
Abstract
Keywords
For immigrant mothers navigating the challenges of an unfamiliar sociopolitical landscape, the burden of discrimination often extends beyond their own experiences. Over time, cumulative exposure to discrimination may influence the environments in which young children grow and develop, with potential biological effects across the lifespan.(Shonkoff et al., 2021). Chronic conditions such as depression and cardiovascular disease, along with their underlying stress biomarkers, often worsen in second-generation Latino immigrants compared to their foreign-born parents (Fox et al., 2015; Scholaske et al., 2021). Although the biological foundations of these conditions are laid early in life (Miller et al., 2011), the role of postmigration adversity during early childhood has been largely overlooked. Psychobiological research with immigrant populations (Anisman et al., 2024) and studies of discrimination (Curenton et al., 2024) have both focused primarily on school-aged children and adolescents, limiting opportunities to intervene during sensitive early developmental periods.
Guided by two developmental theories, the Linked Lives principle and the Biological Embedding of Childhood Adversity Model, we examine the extent to which mothers’ experiences of discrimination in the past year may be related to transcriptional stress biomarkers in their preschool-aged children (3–5 years old
Linked Lives
Linked Lives, a cornerstone of life course theory, proposes that family members’ lives are interdependently lived within the socio-historical and -political context, an interdependence that may connect families across generations (Elder, 1998). These interconnections are more likely to shape life trajectories during sensitive developmental periods and consequential life transitions, such as immigration (Elder, 1998). In this way, immigration-related stress, particularly exposure to post-migration discrimination, may mutually influence maternal and child well-being through shared environmental influences. Although some immigration-related stressors may diminish over time, discrimination may be recognized and felt most profoundly for immigrant families who have most acculturated to the destination country (Flippen & Parrado, 2015; Velásquez et al., 2024).
Interpersonal discrimination refers to unfair treatment that restricts access to resources and opportunities based on perceived social group membership, including race and immigration status; it may be experienced directly or indirectly, such as through caregiver stress (Shonkoff et al., 2021). For immigrants who have been racialized (i.e., assigned to a racial or ethnic category) in the destination country, discrimination is a critical facet of acculturative stress—the challenges immigrants face post-migration as they adapt to the host country.
Discrimination is considered one of the most predominant acculturative stressors and is consistently associated with high levels of psychological distress (Cervantes et al., 2016). As a pervasive and uncontrollable emotional threat, these experiences increase both parenting stress (Condon et al., 2022) and economic strain, particularly among immigrant mothers of young children (Ettinger de Cuba et al., 2025). Discrimination may also exacerbate the social isolation mothers may experience when raising a child far away from familial support and cultural traditions (Lim et al., 2022). In turn, heightened maternal stress and psychological distress can disrupt the quality of the mother-child relationship, including attachment, which scaffolds a young child’s emerging self-concept and social and emotional skills (Bowlby, 1980). Taken together, these interrelated experiences reflect the intergenerational transmission of discrimination-related stress embedded within broader social environments.
By preschool age, Latino children may also witness discrimination against caregivers or experience it themselves (Barajas-Gonzalez et al., 2022; Cervantes et al., 2018). These experiences may continue to shape a child’s sense of self and the social safety of the world (Cervantes et al., 2018; Heard-Garris et al., 2018). In these contexts, children may also develop negative schemas about the social safety of their surroundings and their self-worth — processes that may be bi-directionally linked to immune activation (Slavich, 2020).
Biological Embedding of Childhood Adversity
Toxic stress occurs when a child is chronically exposed to intense stress without sufficient support, as seen in the context of interpersonal and structural discrimination (Cervantes et al., 2018; Shonkoff et al., 2021). According to the Biological Embedding of Childhood Adversity Model, toxic stress can become embedded in the developing hypothalamic-pituitary-adrenal (HPA) and immune axes during early childhood, with implications for lifelong well-being (Miller et al., 2011). The HPA axis, responsible for coordinating the stress response, releases cortisol during threats, which typically suppresses excessive immune activity and restores balance. However, adverse environments can impair this recovery by reducing glucocorticoid receptor activity, prolonging stress responses, and increasing the production of proinflammatory proteins (e.g., IL-8, IL-1β, TNF-α) from the immune system (Nusslock & Miller, 2016).
The relationship between toxic stress and immune dysregulation (i.e., inflammation) has been detected in early childhood (Measelle & Ablow, 2018; Nelson et al., 2020; Tyrka et al., 2015), a period of heightened neuro-immune plasticity (Danese & J Lewis, 2017). Furthermore, in two studies thus far, maternal experiences of ethnic and racial discrimination have been linked to elevated levels of inflammatory proteins in school-aged children (Condon et al., 2019; Slopen et al., 2019). These associations have persisted even after adjusting for maternal depressive symptoms and child body mass index (BMI), a correlate of low-grade inflammation (Slopen et al., 2019). At a transcriptional level, pro-inflammatory mRNA transcripts may be upregulated in response to children’s exposure to and risk for toxic stress (Cole, 2019). Animal models suggest that toxic stress may become embedded in the transcriptome of immune cells as early as the postnatal period (Cole et al., 2012). By adulthood, a pro-inflammatory phenotype underlies most chronic health conditions, contributing to the leading causes of morbidity and mortality worldwide (Furman et al., 2019).
Guided by the Linked Lives principle and the Biological Embedding of Childhood Adversity model, we pursued two a priori aims. First, we tested whether maternal post-migration discrimination was associated with preschoolers’ pro-inflammatory mRNA expression of IL-8 and IL-1β. Second, we evaluated the robustness of these associations by examining whether they attenuated after accounting for maternal depressive symptoms and child BMI.
Methods
Study Design
We conducted a cross-sectional observational study to examine the associations between Latina mothers’ experiences of discrimination and their preschoolers’ pro-inflammatory gene expression. Data were collected May 2022–August 2023 via community-based recruitment. The study was approved by the University of North Carolina Chapel Hill (UNC-CH) Institutional Review Board (IRB).
Sample and Setting
Mother-Child Characteristics
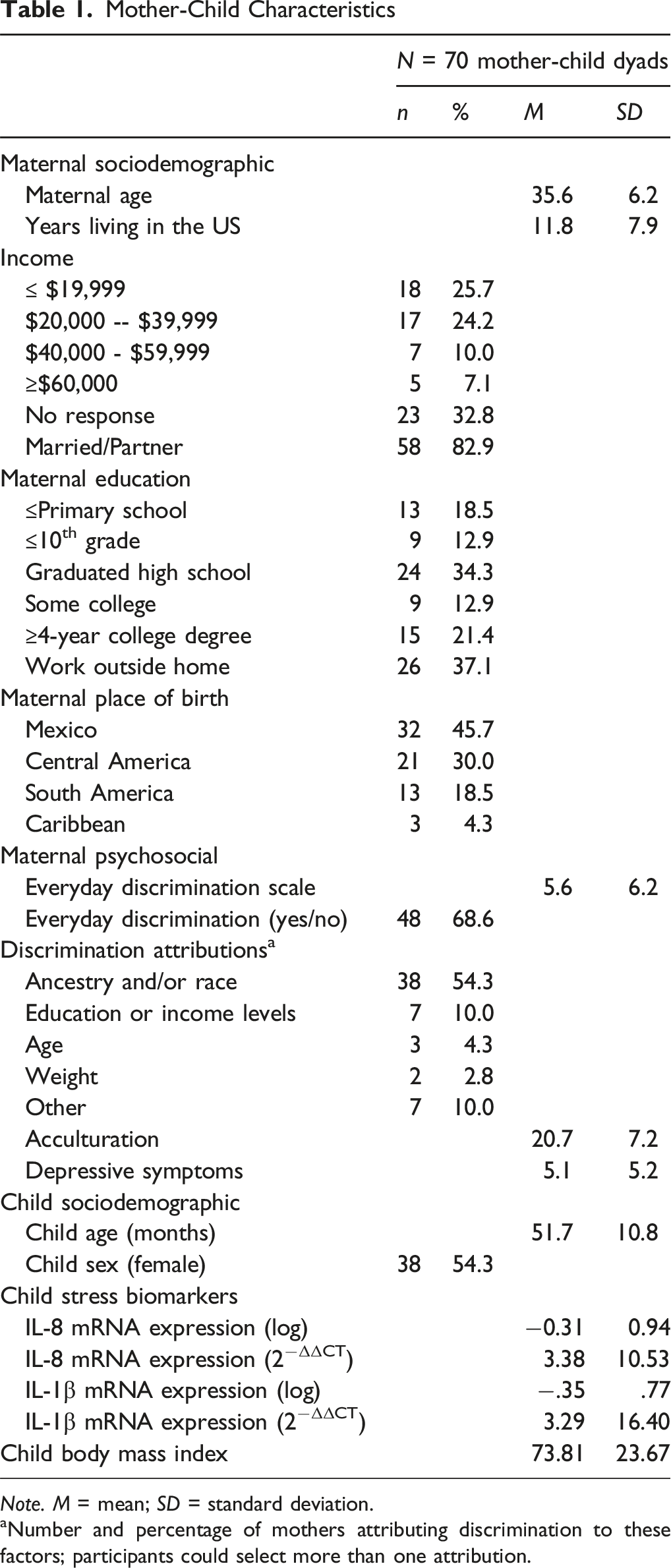
Note. M = mean; SD = standard deviation.
Number and percentage of mothers attributing discrimination to these factors; participants could select more than one attribution.
Measures
Maternal Measures
Maternal Everyday Discrimination
The 9-item Everyday Discrimination Scale (EDS) (Williams et al., 1997) was used to measure the frequency of day-to-day perceived discrimination within the past year. It correlates with measures of institutional racial discrimination and interpersonal prejudice but does not prime the subjects to think about race or ethnicity (Krieger et al., 2005). Participants are asked to rate each item on a 5-point scale (1 = never to 5 = almost every day). Higher scores indicate higher frequency of perceived discrimination. The EDS is validated in Spanish (Park et al., 2018). For this sample, Cronbach’s α = .87.
Child Measures
Child Immune mRNA Expression (IL-8 and IL-1β)
Summary of Raw Ct Values for RT-qPCR Assays
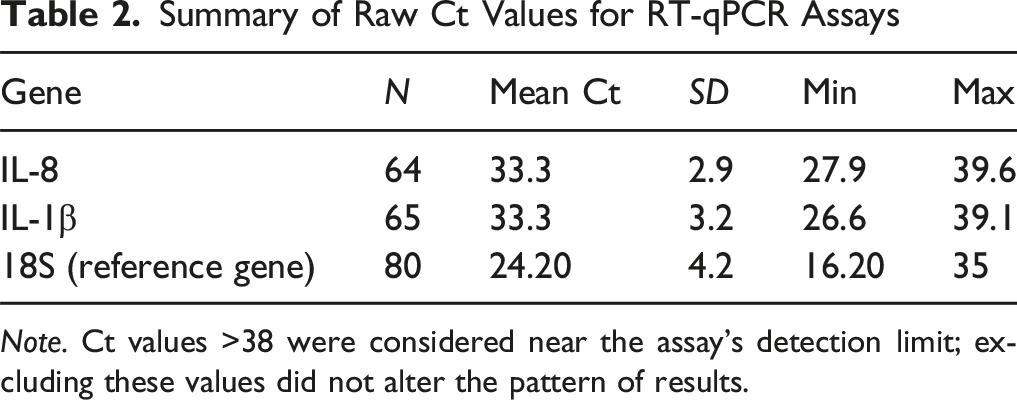
Note. Ct values >38 were considered near the assay’s detection limit; excluding these values did not alter the pattern of results.
Maternal and Child Covariates
Maternal Covariates
Maternal covariates included acculturation, education, and depressive symptoms. Maternal acculturation was measured using the 12-item Short Acculturation Scale for Hispanics (SASH) (Marin et al., 1987). SASH assesses participants’ language use and preference on a 5-point Likert scale (1 = Only Spanish to 5 = Only English). Higher scores indicate greater acculturation (i.e., higher engagement with the English language). For this sample, Cronbach’s α = .89. Maternal education was coded from (1 = Primary school to 6 = Post-graduate degree). Maternal depressive symptoms were measured using the 9-item Patient Health Questionnaire (PHQ-9) (Kroenke et al., 2001), which has been validated in Spanish (Martinez et al., 2023). Participants rate how often they have experienced each item in the preceding two weeks on a 4-point Likert scale (0 = not at all to 3 = nearly every day). The cut-off for clinical depression is ≥10. For this sample, Cronbach’s α = .88.
Child Covariates
We adjusted for child age (months) and sex (female = 1; male = 0). Mothers were asked whether their child had any health conditions; given the low rates of reported diagnoses (asthma, n = 2; autism, n = 3), we included child BMI as the primary child health covariate. Child BMI was calculated from measured height and weight and converted to age- and sex-standardized percentiles using CDC growth charts, with higher percentiles reflecting greater adiposity relative to same-age peers.
Data Collection
Data were collected in person, primarily in participants’ homes (93.5%), by the same bilingual and bicultural researcher to ensure consistency. Self- or researcher-administered questionnaires were completed in English or Spanish based on participant language preference and literacy. All procedures were approved by the UNC-CH IRB, and all mothers provided written informed consent. Given children’s young age (3–5 years old), child assent was not required.
Buccal swab samples for mother and child were collected in tubes containing DNA/RNA shield (Zymo Research, R1106-E). Samples were stored frozen at −80°C at UNC-CH until data collection was completed. Thereafter, samples were transported on dry ice to University of Miami for RNA extraction and biological analyses.
Analytical plan
Power Analyses
Sample size was determined a priori based on hypotheses, feasibility, and assay costs in a hard-to-reach, underrepresented community sample, aiming for 70–96 participants. For a single-outcome multiple regression (α = .05, two-sided), N = 70 affords detection of partial correlations of r = .28 (80% power) and r = .33 (90%), representing moderate-sized effects; smaller effects were underpowered. Joint modeling of IL-8 and IL-1β mRNA levels with correlated residuals did not reduce detectable effects relative to a single-outcome model.
Data Preparation and Assumption Testing
Prior to analysis, biomarker distributions and model residuals were examined for normality and heteroscedasticity. IL-8 and IL-1β mRNA were log-transformed to stabilize variance and approximate normality; Q–Q plots and Shapiro-Wilk tests supported this (IL-8 mRNA: W = 0.98, p = .45; IL-1β mRNA: W = 0.98, p = .22).
Multivariate Models Between Maternal Discrimination and Child Pro-Inflammatory Gene Expression
We fit joint two-outcome (multivariate multiple) regressions for IL-8 and IL-1β mRNA (Gaussian, identity), allowing correlated residuals. We modeled outcomes jointly using full-information maximum likelihood (FIML) under a Missing at Random (MAR) assumption, so that all available cases contributed (analytic N = 70; IL-8 available for 64 children, IL-1β for 65) (Enders & Bandalos, 2001). No predictors or covariates had missing data. All models used robust (sandwich) SEs and included a random intercept for qPCR plate, estimated with Gauss–Hermite quadrature (12 points) to ensure stable convergence. To obtain an overall (omnibus) test of the discrimination effect across IL-8 and IL-1β, we ran a likelihood-ratio (LR) test comparing the joint model with versus without the discrimination paths (df = 2). LR tests were computed under the ML variance estimator (observed information matrix); coefficient CIs use robust (sandwich) SEs. We also report the residual correlation between biomarkers. Multicollinearity was assessed using variance inflation factors (VIFs) from parallel ordinary least squares (OLS) models that included the same covariates.
Model 1 adjusted for random plate effects, child age and sex, and maternal acculturation and education (Cabrera & Hennigar, 2019; Viruell-Fuentes et al., 2012). Model 2 additionally adjusted for depressive symptoms as a sensitivity analysis due to its potential position downstream of discrimination (Berry et al., 2021). Model 3 included child BMI as an additional sensitivity analysis given the relationship between adiposity and proinflammatory gene expression (Ullah et al., 2024). Finally, we conducted exploratory Spearman correlations between plate-adjusted maternal and child mRNA expression values. These analyses were limited to the subset of dyads with both maternal and child biomarkers, (n = 43; IL-1β mRNA) and (n = 40; IL-8 mRNA). All analyses were conducted in Stata (Version 19.5), using an alpha level of .05.
Results
Sample Characteristics
In the analytic sample (
Maternal Discrimination and Child Inflammatory Gene Expression
Multivariate Models for Child Pro-inflammatory Gene Expression (IL-8 and IL-1β) (N = 70)
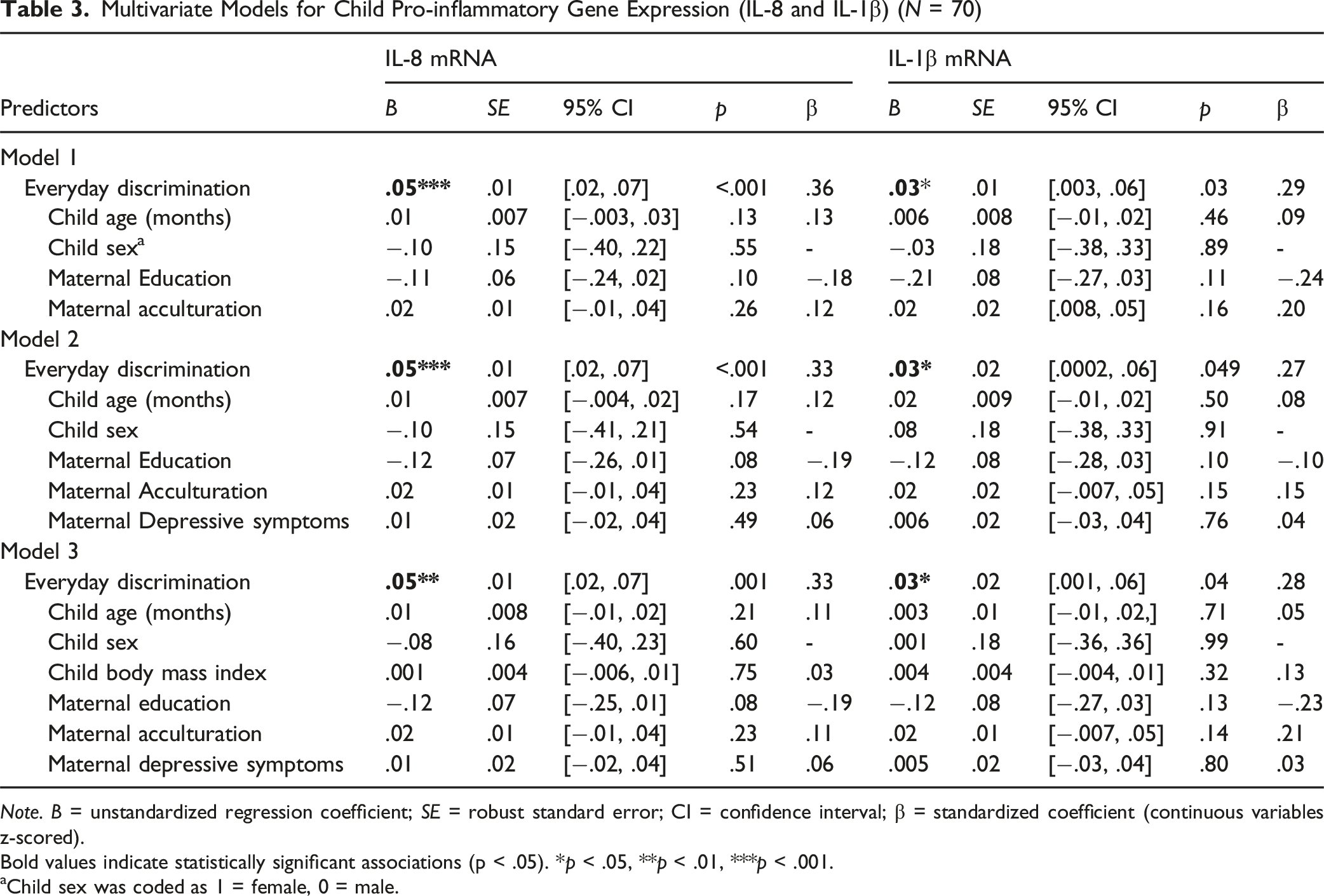
Note. B = unstandardized regression coefficient; SE = robust standard error; CI = confidence interval; β = standardized coefficient (continuous variables z-scored).
Bold values indicate statistically significant associations (p < .05). *p < .05, **p < .01, ***p < .001.
Child sex was coded as 1 = female, 0 = male.
Model 1
In Model 1, higher levels of maternal discrimination were associated with higher child IL-8 (B = 0.05, SE = 0.01, 95% CI [.02, .07], p < .001) and IL-1β mRNA expression (B = 0.03, SE = 0.01, 95% CI [0.003, 0.06], p = .03). Including the discrimination paths improved model fit across biomarkers (LR χ2(2) = 16.90, p < .001), indicating that discrimination was jointly associated with IL-8 and IL-1β expression.
Model 2
The relationship remained significant in Model 2 for child IL-8 (B = 0.05, SE = .01, 95% CI [0.02, 0.07], p < .001) and IL-1β (B = .03, SE = .02, 95% CI [0.0002, 0.06], p = .049) mRNA expression. The omnibus comparison remained significant (LR χ2(2) = 13.38, p = 0.001).
Model 3
Likewise, the relationship was significant in Model 3 for child IL-8 (B = 0.05, SE = .01, 95% CI [0.02, 0.07], p < .001) and IL-1β (B = .03, SE = .02, 95% CI [0.001, 0.06], p = .042) mRNA expression. The omnibus comparison also remained significant (LR χ2(2) = 13.19, p = 0.001).
Exploratory Analyses
Correlations between maternal and child inflammatory gene expression were small and nonsignificant (IL-8: ρ = .14, p = .39; IL-1β: ρ = .15, p = .33).
Discussion
The present study examined relationships between maternal experiences of discrimination in the past year and children’s molecular biomarkers of toxic stress during early childhood—a sensitive developmental period. Children of mothers who reported more frequent discrimination exhibited higher IL-8 and IL-1β mRNA expression, even after adjusting for maternal sociodemographic factors, depressive symptoms, and child BMI. Taken together, these findings suggest that maternal experiences of discrimination during a child’s preschool years may uniquely contribute to children’s developmental trajectories.
Our findings, while not directly predictive of future health outcomes, are best interpreted through a life course lens, consistent with the Linked Lives principle (Elder, 1998) and the Biological Embedding of Childhood Adversity model (Miller et al., 2011). These theories contextualize how early-life exposures to post-migration discrimination may shape long-term health. When mothers face repeated hardship or threat without adequate support, the shared emotional environment may calibrate their young children’s immune systems (National Scientific Council on the Developing Child, 2020). One plausible biological pathway is through the transcriptome of immune cells (Etzel et al., 2024; Marie-Mitchell & Cole, 2022). Over time, early immune programming may increase risk for inflammatory conditions in adulthood, including cardiovascular disease, asthma, and depression (Nusslock & Miller, 2016).
Unpredictable and socially threatening environments that frequently activate the stress response system may exert the strongest direct influence on the developing immune system (Kuhlman et al., 2017; Slavich, 2020). Indeed, qualitative maternal accounts from this sample revealed that experiences of discrimination often (re)produced shared environments of fear and instability (Alvarado-Harris et al., 2025). Discrimination, embedded within interconnected systems and person-place interactions, propagates and perpetuates contextual stress across multiple life domains, such as social relationships, employment, and housing (Harrell, 2000). These exposures may exert a broader influence on child outcomes than maternal depression or economic hardship alone (Harris et al., 2022). Consistent with this, cumulative social adversity was strongly associated with salivary IL-8 and IL-1β proteins in a large sample of children (N = 537, ages 0 to 17 years) (Pantell et al., 2024). Persistently high inflammation in childhood, in turn, has been linked to increased risk of mental and cardiometabolic disorders by early adulthood (Palmer et al., 2024).
Finally, our findings are also situated within broader epidemiological patterns among Latino populations, in which children of immigrants often enter adulthood with worse health than their parents (Fox et al., 2015). Consistent with this pattern, Latino individuals who grew up in the United States tend to show higher inflammatory levels by adulthood than those who migrated later in life, including during pregnancy (Alvarado-Harris et al., 2025; Scholaske et al., 2021). In turn, chronic inflammation underlies numerous early life health conditions that increase across successive post-migration generations, including low birth weight, infant mortality (Montoya-Williams et al., 2022), childhood asthma (Perreira & Allen, 2021) and early differences in brain development (Spann et al., 2024). Indeed, immigrant groups who experience the most discrimination may be the most likely to lose their initial advantage in maternal and child health (Andrasfay & Goldman, 2020; Bakhtiari, 2018; Dondero & Altman, 2024).
Limitations and Implications for Future Research
First, the cross-sectional design limits causal inference. Additionally, the modest sample size reduced power to detect small effects. There is also a risk of underreporting depressive symptoms due to social desirability bias, particularly given the stigma surrounding mental health in Latino communities (Misra et al., 2021). Although maternal gene-expression data were available only for a subset of dyads, exploratory correlations suggested that shared maternal–child inflammatory activity was unlikely to account for the observed associations
A major strength of this study is its focus on discrimination experienced by immigrant mothers while raising young children. This sociocultural and developmental context has received little attention in psychobiological research (Anisman et al., 2024; Haft et al., 2021). Most existing work linking discrimination to inflammation has focused on adults, with far less on earlier, more malleable periods for immune development (Cardenas et al., 2025). Furthermore, early proinflammatory gene expression, often preceding protein-level differences, offers a sensitive window into children’s inflammatory biology. For instance, placental IL-8 mRNA expression has been associated with child behavioral outcomes years later (Zhou et al., 2022).
At the same time, longitudinal studies are needed to clarify developmental timing and potential causal pathways. During pregnancy, maternal exposure to discrimination may confer additional or distinct immune profiles in children through maternal biological stress pathways. (Vaknine Treidel et al., 2025; Veru et al., 2014). To this end, future longitudinal studies could integrate maternal biology and use larger samples to test moderation or mediation pathways linking maternal discrimination to child physiology. Finally, chronic health conditions in children could also be examined as downstream consequences of early stress biology.
Practice and Policy Implications
Home visiting programs for families with young children (0-5 years old), such as Nurse-Family Partnership, are evidence-based two-generation models that have improved maternal and child health in randomized controlled trials (Early Childhood Home Visiting Models: Reviewing Evidence of Effectiveness 2024). Some evidence suggests they may also improve child biomarkers sensitive to early psychosocial stress, including inflammation (Condon et al., 2021). These programs may be especially beneficial for immigrant families with young children, as they can help connect families to community support and access trauma-informed care (Katsiaficas, 2020). Importantly, relationship building inherent to home visiting programs may also allow women to feel heard and valued (Lewy, 2021). However, children in immigrant families are less likely to be enrolled in home visiting programs than children with US-born parents (Katsiaficas, 2020). To increase equity, the Migration Policy Institute recommends explicitly including immigrant and refugee families in needs assessments, partnering with trusted community organizations, and tailoring service models for cultural relevance (Katsiaficas, 2020).
On a broader socio-ecological level, militarized immigration enforcement and public rhetoric scapegoating immigrants can generate and exacerbate intergenerational trauma while increasing societal costs (Cerdeña et al., 2021; Dondero & Altman, 2024). Ultimately, cultivating welcoming environments for immigrants and their children not only mitigates risk but harnesses, rather than squanders, child potential that contributes to social capital and societal well-being.
Conclusion
Early life experiences that devalue and racialize immigrants and their children may be critical – yet understudied predictors – of health disparities that develop after migration. However, the vulnerable period following migration also represents a critical window of opportunity to positively support the long-term intergenerational trajectories of immigrant families. Future studies using stress biomarkers in young children of immigrants must value and center their unique experiences. In so doing, we can identify the most salient risk, protective, and promotive factors.
Footnotes
Acknowledgements
Thank you to all the remarkable mothers and their beautiful children who participated in this study. Thank you to Nicholle Johnson for conducting all the biological analyses.
Author Contributions
Alvarado-Harris, R contributed to conception and design contributed to acquisition, analysis, and interpretation drafted manuscript critically revised manuscript gave final approval agrees to be accountable for all aspects of work ensuring integrity and accuracy Perriera, K contributed to interpretation critically revised manuscript gave final approval agrees to be accountable for all aspects of work ensuring integrity and accuracy Santos, H contributed to conception contributed to acquisition critically revised manuscript gave final approval agrees to be accountable for all aspects of work ensuring integrity and accuracy.
Funding
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Rebeca Alvarado-Harris was funded by NIH NINR under 1F31NR020843, NICHD/NIH NRSA T32 Postdoctoral Traineeship in the Biosocial Training Program 5T32HD091058-09, Caroline H. and Thomas S. Royster Fellowship, Barbara W. Madden Graduate Research Award, Alpha Chapter of Sigma Theta Tau International Research Grant, Linda Waring Matthews Research Award. Hudson P Santos Jr. was funded by Professorship start-up funds.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.