoform is concentrated on dendritic spine heads of pyramidal cells and produces 2-AG that participates in retrograde synaptic signaling with presynaptic CB1 receptors located on afferent terminals, which serves to adjust neurotransmitter release as a function of postsynaptic activity. 96 Monoacylglycerol lipase is found in laminar and punctate staining patterns on the afferent axon terminals instead of the postsynaptic structures at the ultrastructural level, demonstrating the post- and presynaptic segregation of the primary enzymes responsible for synthesis and elimination of 2-AG, respectively, in the human hippocampus. The anatomical arrangement is consistent with the endocannabinoid-mediated retrograde regulation of neuronal activity and synaptic plasticity in the cortex, hippocampus, amygdala, and dorsal striatum involved in executive function, perception, integration, learning, memory, extinction, recall, and habit. 86 Inhibition of synaptic transmission by CB1 agonists has been demonstrated in diverse regions of the central and peripheral nervous systems, involving glutamatergic, GABAergic, noradrenergic, cholinergic, and other neurotransmitters. 97 For instance, the cannabinoid receptor also functions in the peripheral (although still neuronal) and central (spinal and supraspinal) levels in the pain pathway. Even at the subcortical level where CB1 density is relatively sparse, the endocannabinoid system regulates the release of hypothalamic orexigenic and anorexigenic signals and modulates activity in mesolimbic dopaminergic incentive pathways and opioidergic hedonic circuits controlling motivation for and perceived value of rewards such as palatable food. 98 The CB2 receptor is expressed to a much lower extent in neurons in the mammalian CNS. In particular, CB2 messenger RNA and receptor protein have been found in several brain stem regions, including the dorsal motor nucleus of the vagus, the nucleus ambiguus, and the spinal trigeminal nucleus. Although CB2 signaling has putative roles in addiction and behavior, its impact in CNS functions and pathologies is less certain than that of the CB1 system. 99

Neuronal endocannabinoid system. Reproduced with permission from Lu and Mackie. 100
Components of the endocannabinoid system are also distributed throughout the periphery and in several nonneuronal tissues, where they participate in a range of physiological and pathological processes. For instance, endocannabinoid signaling in the gastrointestinal tract, adipocytes, liver, kidney, skeletal muscle cells, and pancreas contributes to the control of intestinal motility, nutrient intake, energy storage, metabolism, and elimination processes involved in homeostasis.101,102 In the male and female reproductive systems, endocannabinoid signaling functions along with hormones in the control of fertility. 103 Cannabinoid receptors are also found on several immune cell types and within organs (eg, spleen and thymus) with immune system function14,104 that can produce and degrade endocannabinoids. Endocannabinoid tone and exogenous cannabinoid agents such as Δ9-THC modulate cytokine (eg, tumor necrosis factor-α, interleukin-1β, interleukin-6, interleukin-12, and interleukin-10) and chemokine production, the expression of adhesion molecules, and the migration, proliferation, and apoptosis of cells.105,106 Thus, the endocannabinoid system also provides an integral component in the regulation of the cell cycle, immune system function, and inflammation. For example, the expression of CB2 is increased in activated astrocytes, reactive microglia, oligodendrocytes, perivascular microglia, and neural progenitor cells, making the CB2 receptor a particularly interesting biochemical target for the development of therapeutics for cancer, neuroinflammatory and neurodegenerative diseases, ischemic trauma, and stroke. 107
Conclusions and Future Directions
Many advances in our understanding of the receptors, biochemicals, and enzyme systems composing the endocannabinoid system have been made in the past few decades. Because of the widespread distribution and diverse functioning of the system components, the effects observed during its modulation or dysregulation can be numerous and varied. The neuromodulatory role of the system and its actions in the control of energy balance (at the level of both food intake/satiety and peripheral storage and metabolism) are two of the most well-characterized effects observed in humans. Individuals increasing their endogenous cannabinoid tone with cannabis or cannabinoids experience altered sensations, perceptions, cognition, learning, memory, and behavior. The pharmacological effects of either herbal cannabis or Δ9-THC vary in intensity with dose and route of administration and are modulated to some degree by the modest affinity and efficacy of the partial agonist Δ9-THC. The subjective effects are often perceived as pleasant or euphoric, but increased appetite and antiemetic effects are the only clinically approved therapeutic indications for synthetically derived Δ9-THC (as dronabinol, trade name Marinol) in the United States, and it remains a scheduled substance because of its abuse liability. Consumption of illicit herbal products (eg, “Spice”) containing synthetic cannabimimetic alkylindoles with high affinity and efficacy at CB1 and CB2 receptors can produce extreme alteration in the subjective effects observed in humans, leading to panic, confusion, anxiety, and other undesirable effects quite dissimilar from Δ9-THC. Chronic repeated exposure to cannabinoid agonists can alter receptor signal transduction and trafficking pathways (eg, receptor internalization and downregulation/desensitization) in a process of adaptation that can lead to tolerance, dependence, and withdrawal upon abstinence. CB1 receptor expression levels and availability in homeostatic and mesolimbic reward regions are inversely related to body mass index across individuals with and without food intake disorders. 98 Obese individuals decreasing their endogenous cannabinoid tone with the human-use formulation of the inverse agonist SR141716A (rimonabant) tended to lose weight and have improved biological markers of diabetes mellitus.108–111 However, the drug was removed from the market because of treatment-associated depression and suicidal ideation that could be associated with its inverse agonist activity in the mesolimbic reward system.98,109,110,112,113
It should be emphasized that the integration and function of the various components of the endocannabinoid system are complex and that modulation influences diverse physiological processes that continue to be identified and further characterized. As a result, our understanding of drug use, addiction, obesity, and other pathological conditions for which endocannabinoid-based therapeutics might be targeted continues to increase dramatically. Moreover, because of the multifaceted functioning of the system, opportunities for advances in therapeutic agents that directly or indirectly affect CB1 and CB2 receptors (eg, medicinal cannabis and synthetic Δ9-THC) are unlikely to provide clinical advantage in the absence of unwanted side effects. Nevertheless, it should also be recognized that the continued investigation of the endocannabinoid system provides a knowledge-based foundation for cannabis use/abuse and the development of more selective, effective, safe therapeutic agents. For example, compounds that selectively target CB1 but do not cross the blood-brain barrier could eliminate CNS effects, such as abuse liability, yet retain peripheral effects on energy storage and utilization. Therefore, further investigation and development of endocannabinoid system modulators appear to be well -warranted and promising.
Footnotes
Acknowledgements
Dr Gerald T. Pollard assisted in the critical review and editing of this manuscript.
Peer review:
Four peer reviewers contributed to the peer review report. Reviewers’ reports totaled 699 words, excluding any confidential comments to the academic editor.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by the National Institute on Drug Abuse DA-040460.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
BFT conceived and wrote the brief review.
Disclosures and Ethics
As a requirement of publication author(s) have provided to the publisher signed confirmation of compliance with legal and ethical obligations including, but not limited to, the following: authorship and contributorship, conflicts of interest, privacy and confidentiality and (where applicable) protection of human and animal research subjects. The authors have read and confirmed their agreement with the ICMJE authorship and conflict of interest criteria. The authors have also confirmed that this article is unique and not under consideration or published in any other publication, and that they have permission from rights holders to reproduce any copyrighted material. Any disclosures are made in this section. The external blind peer reviewers report no conflicts of interest.