
Abstract
Purpose
To characterise the sequential macular changes using spectral-domain optical coherence tomography (SD-OCT) in acute retinal necrosis (ARN) and compare these with the von Szily model of neuronally transmitted herpetic retinitis.
Methods
Single-patient case study with serial SD-OCT imaging and analysis of aqueous humour varicella zoster virus (VZV) levels.
Results
A 30-year-old immunocompetent woman presented with 360° confluent peripheral retinal necrosis and scattered multifocal lesions in the right eye. VZV DNA was detected in the aqueous humour by polymerase chain reaction (PCR). At baseline, multiple radial retinal folds around the fovea were visible, appearing as sharp inner retinal undulations on SD-OCT. Numerous continuous and beaded vertical hyperreflective strips (VHRS) were present within the outer nuclear layer; in the parafovea, these followed the oblique course of the Henle fibre layer. By days 4–5, neurosensory detachments developed beneath VHRS, with subfoveal choroidal thickness peaking at day 9. Despite systemic and intravitreal antivirals, the right eye deteriorated to light perception, hypotony, and phthisis.
Conclusion
Sequential macular SD-OCT in VZV-associated ARN revealed inner retinal undulations and vertical hyperreflective strips in the outer nuclear layer preceding neurosensory detachments. These observations may correspond to viral dissemination along neuronal pathways from the inner to outer retina, consistent with mechanisms described in animal models. Recognition of these imaging biomarkers could support early detection of macular involvement and improve understanding of ARN pathogenesis in humans.
Keywords
Introduction
Acute retinal necrosis (ARN) was first described by Urayama et al. in 1971 1 It is most commonly caused by varicella-zoster virus (VZV), herpes simplex virus type 1 (HSV-1), and herpes simplex virus type 2 (HSV-2). 2 These double-stranded DNA neurotropic viruses establish lifelong latency within human ganglia, and can reactivate later in life. 3 While the clinical manifestations of ARN are well characterised, and diagnostic criteria defined,4,5 the mechanisms underlying pathogenesis in humans remains poorly understood. 6
In animal models, particularly the von Szily model, 7 herpetic retinitis arises through neuronal propagation of virus from the central nervous system to the retina.8,9 Genetically tagged viral strains have confirmed direct axonal transport to the eye along ganglion cell pathways, with spread from the inner to the outer retina. 8 However, direct in-vivo evidence of such stepwise neuronal dissemination in humans has been limited.
Here, we report a case of VZV-associated ARN with detailed multimodal imaging, including sequential SD-OCT, and VZV detection from aqueous samples. We evaluate whether the observed structural changes align with neuronal viral spread described in animal models, offering new insights into human pathogenesis.
Case report
Clinical presentation and examination
A 30-year-old immunocompetent woman presented with a one-week history of right eye irritation, redness, and blurred vision. She reported no recent infection, steroid exposure, or vaccination. The patient reported severe childhood varicella but no recent history of cutaneous lesions suggestive of shingles. HIV serology was negative, and lymphocyte subsets, immunoglobulins, and HbA1c were within normal limits. Visual acuity was 6/24 in the right eye (OD) and 6/9 in the left eye (OS) with correction. Intraocular pressures were 20 mmHg (OD) and 19 mmHg (OS). The right eye exhibited 2 + anterior chamber cells, fine keratic precipitates, and grade 1 vitreous haze. There was no iris atrophy, heterochromia or posterior synechiae. The anterior chamber of the left eye was unremarkable. Ultra-widefield imaging of the right eye (Optos California, Dunfermline, UK) revealed 360° of confluent peripheral retinal necrosis with superficial haemorrhages and scattered multifocal, yellow–white lesions of varying sizes not aligned to the retinal vasculature (Figure 1). The patient was diagnosed with ARN and commenced on oral valaciclovir 2 g three times daily, together with topical dexamethasone 0.1% eye drops every two hours. Anterior chamber paracentesis confirmed high-level VZV DNA by PCR (Ct 22.3), while assays for herpes simplex viruses 1 and 2, cytomegalovirus (CMV) and Epstein-Barr virus were negative. Whole genome sequencing did not reveal any mutations associated with antiviral resistance in the DNA polymerase gene (ORF 28) or the thymidine kinase gene (ORF 36).
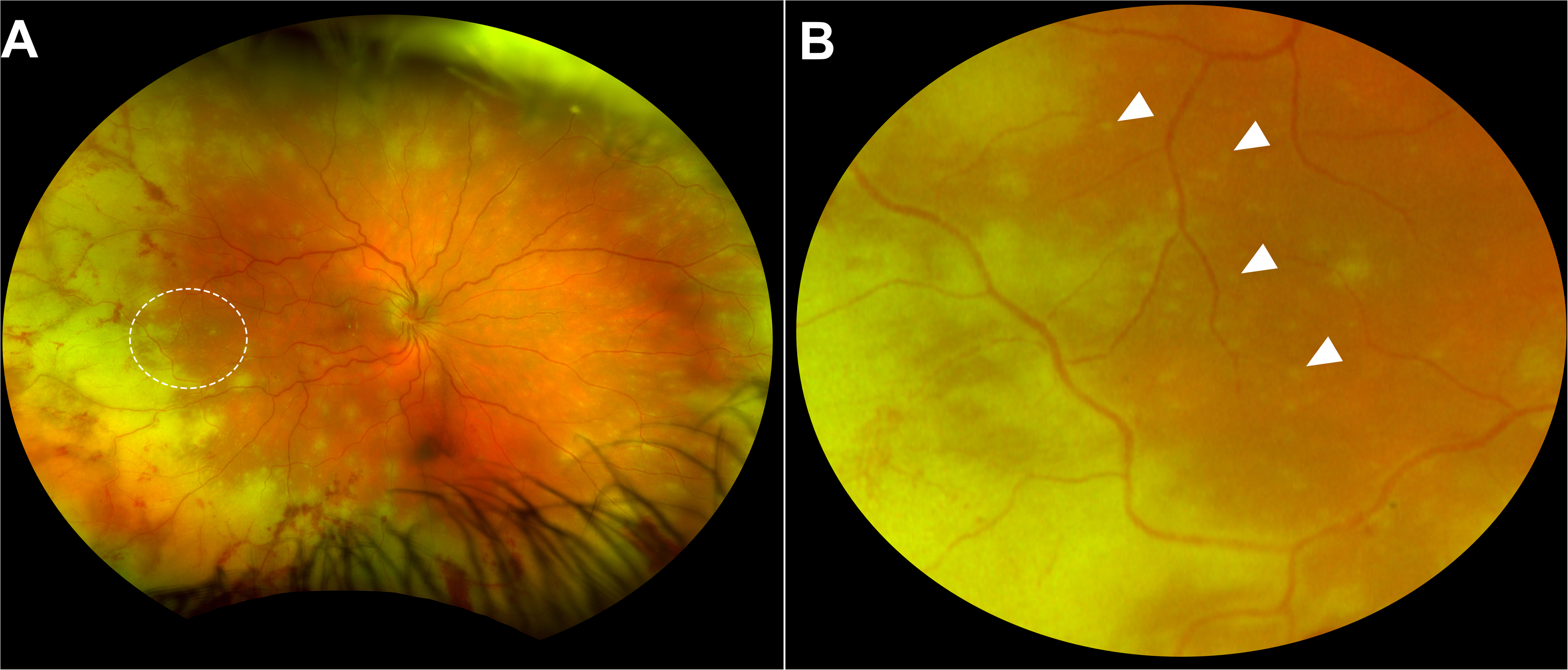
Fundus appearance of the right eye at presentation. (A) Ultra-widefield image showing 360°confluent and non-confluent areas of acute retinal necrosis in the peripheral retina, with haemorrhagic vasculitis involving arteries and veins. The white ovoid highlights the area of enlargement. (B) Magnified view of the ovoid area. White arrows indicate multiple small, yellow–white non-confluent foci of early necrosis.
Baseline macular infrared imaging of the right eye showed peripapillary wrinkles and multiple radial inner retinal folds centred on the fovea (Figure 2A) which appeared as sharp undulations of the inner retinal surface resembling a ‘saw-tooth’ on spectral-domain OCT (SD-OCT; Heidelberg Spectralis, Heidelberg Engineering, Germany), some of which exhibited hyperreflective foci at their peaks (Figure 2B and C). In some areas, hyporeflective cavities within the ganglion cell layer were visible suggestive of necrosis (Figure 2B) whereas in the temporal macular, atrophy was present in the ganglion cell layer (Figure 2D).
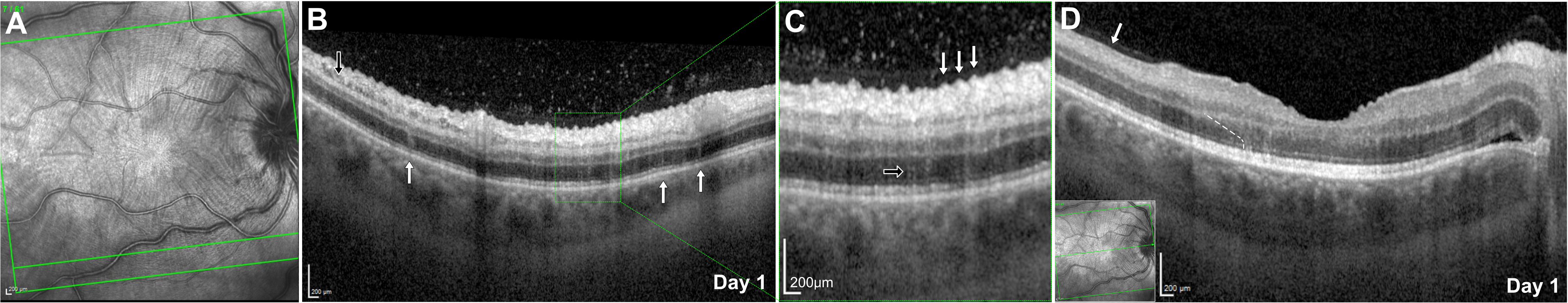
Right eye inner retinal folds and VHRS at presentation. (A) Infrared imaging demonstrating multiple radial inner retinal folds around the fovea, oriented perpendicular to the arcuate course of the retinal nerve fibre layer. The green line indicates the site of SD-OCT acquisition shown in panel B. (B) SD-OCT beneath the inferior arcade of the right eye. The black arrow marks cavitations in the ganglion cell layer. White arrows indicate multiple VHRS in the outer nuclear layer. The green box corresponds to the enlargement in panel C. (C) SD-OCT showing the serrated inner retinal border (“saw-tooth” appearance) (white arrows). Enlargement highlighting a beaded vertical hyperreflective strip (black arrow). (D) SD-OCT showing a parafoveal beaded VHRS aligned with the Henle fibre layer. White dashed line indicates the angulation. Insert box shows scan acquisition location.
Numerous scattered, outer nuclear layer vertical hyperreflective lines (termed vertical hyperreflective strips (VHRS) in prior literature10–12 were present on SD-OCT at baseline. These showed two morphologies: continuous thin lines and beaded lines resembling a ‘beads-on-a-string’ pattern (Figure 2C). Perifoveally, several beaded VHRS followed the oblique trajectory of the Henle fibre layer (Figure 2D). A small, isolated peripapillary neurosensory retinal detachment retinal was also present at baseline (Figure 2D). Sequential SD-OCT imaging at day 4 and 5 demonstrated increases in both retinal thickness and ipsilateral optic disc swelling. Subtle peripapillary inner retinal folds were observed at baseline in the asymptomatic left eye which remained unchanged throughout the presentation (Supplemental Figure 1).
By day 5, multiple, focal areas of neurosensory retinal detachments occurred in areas of VHRS (Figure 3 and Supplemental Figure 2). Underlying some areas of neurosensory retinal detachment, a separation of the retinal pigment epithelium and Bruch's membrane was visible (Figure 3). Subfoveal choroidal thickness was 325μm at baseline and peaked at 468μm by day 9. On day 15, a large intraretinal cyst could be seen at the fovea with an eruption-like appearance extending from the inner retina (Figure 3).
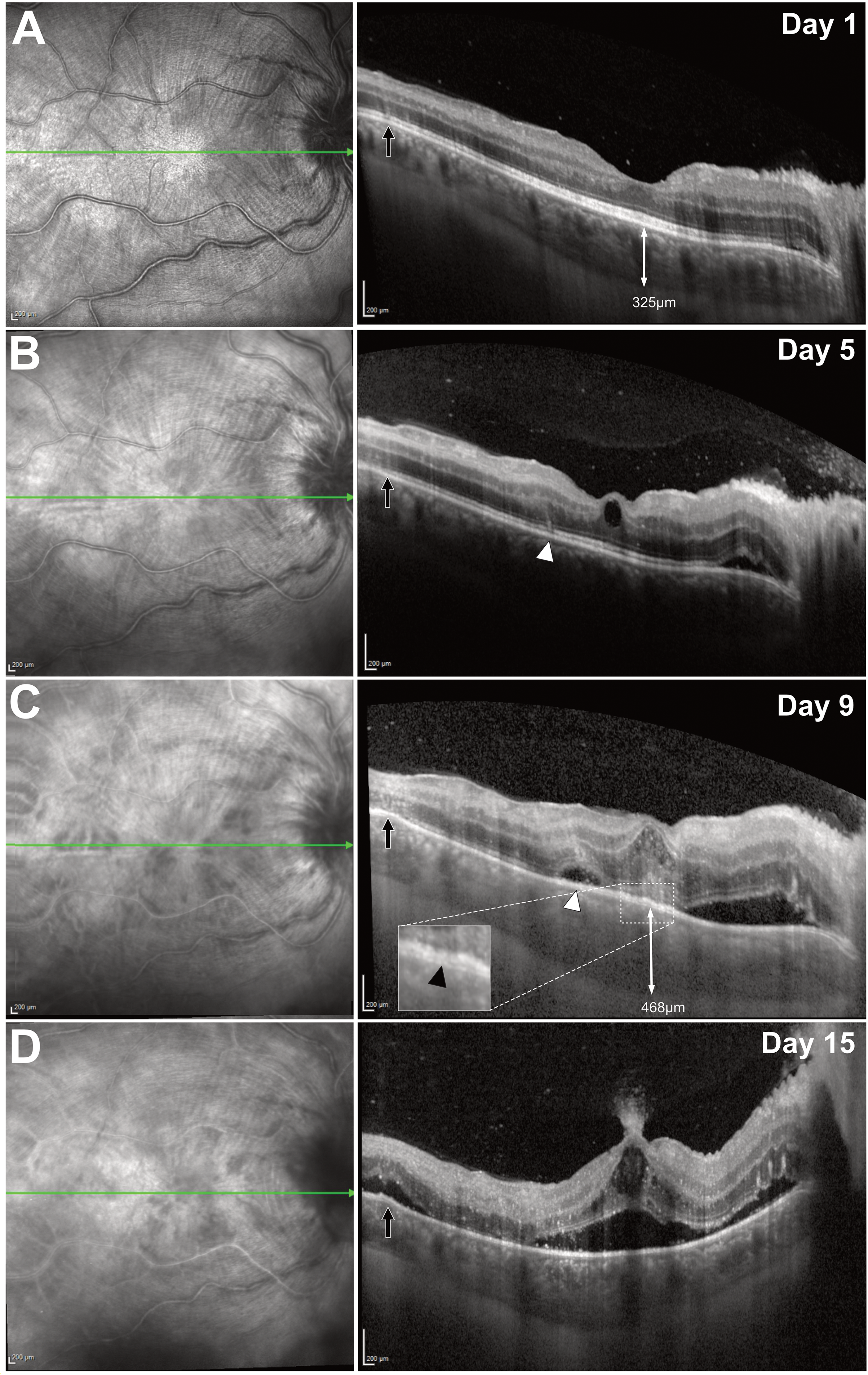
Sequential macular SD-OCT of the right eye. (A) Baseline image showing a small peripapillary neurosensory detachment with peripapillary wrinkles. Black arrow highlights several VHRS. (B) Day 5: appearance of a perifoveal VHRS following the Henle fibre layer trajectory (white arrowhead). (C) Day 9: neurosensory detachment at the site of a prior VHRS (white arrowhead). Insert highlights example of retinal pigment epithelium and Bruch's membrane separation (black arrowhead). (D) Day 15: neurosensory detachment at the site of the previous temporal VHRS (black arrow), with underlying retinal pigment epithelium –Bruch's membrane separation. Central and peripapillary detachments have coalesced, with a plume of hyperreflective material seemingly erupting from the foveal inner retina. Between days 1 and 9, subfoveal choroidal thickness increased in the affected right eye with no change in the unaffected left eye.
Despite oral valaciclovir 2 g three times daily from presentation, clinical deterioration, and a rising aqueous VZV load by Day 5 prompted an escalation of therapy to intravenous aciclovir 10 mg/kg three times daily and intravitreal foscarnet (2.4 mg/0.1 ml) which was repeated every 3rd day for 19 days. A further 5 intravitreal foscarnet injections were given with increasing intervals until day 48, totalling 12 injections. Three intravitreal injections of ganciclovir (2 mg/0.1 ml) were also administered from day 18 with a 3-day interval between injections.
Graphical representation of aqueous humor VZV log10 value, systemic and intravitreal antiviral treatment is outlined in Supplemental Figure 3. The VZV values progressively declined with a small plateau after venous access was lost on day 20 resulting in treatment transitioning to oral valaciclovir 2 g four times daily. Despite treatment, the right eye deteriorated, developing cataract and progressive hypotony, ultimately becoming phthisical, with final visual acuity reduced to light perception. At three-year follow-up, the fellow eye remains unaffected with 6/5 vision on prophylactic oral valaciclovir 1 g once daily.
Discussion
The neuronal dissemination of herpesvirus-induced retinal necrosis has been well established in animal models,6,8,13 yet the mechanisms underlying clinical disease in humans remain incompletely understood. 6 The von Szily model first demonstrated neuronal propagation when inoculation of HSV into the anterior chamber of a rabbit model resulted in contralateral retinitis. 7 Subsequent studies using electron-microscopy and genetically tagged HSV strains confirmed direct axonal transport to the retina along ganglion cell axons and descent through the retina in a multifocal pattern.8,13 Notably, Vann et al reported scattered foci throughout the retina and infected cell foci oriented vertically within the retinal plane, extending partially or completely from the ganglion cell layer to the photoreceptor layer. 8
To date, evidence in humans of neuronal dissemination of herpetic virus from the central nervous system to the eye is supported by the occurrence of bilateral ARN following intracranial injection of oncolytic HSV-1, 14 eleven reported cases of the von Szily reaction in human patients, 15 cases of ARN preceded by optic neuropathy,16,17 arcuate nerve-fibre changes extending from the disc to macula,18,19 and the observation of inner necrosis preceding outer necrosis.18,20 In the present case, sequential SD-OCT imaging provided a rare opportunity to observe early macular changes that are consistent with the neuronal dissemination sequence described in animal models.
We are unaware of prior reports of saw-tooth like undulations of the inner retina associated with ARN as seen in this case (Figure 2). These appear comparable to peripapillary wrinkles observed in several causes of optic disc swelling,21,22 but extend beyond the peripapillary region. Similar undulations which increased in number and amplitude in the days preceding areas of vertical outer nuclear layer hyperreflectivity in the sequential eye involvement of a case of relentless placoid chorioretinitis have been reported. 23 The spoke-like arrangement of the inner retinal folds around the fovea (Figure 2A) appears perpendicular to the arcuate anatomical trajectory of the ganglion cell axons that form the retinal nerve fibre layer (RNFL). We hypothesise a contractile force within this layer may account for the undulations potentially due to a disruption in axoplasmic flow. It is unclear if this occurs in ARN as a consequence of viral spread within ganglion cell axons, however similar sharp undulations of the inner retina were seen on histological specimens in the von Szily model. 13
VHRS are a repeatedly described OCT feature of ARN.10,12 While not pathognomonic, their presence may aid in differentiating herpetic infections from toxoplasmic retinochoroiditis, in which VHRS were not observed. 10 In our case, VHRS appeared narrow and often beaded (Figure 2, Supplemental Figure 2), although broader bands resembling those observed in acute posterior multifocal placoid pigment epitheliopathy have also been described in ARN.11,24 In contrast to the confluent, full-thickness hyperreflective columns observed in vitreoretinal lymphoma, VHRS are predominately limited to the outer nuclear layer. 25 Furthermore, we observed that outside the perifoveal region VHRS were vertically orientated, whereas in the perifovea they followed the oblique trajectory of the Henle fibre layer. While no histological confirmation of their aetiology currently exists, we hypothesised – based on their anatomical orientation – that both continuous and beaded VHRS, including those aligned with the Henle fibre layer, may represent structural consequences of viral propagation along photoreceptor axons and/or Müller cell processes. It has previously been hypothesised that the beaded appearance may represent the formation of axonal spheroids, 23 secondary to disrupted axoplasmic flow, 26 which can occur as a consequence of neuronal viral infection. 27
The occurrence of serous detachments in ARN has also been previously observed.11,28 However, the observation of VHRS preceding the formation of serous detachments, to the best of our knowledge, has not been previously described. If VHRS arise in ARN due to viral transmission from the inner to the outer retina – consistent with neuronal spread seen in animal models – this could account for the focal neurosensory retinal detachments. These detachments may result from an inflammatory response and/or a localised increase in choroidal permeability. In some instances, on the basis of the tenting appearance of the ellipsoid zone (Supplemental Figure 2, example B and C) a vertical intraretinal traction along the orientation of the VHRS may account for the neurosensory detachment and the retinal pigment epithelium/Bruch's membrane separation given the prior photoreceptor-retinal pigment epithelium interdigitation. A schematic of this hypothesised pathophysiology is outlined in Supplemental Figure 4.
The absence of increased choroidal thickness in viral ARN has been suggested as a useful distinguishing feature from toxoplasmic retinochoroiditis,10,29 However, significant increases in choroidal thickness were observed in our case (Figure 3) as have been reported in other cases,11,28,30 suggesting this feature may not be a reliable sign for distinguishing viral ARN from toxoplasmic retinochoroiditis. Because serous neurosensory retinal detachments often coexists with choroidal thickening in ARN, a primary choroidal inflammatory pathogenesis is commonly presumed. 11 By contrast, the preceding retinal changes in our case suggest that the choroidal thickening is a secondary, reactive phenomenon, as others have hypothesised. 31 This interpretation is supported by animal models in which choroidal vessels are dilated and contain numerous polymorphonuclear leucocytes, yet no virus is detected within the choroid. 13
Although the response to treatment was not the primary focus of this paper, it is notable that the aqueous VZV load initially increased despite oral valaciclovir (2 mg three times daily) (Supplemental Figure 3) and again following the switch from intravenous aciclovir to oral therapy. This may reflect either a lag in achieving therapeutic intraocular levels with oral valaciclovir or potentially superior efficacy of intravenous aciclovir in acute disease.
Limitations
There are several limitations to this study. First, the reliance on in-vivo imaging necessitates interpretation of secondary retinal changes as indirect evidence of viral passage, in the absence of histopathological confirmation. Second, the findings are based on a single case, and further examples are needed to provide verification. Consequently, the observations may not be considered generalisable without corroboration in larger cohorts.
Conclusion
This case provides a unique window into the acute dynamic retinal changes occurring in varicella-zoster virus–associated acute retinal necrosis, captured through sequential SD-OCT imaging. We describe a potential sequence of events beginning with inner retinal undulations, and VHRS compatible with viral transit toward the outer retina, culminating in secondary serous detachments. While this mechanism remains hypothetical, the sequence is consistent with the neuronal pathway described in animal models, offering novel insights into the pathophysiology of viral retinal infection in humans. Further cases and experimental studies will be essential to validate this model and clarify its relevance to human disease.
Supplemental Material
sj-docx-1-ejo-10.1177_11206721251388239 - Supplemental material for Sequential multimodal imaging of varicella-zoster virus-associated acute retinal necrosis: Insights supporting a neurotropic hypothesis
Supplemental material, sj-docx-1-ejo-10.1177_11206721251388239 for Sequential multimodal imaging of varicella-zoster virus-associated acute retinal necrosis: Insights supporting a neurotropic hypothesis by Paul J Steptoe, Sandeep Ramalingam and Baljean Dhillon in European Journal of Ophthalmology
Footnotes
Acknowledgements
We thank the Princess Alexandra Eye Pavilion's ophthalmic imaging team for the imaging provided.
Ethics
The requirement for consent was waived by the ethics committee due to the retrospective data collection. Written informed consent for publication of this case and associated images was obtained from the patient, in line with institutional guidelines
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.