
Abstract
The bacterium Oceanobacillus jordanicus strain GSFE11 is a halotolerant endophyte isolated from sterilized roots of Durum wheat (Triticum turgidum ssp. Durum) growing in hot and arid environments of Ghor Safi area in the Jordan Valley. The draft genome sequence and annotation of this plant growth-promoting endophytic bacterium are reported in this study. The draft genome sequence of Oceanobacillus jordanicus strain GSFE11 has 3 839 208 bp with a G + C content of 39.09%. A total of 3893 protein-coding genes and 68 RNA coding genes were predicted. Several putative genes that are involved in secretion and delivery systems, transport, adhesion, motility, membrane proteins, plant cell wall modification, and detoxification were identified, some are characteristics of endophytes lifestyle including genes that are involved in metabolism of carbohydrate, genes for xylose, fructose and chitin utilization, quinone cofactors biosynthesis, genes associated with nitrogen, sulfur, phosphate and iron acquisition, in addition to genes involved in the biosynthesis of plant hormone auxin. This study highlights the importance of using genome analysis and phylogenomic analysis to resolve the differences between closely related species, such analysis showed Oceanobacillus jordanicus strain GSFE11 to be a new species closely related to Oceanobacillus picturae (genome size 3.67 Mb), Oceanobacillus jordanicus has higher a number of predicted genes compared with Oceanobacillus picturae (3961 genes vs 3823 genes).
Introduction
Jordan Valley stretches from the Sea of Galilee to the Dead Sea; this valley has the lowest elevation on Earth, with less than 440 m below sea level. The Ghor area east of the Dead Sea has been cultivated for millennia, but in recent years the area has suffered from the scarcity of rainwater and an increment in soil salinity. 1 The abiotic stresses from drought, salinity, and extreme temperatures can limit plant growth and crop development. 2 Nowadays, the increase in soil salinity is a major worldwide concern, affecting areas like river deltas and many other semi-arid regions. 3 Salinity can have an effect on cellular metabolism, leading to several physiological, morphological, biochemical, and molecular changes in plants. 4 Salinity can impose osmotic stress which effect water availability and might contribute to the accumulation of toxic ions in the cells, oxidative stress damage, and nutrient imbalances. 5 These factors can impact plant growth and development.
Nonetheless, many plants seem to have adapted to living in such inhospitable environments. 6 Endophytic bacteria reside within plants, usually promoting plant growth by several mechanisms; they stimulate uptake of nutrients, usually by producing siderophores that can sequester iron, they participate in phosphorus solubilization and nitrogen fixation, 7 they can also produce certain bioactive secondary metabolites such as phytohormones, 8 and they might play a role in keeping the plant healthy by fighting other pathogens such as bacteria and fungi. 9
In a recent study, several rhizobacteria associated with durum wheat (Triticum turgidum ssp. durum) that have plant growth-promoting properties were isolated from fields of high salinity in Ghor area near the Dead Sea. 10 The isolated rhizobacteria strains possessed several plant growth-promoting traits including siderophore production, inorganic phosphate solubilization, nitrogen fixation, auxin production, and antifungal activity against Fusarium culmorum. Among the isolated halotolerant bacterial endophytes, GSFE11 was identified as Oceanobacillus sp. by sequencing the 16S ribosomal RNA (rRNA) gene. Interestingly, the endophytic strain GSFE11 has similar 16S rRNA sequence to GSFR11 a rhizospheric strain isolated from the soil surrounding the same plant indicating the occurrence of the strain in environmental and plant samples. 10 The endophytic strain GSFE11 was initially described as a gram-positive and rod-shaped bacteria forming small white colonies that were able to grow on saline media (15% NaCl). The strain was characterized for several plant growth-promoting traits and was found to produce Indole acetic acid (IAA) and degrade aminocyclopropane-1-carboxylic acid (ACC) and improved substantially the growth of durum wheat plants under salinity and drought conditions. Furthermore, GSFE11 showed the strongest inhibitory effect against antifungal activity against Fusarium. culmorum restricting its growth in a dual culture plate assay when compared to other strains. Oceanobacillus bacteria belong to the phylum Firmicutes, within the family Bacillaceae. The genus includes several species that are motile Gram-positive rods, growing obligatory aerobically or facultative anaerobically. Some species are moderately halophilic bacteria as GSFE11 strain is. Oceanobacillus bacteria were isolated from various environmental samples, including salt fields, 11 deep-sea sediment cores, 12 several food samples as in the traditional Korean fermented food (kimchi), 13 fermented shrimp samples, 14 equipment used in the production of soy sauce. 15 Oceanobacillus picturae was isolated from salt field 11 and human stool sample. 16 Oceanobacillus endoradicis, an endophytic bacterial species, was isolated from the root of Paris polyphylla, 17 and in another study, O. picturae (CML15) and O. picturae (CrL11) were isolated from Cakile maritima leaves. 18 Oceanobacillus has been shown to promote plant growth in several plants.11,19,20 As of this study, no whole genome of an endophytic Oceanobacillus is available yet.
Here, we report the first draft genome of an endophytic Oceanobacillus species. The draft genome of O. jordanicus strain GSFE11 has been assembled. Here, the details of the draft genome sequence are described.
Materials and Methods
Genomic DNA isolation, library preparation, and sequencing
From a single isolate colony, a 5 ml culture was grown in liquid NA media, this was used for total genomic DNA isolation using the Wizard® Genomic DNA Purification Kit (Promega Madison, WI, USA) following the manufacturer’s instructions. The isolated gDNA quality was assessed by gel electrophoresis in a 1% agarose gel stained with Red Safe (Intron, Bio-tek, Korea) visualized by using Gel Doc™ XR Gel Documentation System (Bio-Rad, Hercules, CA, USA). The concentration and quality of the gDNA were determined by using a spectrophotometer (Smart-Spec™ plus spectrophotometer, BIO-RAD, Hercules, CA, USA). The gDNA was sent to the service provider Macrogen Inc. (Seoul-Korea) to be sequenced using Illumina NovaSeq® platform, library type: TruSeq Nano DNA (350), type of reads: paired-ends, read length: 151 bp.
Pre-processing, genome assembly, and annotation
To assess the quality of the raw reads, FastQC v0.11.9 21 was used. Fastp 22 was used to trim the adaptor sequences and remove bad reads (too low quality, too short, or too many N). The sequence reads were de novo assembled into 36 contigs using SKESA v2.4.0 23 using the default parameters. The resulting 36-scaffolds draft genome sequence was submitted to NCBI. The draft genome was annotated using Rapid Annotations using Subsystems Technology (RASTtk) implemented in both PATRIC server 24 and in The Department of Energy Systems Biology Knowledgebase (KBase) serever 25 using the default settings.
Bioinformatics
Nine most closely related species with available published genomes (Oceanobacillus picturae Heshi-B3, Oceanobacillus picturae DMTMMB24, Oceanobacillus iheyensis HTE831, Oceanobacillus caeni DSM-21910, Virgibacillus halodenitrificans PDDB-F2, Oceanobacillus oncorhynchi, Oceanobacillus massiliensis, Oceanobacillus limi IBRC-M, Oceanobacillus kimchii-X50, ) were chosen to construct several phylogenetic trees as follow: a phylogenomic analysis was carried out via KBase serever, 25 utilizing the species tree builder functionality,, and OrthoANI (Orthologous Average Nucleotide Identity) utilizing the Orthologous Average Nucleotide Identity Tool (OAT) software. 26 In addition, a genomic signature distance phylogenetic tree was constructed via the GScompare server (http://gscompare.ehu.eus/) based on octanucleotides frequencies. A digital DNA-DNA hybridization was performed via Genome-to-Genome Distance Calculator 3.0 (GGDC) 27 implemented in TYGS server (https://tygs.dsmz.de/), a phylogenetic tree was inferred with FastME 2.1.6.1 28 from Genome Blast Distance Phylogeny (GBDP) distances calculated from genome sequences. The branch lengths are scaled in terms of GBDP distance formula d5. The numbers above branches are GBDP pseudo-bootstrap support values >60 % from 100 replications, with an average branch support of 80.6%. The tree was rooted at the midpoint. 29 The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) were predicted by CRISPRCasFinder online program. 30 Pangenomic analysis was conducted using KBase Compute Pangenome and Pangenome Circle Plot function. 25 Circular graphical display of the distribution of the genome annotations was carried out via PATRIC server Comprehensive Genome Analysis function. 31
Accessing the genome sequence
The data from this draft genome project (the raw sequences and the assembled draft genome of 36 scaffolds) have been deposited at GenBank with BioProject ID PRJNA753452 and BioSample ID SAMN20694754.
Data Set Interpretation
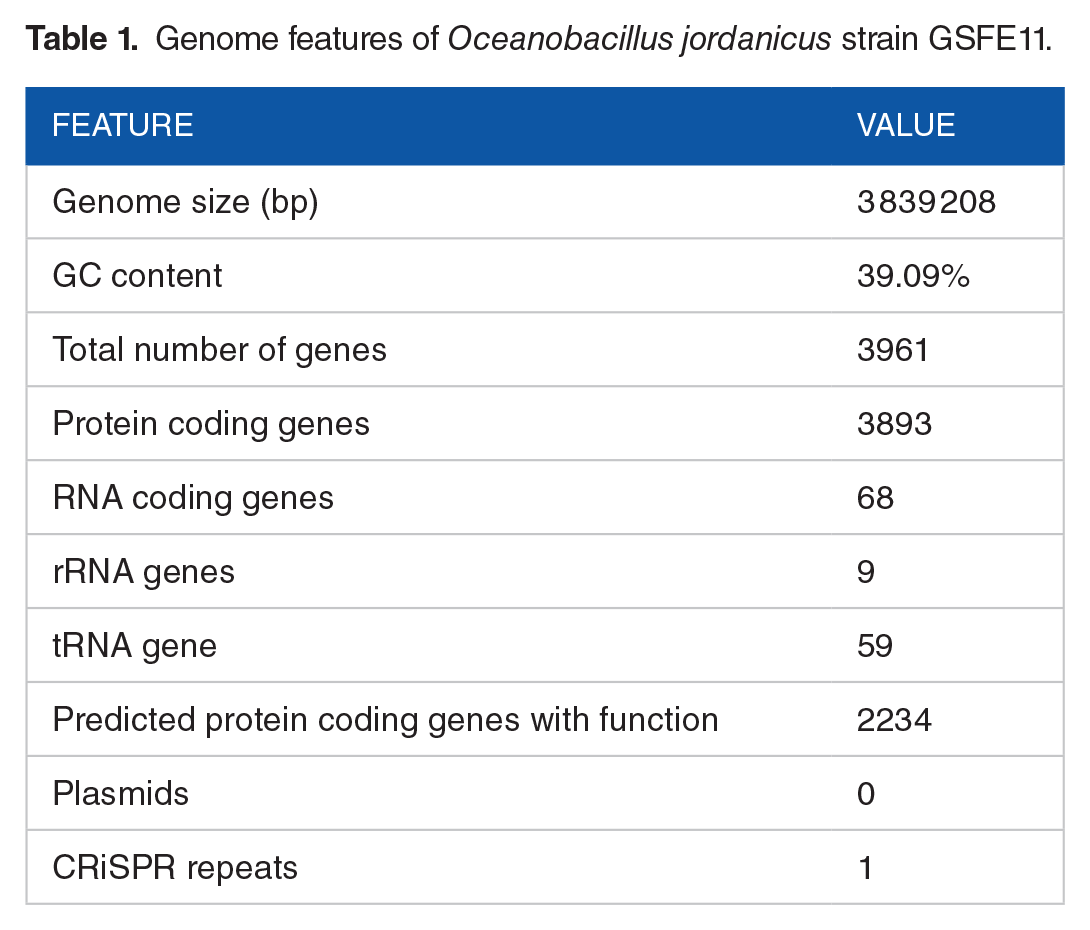
The draft genome of Oceanobacillus jordanicus strain GSFE11 had 36 contigs with a total length of 3 839 208 bp, with an N50 value of 377 721 bp, and G + C content of 39.09%. The number of coding genes was predicted to be 3961, of which 3893 genes were protein-coding and 68 genes were RNA coding. The predicted RNA coding genes consist of 59 transfer RNAs (tRNAs) and 9 rRNAs (7 5S, 1 16S, and 1 23S). The main features of the genome are presented in Table 1.
Genome features of Oceanobacillus jordanicus strain GSFE11.
The whole genome-based taxonomic analysis showed that O. jordanicus strain GSFE11 was closely related to O. picturae as shown in Figure 1a. This conclusion was also supported by the comparison of genomic signatures results (Figure 1b) placing O. jordanicus GSFE11 as a close relative to O. picturae.

(a) Whole-genome sequence-based phylogenetic tree, species cluster has the same color code, the GC% is indicated in shades of blue, genome size and protein count is presented in black and brown bars, respectively and (b) genomic signatures based on phylogenetic tree and the frequencies of octanucleotides are presented at the scale under the tree.
Oceanobacillus jordanicus GSFE11 has 62.7% digital DNA-DNA hybridization (dDDH) score, and 0.07% G + C% content difference with O. picturae DMTMMB24. This observed dDDH of 62.7% was lower than 70% which is the species boundary value for defining bacterial species as closely related species. 32 The difference in genome size and number of coding genes between O. jordanicus and O. picturae (Genome size ~3.8 bp vs ~3.6bp, number of coding genes 3961 vs 3823, respectively). In addition, the difference in the genomic signature as seen in the second phylogenetic tree in Figure 1b, also supports our proposition that the isolated bacteria O. jordanicus is a new species.
To further assess the difference between O. jordanicus GSFE11 and other closely related species identified by whole-genome alignment (represented in the phylogenetic trees of Figure 1), we ran a pangenome analysis, which represents the collection of all genes found in a collection of related organisms grouped by sequence homology. First by identifying clade of specific genes between the genomes of O. jordanicus against O. picturae and with O. iheyensis as an outer group (Figure 2a); then between the genome of O. jordanicus against all 20 closely related species identified during whole-genome alignment step (Figure 2b).

Pangenome analysis results: (a) between the genomes of O. jordanicus against O. picturae and with O. iheyensis as an outer group, (b) between the genome of O. jordanicus against all 20 closely related. Red lines represent singletons, which are genes with no sequence homology to genes in any other genomes, cyan lines represent non-core genes, purple lines represent clade-specific core orthologs present in all but the outgroup genome, dark blue lines represent core ortholog sets with at least 1 gene from the ortholog set with a gene in each of the genomes, gray lines represent pangenome non-core, which are orthologs between Core and Singleton clusters present in more than 1 genome and fewer than all.
The differences in pangenome analysis results between O. jordanicus against O. picturae, represented by the high degree of base singletons (unique sequences), further supports our classification of O. jordanicus GSFE11 as a new species.
Classification of CRISPR-Cas variants could capture the evolutionary relationships between species, and can help in comparative genomic analysis. 33 We were able to identify only one CRISPR system with 1 spacer (Table 2). While O. picturae DMTMMB24 genome has 2 CRISPR system with 2 spacers and different repeat consensus in comparison to O. jordanicus GSFE11.
Clustered Regularly interspaced Short Palindromic Repeats (CRiSPR) sequences present within O. jordanicus strain GSFE11 identified using CRiSPRCasFinder.

Functional classification of the genes in O. jordanicus GSFE11 based on RASTtk annotation implemented in PATRIC server 31 was carried out. The annotation included 1328 hypothetical proteins and 2633 proteins with functional assignments (Table 3). The proteins with functional assignments included 828 proteins with Enzyme Commission (EC) numbers, 34 701 with Gene Ontology (GO) assignments, 35 and 616 proteins that were mapped to KEGG pathways. 36 PATRIC annotation includes 2 types of protein families, 37 and this genome has 3501 proteins that belong to the genus-specific protein families (PLFams), and 3597 proteins that belong to the cross-genus protein families (PGFams).
Protein features of O. jordanicus strain GSFE11.

When compared to annotations of close species present in the phylogenetic tree (Supplementary Figure 1), O. jordanicus has 3499 genes present in 3344 homolog families, and 461 singletons (Supplemental Table 1). It also showed that O. jordanicus has 3082 shared homolog families with O. picturae, and less than 2000 homologs with other related species (Supplemental Table 2). Further supporting the proposition that O. jordanicus is a new species.
The annotation showed a large number of the genes predicted (Supplemental Figure 2) are involved in the metabolism of carbohydrates, genes for xylose, fructose, and chitin, quinone cofactors phylloquinone and menaquinone biosynthesis (they function as an electron transporter in photosynthesis as a redox molecule). Genes associated with nitrogen, sulfur, phosphate and iron acquisition were present, in addition to genes involved in the biosynthesis of plant hormone auxin, such genes might be important for symbiotic plant-bacteria interaction.
Genes possibly involved in transport, cell motility, delivery systems, secretion, membrane proteins and plant cell wall modification, siderophores, transcriptional regulators, and substrate utilization were identified. Genes involved in detoxification and stress protection against heavy metals such as cobalt, zinc, cadmium, and chromium were present (Table 4). In addition to antibiotic resistance genes against fosfomycin and fluoroquinolones. Genes involved in bile hydrolysis were identified, hinting at a possible lifestyle of O. jordanicus GSFE11 as a gut microbe. (The full list of annotated genes is available online: BioProject ID PRJNA753452).
List of key specialty genes present in Oceanobacillus jordanicus GSFE11.

O. jordanicus strain GSFE11 has been shown to have a plant growth-promoting effect. 10 For instance, it was the most effective bacterial strain with antifungal activity against a very aggressive isolate of Fusarium culmorum. In addition, O. jordanicus GSFE11 was able to produce indole acetic acid (IAA) and has a set of genes that potentially contributed to an improved tolerance against drought and salinity in durum wheat. In the light of the current draft genome, future transcriptome and molecular analysis studies will help elucidate many of the hypothetical proteins predicted here and the possible mechanism by which such endophyte enhances the growth of plants. Bacterial whole genome sequencing followed by comparative genomic analysis also proved to be a necessary and sufficient procedure to distinguish between closely related bacterial species.38-40 This approach will help improve our understanding and increase the discovery rate of new species in environmental samples.
Supplemental Material
sj-docx-1-evb-10.1177_11769343211071114 – Supplemental material for Draft Genome Sequence of Oceanobacillus jordanicus Strain GSFE11, a Halotolerant Plant Growth-Promoting Bacterial Endophyte Isolated From the Jordan Valley
Supplemental material, sj-docx-1-evb-10.1177_11769343211071114 for Draft Genome Sequence of Oceanobacillus jordanicus Strain GSFE11, a Halotolerant Plant Growth-Promoting Bacterial Endophyte Isolated From the Jordan Valley by Tareq Alhindi and Randa Albdaiwi in Evolutionary Bioinformatics
Footnotes
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The Deanship of Scientific Research – The University of Jordan (Grant Number: 2350).
Declaration of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Work was planned by RA and TA. Sample collections, identification and molecular work was done by RA. Bioinformatic analysis was done by TA. Manuscript was written by TA, and both authors read and approved the final manuscript.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.